胚胎发育期经历的寄主刺激物对棉铃虫初孵幼虫嗅觉偏好性的影响
2018-09-28吴广庆李为争杨淑芳
吴广庆,李为争,杨淑芳*
(1.聊城大学 东昌学院,山东 聊城 252000; 2.河南农业大学 植物保护学院,河南 郑州 450002)
经历(Experience)是指生物与环境信息相互作用并将该过程嵌入神经系统印记的历史,也可描述为新生命诞生后沿着时间轴能够改变后天行为的系列前期事件[1]。目前已经发现昆虫世代发育链的许多环节存在明显的经历效应。例如,幼虫取食经历影响大龄幼虫的取食行为[2-3]、羽化后成虫的产卵选择[2,4]、成虫配偶选择[5-6],成虫的蜜源植物搜索经历也影响产卵寄主的搜索[7]。卵在昆虫的世代发育链上有着承上启下的独特地位。尽管不摄入任何固态食物,但卵发育期间与外界存在复杂的气体交换,这种经历可能对孵化后幼虫的行为有重要的影响[8]。
蛾类卵期经历的效应有2个互斥的假设。一方面,蛾类初孵幼虫体型小,活动能力差,自然选择会促使雌成虫将卵产在后代容易获得食物的位置。前期经历影响后期行为(如取食行为)的案例中,往往涉及外周感觉系统的逐渐“调谐”作用[9],但是在卵中胚胎的外周神经系统却是逐渐发育完善的[8]。另一方面,尽管寄主植物识别和评测主要是雌成虫的任务,但在某些情况下也要求幼虫自身具备识别陌生寄主植物的能力,例如食物被消耗完毕或幼虫从初生寄主植物上掉落的情况下[10-12]。因此,卵发育期间经历的记忆可以为这类个体重新定殖到寄主植物上提供有用的信息[13]。
棉铃虫[Helicoverpaarmigera(Hübner)]是一种食性非常广泛的农作物害虫[14]。随着转基因棉花的推广,目前棉铃虫在棉花上的危害逐渐得到了控制[15]。然而,获得批准生产的转基因农作物种类是有限的。作为一种多食性害虫,棉铃虫对其他农作物的危害也很大,尤其是2017年在新疆油葵上的危害非常严重,仍然需要大量喷施化学杀虫剂。研究卵期发育经历对初孵幼虫寄主选择行为的影响,可以为棉铃虫生态调控提供依据。例如,初孵幼虫期是对生存环境中的生物因子和非生物因子最敏感的虫态,干扰该阶段的寄主搜索或延长其寄主搜索时间,有望降低1龄幼虫基数,从而降低大龄幼虫阶段的防治成本。为此,通过使棉铃虫雌蛾在不同寄主植物粗提物处理过的纱布上产卵,收集初孵幼虫并测试其寄主选择行为,研究棉铃虫卵期发育经历对初孵幼虫寄主选择行为的影响,为棉铃虫的生态调控提供依据。
1 材料和方法
1.1 供试昆虫
2011年8月,在河南农业大学科教园区烟田的烟草花器上采集3~4龄的棉铃虫幼虫。带回室内的幼虫置于隔离的指形管(10 cm×2 cm)中,饲喂烟草嫩叶,每天更换新鲜的叶片,直到羽化。饲养幼虫用的人工气候箱条件如下:光照期的温度为(30±20)℃,暗期的温度为(28±2)℃,相对湿度为60%~70%,每天的光照期和暗期时间比为15 h∶ 9 h(气候箱的光照于每天8:00开启,23:00关闭)。 羽化之后,用10%的蔗糖溶液饲养成虫。从下一世代开始,幼虫用麦胚基人工饲料饲养。人工饲料的制备方法如下:将3 g山梨酸加入600 mL沸腾的蒸馏水中搅拌,直到其完全溶解,然后加入150 g小麦胚芽粉、80 g黄豆粉和25 g烘焙酵母;将20 g琼脂、20 g蔗糖和40 g干酪素加入到另外一份 800 mL的沸腾蒸馏水中搅拌均匀;2份材料合并之后充分搅拌,温度降低至大约40 ℃之后,加入3 g 维生素C和8 g维生素复合粉。
1.2 供试植物
烟草(Nicotianatabacum) (品种:NC89)、辣椒(Capsicumannum) (品种:中蔬6号)、番茄 (Solanumlycopersicum) (品种:金棚8号)和棉花(Gossypiumarboreum) (品种:鲁28)在河南农业大学科教园区种植。生物测定时,这些植物的株高为30~50 cm。
1.3 胚胎发育期间的经历诱导
为了消除不同寄主植物种类叶片物理性质的差异,用寄主植物乙醇粗提物处理过的纱布而不是自然的叶片作为经历诱导时雌蛾的产卵基质。从大田植株上采集棉花、烟草、辣椒和番茄的幼嫩叶片,带回室内冲洗、晾干并称质量。叶片粉碎之后,加入等质量的无水乙醇(Kermel Co., Ltd.),室温下提取48 h后过滤。这些滤液用于涂布产卵基质纱布或者用于涂布初孵幼虫测试时的滤纸碟。将棉纱布剪裁成30 cm×30 cm的方块,分别用上述4类提取物饱和处理,在通风橱中放置20 min使溶剂挥发。对照(无经历)的产卵基质用溶剂无水乙醇做同样的处理。每天黄昏时,将4对羽化3 d的棉铃虫成虫释放到圆台形的塑料笼(上口直径20 cm,下口直径15 cm,高度25 cm)中,塑料笼底部中央放置浸渍蔗糖溶液的脱脂棉球,然后将上述处理过的棉纱布罩上。不同处理的笼子转移到不同的人工气候箱中防止气味交换。每天早上检查新产的卵,一旦发现有孵化的情况及时将纱布取下,更换新的、经过相应处理的纱布,取孵化的个体进行嗅觉选择测试。
1.4 嗅觉选择测试步骤
为了确保经历诱导和随后测试使用的化学刺激物的一致性,在嗅觉偏好性测试中,将与上述相同的提取物分别涂布在直径1.5 cm的滤纸碟上作为气味源,每个滤纸碟涂布40 μL。将涂布不同粗提物的4类滤纸碟等距离交叉放置在直径14 cm培养皿的底部。溶剂挥发后,将来自特定处理组的1头初孵幼虫放入培养皿底部中央,在均匀的光照下观察幼虫对这些滤纸碟的选择反应。5 min没有反应的被试昆虫在统计分析中剔除,并测试下一头。相邻2头幼虫的测试中,不同滤纸碟的位置通过Excel随机数表进行随机化处理。每个处理组测试100头幼虫。
1.5 统计分析
初孵幼虫对不同气味源选择频次的差异用卡方测验分析。计算:(1)5个处理、4个气味源选项生物测定数据的整体差异显著性;(2)同一个处理组对不同气味源选项的差异显著性(即组内差异);(3)不同处理组对特定气味源选择频次的差异(组间差异);(4)不同处理组分别和对照组进行寄主选择模式的比较。此外,为了从整体检验结果中检出特定的处理×气味源选项组合,进一步将测试数据分为6个亚组,每个亚组抽取2个寄主气味源选项及其相应的处理组,以及对照组对这2种气味源的选择频次数据,进一步采用系列详细的卡方测验分析。自由度为1时,采用Yates修正的卡方测验方法进行分析。
2 结果与分析
2.1 卵期经历不同发育环境的棉铃虫初孵幼虫选择频次的整体卡方测验结果
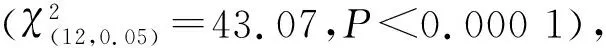
表1 卵期经历不同发育环境的棉铃虫初孵幼虫选择频次的整体卡方测验结果
注:ns、*、**分别代表无显著差异(P>0.05)、显著差异(P<0.05)、极显著差异(P<0.01)。
2.2 抽取寄主对的比较
从图1可以看出,抽取棉花-烟草寄主对进行比较时,烟草经历组对烟草的嗅觉偏好性显著强于棉花经历组(χ2= 4.03,P=0.044 7)和对照组(χ2=6.67,P=0.009 8),尽管每个组都不存在显著的组内差异。抽取棉花-辣椒寄主对进行比较时,辣椒经历组的个体对辣椒气味源的选择频次比对照组显著增多(χ2=4.87,P=0.027 3)。抽取棉花-番茄寄主对进行比较时,对照组对棉花气味源的选择频次极显著多于棉花经历组(χ2=17.92,P<0.000 1)和番茄经历组(χ2=12.79,P=0.000 3)。抽取烟草-辣椒寄主对进行比较时,烟草经历组对烟草和辣椒气味源的选择频次表现出显著的组内差异(χ2=5.49,P=0.019 1),具体表现为选择烟草的频次更多,但没有检出任何显著组间差异。抽取烟草-番茄寄主对进行比较时,既没有显著的组内差异,也没有显著的组间差异,但仍然可以看出烟草经历组对烟草的选择频次较番茄经历组更多。抽取辣椒-番茄寄主对进行比较时,对照组对辣椒气味源的选择频次极显著多于选择番茄气味源的频次(χ2=8.76,P=0.003 1),番茄经历组则显著弱化了这种对辣椒的本能性趋向偏好(χ2=6.92,P=0.008 5)。 有趣的是,卵发育期间经历辣椒粗提物的个体,对辣椒气味源的嗅觉偏好性却没有进一步的提升。
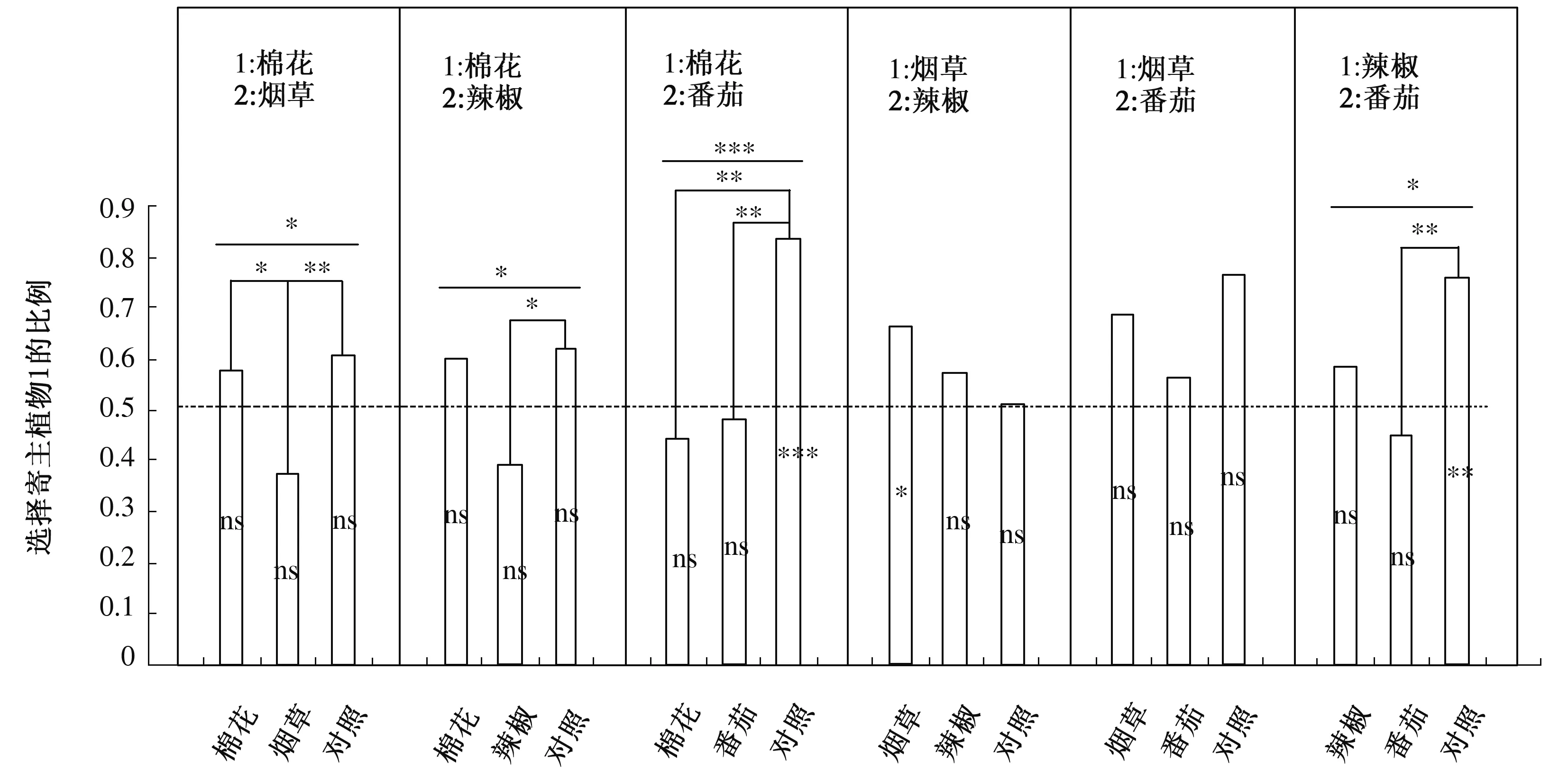
胚胎发育期经历的环境每个亚组包含3个组的棉铃虫(寄主1粗提物处理组、寄主2粗提物处理组和对照组)对寄主1和寄主2的选择频次,纵轴表示各处理组对寄主1的选择比例。组间差异(柱条上部)和组内差异 (柱条内部)采用的统计方法为卡方测验。ns表示特定处理组对寄主1和寄主2的选择频次没有显著的组内差异,*、 **、***分别表示在P=0.05、P=0.01、P=0.001的水平上有显著的组内差异或组间差异。为简化制图,组间无显著差异的未标注
3 结论与讨论
本研究通过测试棉铃虫的卵在4种不同寄主化学刺激物环境下发育对随后孵化幼虫嗅觉偏好性的影响,发现了一个明显的规律:凡是在烟草刺激物环境下发育的个体,无论与何种寄主植物配对比较,对烟草挥发物的趋性总是上升;另外2种茄科的寄主植物番茄和辣椒也有诱导正面经历效应的功能,但是弱于烟草的诱导作用。
昆虫虫态与虫态之间的联系有重要的生理、生态功能。例如,某个发育阶段经历的环境条件及其生理表现也会影响大龄阶段的适合度,这是表型可塑性的一种形式[8]。卵发育经历效应的研究不仅有很强的理论意义,也有很好的应用价值。从理论上讲,卵阶段是完全变态类昆虫寄主范围拓展或者寄主转移生态过程的关键环节,因此本研究有助于从世代发育链的角度理解昆虫经历效应或者表型可塑性。然而,此前的研究主要关注的是雌成虫的产卵偏好性与其后代生长表现的关系[16],或者幼虫取食经历对随后成虫产卵选择反应的影响[6]。从实践上讲,杀虫剂防治鳞翅目食叶类害虫的最佳时期是低龄幼虫阶段,而卵则对常见的杀虫剂耐性较强。在杀虫剂喷雾不均匀的情况下,卵发育期间经历的化学环境可能对防治效果有正面或者负面的作用,具体取决于经历效应的方向。
本研究提供了卵期发育的环境能够影响初孵幼虫寄主趋向选择的详细证据。与人工饲料饲养的棉铃虫个体相比,卵阶段经历的所有寄主植物刺激物都会显著改变初孵幼虫的嗅觉偏好性,而且通常是促进了对所经历寄主植物的选择。推测这是由于卵壳表面携带的微量寄主植物化学刺激物质渗透到卵内部造成的结果。在涉及番茄气味源的比较中,发现一个有趣的现象:无经历对照组对番茄粗提物涂布滤纸碟的选择频次总是比涂布其他3种寄主植物粗提物的滤纸碟低。推测这是由于番茄挥发的气味对无经历对照组的初孵幼虫有驱避作用造成的,而不是其对其他3种植物的嗅觉偏好性太强所致。据报道,棉铃虫在寄主范围内植物上的取食经历可以诱导出对相应植物的取食偏好性,但是如果用麦胚基人工饲料饲养3代以上,这些昆虫的取食偏好性就会回归到祖先寄主(棉铃)上[17]。这种“返祖”现象似乎能够解释番茄气味源 vs. 棉花气味源的测试结果。然而,没有证据表明烟草和辣椒也是棉铃虫的祖先寄主[14,18],但在烟草 vs. 番茄以及辣椒 vs. 番茄的测试中也得到类似的结果,这就说明麦胚基人工饲料的饲养过程造成了棉铃虫初孵幼虫对番茄气味的回避行为。
除了番茄经历组之外,卵期暴露于其他3种寄主植物化学刺激下也能削弱或者抑制人工饲料饲养的无经历对照组个体对番茄挥发物的强烈回避反应。因此,对番茄挥发物的嗅觉习惯化不能解释本研究的结果,应当更多地注意寄主经历效应的对立面。也就是说,当前多数学者是用麦胚饲料纯化昆虫的取食背景,但本研究发现这种饲养过程不仅消除取食背景对生物测定结果的干扰,而且代表着一种新的经历。貌似无关的因素如幼虫环境、饲料或者笼子的颜色也有可能导致试验结果之间的差异,而这一点被错误地解释为试验设计处理之间的差异,最终导致不同饲养来源的同种被试昆虫得到的结果缺乏可重复性[19]。例如,在缺乏气味的人工饲料上饲养的黑腹果蝇(Drosophilamelanogaster),和补偿气味的相同人工饲料饲养的个体相比,嗅觉接受神经元的电脉冲图案有很大的差异,具体表现为饲养过程中剥夺气味感觉的个体嗅觉敏感度和锐度下降[20],类似的结果也在人工饲养的黑豆蚜(Aphisfabae)中发现[21]。如果这一点在本研究中是事实,期望结果是用人工饲料饲养数代的棉铃虫幼虫对嗅觉驱避剂不再敏感,然而本研究中发现了相反的现象:人工饲料长期饲养造成棉铃虫对番茄挥发物的回避行为更强。推测这是一种典型的嗅觉敏感化现象,和黑腹果蝇及黑豆蚜的情况相反。为了确定这种驱避性的化学基础,未来有必要进一步研究番茄叶子释放出的挥发物成分,筛选出有行为活性的单体或混合物。
