高温胁迫对马铃薯幼苗生长和部分生理指标的影响
2018-09-26戴鸣凯张志忠樊佳茹王革伏
戴鸣凯,张志忠,刘 爽,樊佳茹,王革伏,3
(1福建农林大学园艺学院,福州350002;2福建农林大学-戴尔豪西大学联合实验室,福州350002;3戴尔豪西大学农学院,植物食品和环境科学系,加拿大新斯科舍B2N5E3)
0 引言
高温是农业生产中面临的重要问题,随着全球气候变暖,温度逆境对农作物生产影响越来越大。马铃薯属茄科多年生草本植物,为粮菜兼用型作物,目前已成为继小麦、水稻和玉米之后的第四大作物,中国是马铃薯第一生产大国,在粮食安全方面马铃薯扮演着重要角色。高温会严重抑制马铃薯生长发育、产量形成和品质,严重时可导致大幅减产,造成经济损失[1-3]。进行高温胁迫对马铃薯的影响研究具有较为重要价值。高温胁迫可对植物多方面产生不良影响,模拟大气增温田间试验表明高温胁迫可使马铃薯生理生态和产量形成过程发生了显著变化[4],高温会减少马铃薯块茎数目及大小[5],块茎形成过程受到高温抑制可能是影响其生物量及产量的原因[6-7]。采用数学和计算机建模分析表明随着气候变暖,气温升高,2080—2100年期间南亚区域印度的马铃薯产量将降低10%~40%[8]。研究表明,高温胁迫会打乱植物生理代谢,损害细胞结构,然而面对胁迫植物会启动自身的保护机制来减少受到的伤害[9-10]。试验以马铃薯品种‘闽薯1号’为材料,研究了持续高温胁迫下马铃薯幼苗植株生长情况变化,超氧阴离子(O2-)、丙二醛(MDA)、过氧化氢(H2O2)和脯氨酸(Pro)的含量以及超氧化物歧化酶(SOD)、过氧化酶(POD)、过氧化氢酶(CAT)等酶活性的变化,研究结果将有助于深入理解高温逆境对马铃薯生长发育和生理生化变化的影响。
1 材料与方法
1.1 植物材料
‘闽薯1号’(费乌瑞它×大西洋,高产、抗病、适应性强,商品薯率高、食用品质好,无裂薯、薯型椭圆),由福建省农业科学院作物研究所提供。
1.2 试验方法
1.2.1 种薯处理 置于人工气候箱避光,气温20℃(恒温),湿度30%,放置2周,促进顶芽生长,待顶芽长至豆粒大小,温度降至12℃(恒温),并给予光照,光照强度为144µmol/(m2·s),抑制顶芽生长,促进其他芽眼萌芽。
1.2.2 材料处理 马铃薯块茎切块,每个薯块质量控制在50 g左右,在花盆中以草炭土、蛭石和珍珠岩按照3:1:l的比例培养,每盆1株,在自然温光条件下室温培养至株高15 cm左右,选择生长相对一致的幼苗移至人工气候箱,调节光照强度为144µmol/(m2·s),空气湿度为75%,光照16 h,暗培养8 h;3个温度处理:22/14℃作为对照,35/27℃、39/31℃作为高温处理,每一个处理放置15株生长相对一致的幼苗,以0、1、3、5、7天为取样时间点,每个温度同一时间点取3株幼苗,每株取第3~5叶位展开的生长相对一致的功能叶2片,为减小试验误差叶片去掉叶脉并剪碎混样,单个指标每个样品取0.1 g,使用锡箔纸包裹,立即使用液氮充分冷冻,使用塑封袋放置-80℃低温保存以测定生理指标,取样前拍摄照片并记录植株生长状况。所有试验进行3次重复。
1.2.3 生理指标测定 超氧阴离子(O2-)含量采用苏州科铭生物技术有限公司生理指标测试试剂盒测定:0.1 g组织样品添加1 mL提取液进行冰浴匀浆,然后1000 g,4℃,离心20 min,取上清置冰上待测,按测定操作表依次加入指定试剂,混匀,8000 g,25℃,离心5 min,吸取上层水相1 mL,加入1 mL玻璃比色皿,分光光度计预热30 min以上,波长调节至530 nm,空白调零,测定A530,按样本质量计算超氧阴离子含量(umol/g鲜重),如式(1)所示。
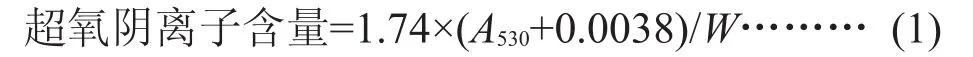
丙二醛(MDA)、过氧化氢(H2O2)和脯氨酸(Pro)的含量以及SOD、POD、CAT的酶活性均采用苏州科铭生物技术有限公司生理指标测试试剂盒测定(样品提取、测定及数据计算)。
1.2.4 数据处理 数据采用WPS Excel和DPS统计分析软件包进行处理,以单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)分析不同胁迫温度下生理指标的差异。试验在福建农林大学-戴尔豪西大学联合实验室,于2017年9月—2018年1月进行。
2 结果与分析
2.1 植株生长表型随胁迫时间的变化
高温胁迫明显了影响马铃薯幼苗生长发育(图1),35℃与39℃高温处理的第0、1天,植株还未表现出生长的异常,从第3天起,2个高温处理下的植株生长缓慢,叶片萎焉,22℃常温处理下的植株生长良好;观察发现(见图2a、b)35℃下第5天的马铃薯幼苗叶片不仅短小,且叶片、叶柄(见图2c)发生明显的卷曲,顶叶与侧叶的大小也小于常温对照,图3显示幼苗叶片长度在常温与高温下存在显著性差异,常温下幼苗叶片生长速度高于高温处理,35℃与39℃高温处理下叶片生长缓慢,尤其是39℃高温,呈现出非常缓慢的生长速率,叶片长度显著小于35℃。
2.2 保护酶活性随胁迫时间的变化
植物在受到环境胁迫时,通过调节体内的SOD、POD、CAT等抗氧化酶的活性来清除体内积累的活性氧,以减轻植物体受到来自活性氧的伤害。高温胁迫对马铃薯幼苗SOD、POD、CAT活性的影响如图4。

图1 不同温度下植株的生长表型记录

图2 高温下植株及叶片生长表型
SOD从第1天开始2组高温处理下,SOD的活性显著上升,其中第1天35℃处理下的SOD活性提高了约113%,39℃下提高约107%;但随着胁迫时间的延长,第3、5、7天高温处理下由于胁迫程度的加强,SOD的活性持续降低,常温处理一直处于比较稳定的状态。如图4(b)显示,35℃下POD活性于第1天显著上升,于第3、5天下降,又于第7天上升,而39℃时,POD在第1天就出现了降低的趋势,随着胁迫的加剧,活性又缓缓升高;与SOD类似,35℃高温下的活性要高于39℃。如图4(c)显示,高温处理第1天CAT活性显著降低,35℃CAT活性下降约31.7%,39℃更是下降约55.8%,但随着胁迫时间的延长,CAT活性又逐渐升高,与SOD、POD不同的是,39℃下CAT活性的上升更为显著。总而言之,受到高温胁迫,SOD,POD,CAT活性均有显著变化,其中SOD活性受到胁迫第一时间显著上升,随着胁迫时间的延长而缓慢降低,而POD与CAT表现出大致先下降而后升高的趋势。
2.3 MDA含量随胁迫时间的变化
高温胁迫会加剧植物细胞膜的膜质氧化,而MDA是膜质氧化产物之一。如图5,35℃和39℃高温处理植株的MDA含量随着胁迫时间的延长而逐渐增长,表明植株叶片膜质氧化程度正在逐渐增强,常温处理则无显著变化;同时35℃与39℃高温处理之间也具有显著性差异,可以发现39℃下植株的MDA含量要显著高于35℃。SOD是清除氧自由基缓解植物膜质过氧化的主力军,尽管SOD活性大大提高如图4(a),MDA含量依旧在持续增多,可见随着胁迫的增强,通过植物体自身来缓解伤害的能力还是很有限的。
2.4 O2-和H2O2含量随胁迫时间的变化
O2-是生物体受到胁迫后首先生成的氧自由基,可以经过一系列反应而生成其他氧自由基,参与启动膜质过氧化[11],如图6(a),35℃与39℃高温下O2-含量相对于常温的增长并不显著,其中第5天2个高温下的O2-含量显著高于常温且39℃显著高于35℃。SOD在清除超氧阴离子的同时又会有H2O2生成,如图6(b)显示,从第3天开始到第7天,H2O2含量持续增加,到第7天为止35℃下H2O2含量相比于0天增加了约79.4%之多,39℃增加了约64.6%。CAT是参与清除H2O2的主力军,虽然CAT的活性随着胁迫的持续活性逐渐提高如图4(c),但H2O2含量依旧没有降低,这可能是因为胁迫所产生H2O2的速度超出了CAT的清除速度,导致植株来不及清除体内不断产生的H2O2,以至于H2O2不断积累增多。

图3 叶片长度随胁迫时间的变化
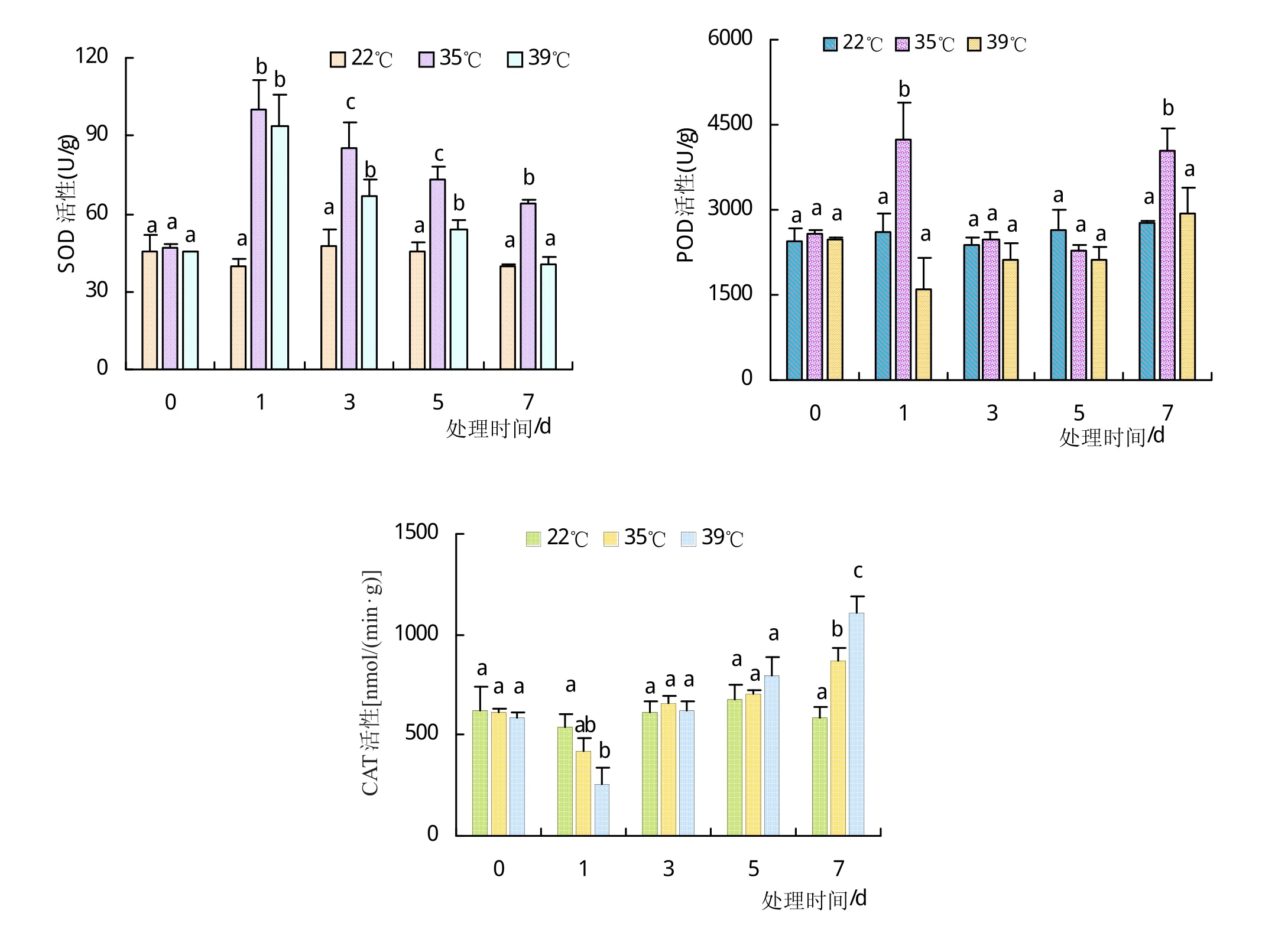
图4 保护酶活性随胁迫时间的变化
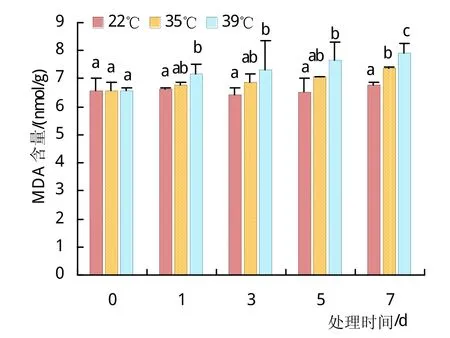
图5 MDA含量随胁迫时间的变化

图6 O2-和H2O2含量随胁迫时间的变化
2.5 叶片Pro含量随胁迫时间的变化
脯氨酸是植物的主要渗透调节物质,一般认为高温胁迫下植物体内的Pro含量会显著增加[12],如图7,Pro含量从第1天开始持续上升,且在39℃高温下积累的量要高于35℃;35℃与39℃处理下在第1、3天出现了相似的升高幅度与趋势,但从第5天开始,39℃下植株的Pro含量增幅要显著高于35℃,第5天39℃下Pro含量相对于常温增长了约44.4%,35℃下仅增加了约18.3%,第7天39℃下Pro含量相对于常温增长了约49.1%,35℃下仅增长了约18.8%;说明随着高温胁迫的持续和加强,会使马铃薯幼苗体内产生并积累更多的脯氨酸;常温对照组脯氨酸含量则无显著变化。
3 讨论
植物受在高温胁迫下,其形态结构及生理生化均会受到一定程度的影响。尤超等[13]以油桃为材料,发现在33℃高温下嫁接苗初期几乎不受影响,仅在第三天出现少部分叶片卷曲,但在39℃和45℃下,随着受害指数的增加,叶片出现卷曲、皱缩、变软的概率大大上升,成正相关,与本实验的结果十分相似。研究表明高温会使植物体内累积更多活性氧,而植物体内清除活性氧的酶类主要有SOD、POD、CAT等,SOD可以催化超氧阴离子的歧化反应而产生氧气和过氧化氢,大大减少超氧阴离子的含量从而减轻其对植物的伤害;与此同时产生的H2O2可产生氧化能力极强的羟基自由基和单线态分子氧,这将对植物细胞造成很大的伤害,因此POD和CAT将会进一步清除产生的H2O2从而保护植物体[14-15]。本试验中,马铃幼苗受到高温处理时,SOD在高温初期活性大幅提升,而随着高温的持续,活性持续降低,这可能与高温的持续在一定程度上破坏了酶的结构有关,也有相关研究在乌菜幼苗和大白菜上发现了同样的规律[16-17];本试验中POD、CAT活性与SOD活性不同,出现了先降低后升高的趋势,这可能与初期高温胁迫了植物体抑制了酶的活性有关,同时SOD清除超氧阴离子时产生了H2O2,H2O2积累再一次胁迫植物体,植物体便逐渐提高了POD与CAT活性以保护自身。其中35℃时,POD活性在第1天有显著提高,可能是由于35℃对植株未构成足够强胁迫,同时在初期伴随着一定量H2O2的产生,植株本能的提高了其活性,而随着高温的持续,胁迫加重,POD活性又在第3天降低,到后期H2O2大量积累,植株通过提高POD活性来应对胁迫,叶凡等[17]也在不结球白菜上发现了同样的规律。这几种重要的抗氧化酶适应高温胁迫的特点不同,对高温胁迫的反应也有一定的差异。
膜质过氧化是高温对细胞膜伤害的一种表现,MDA是膜质过氧化的产物,也是细胞膜被破坏的标志物质。研究表明,高温处理下,大多数植物的MDA含量都表现为增加的趋势[19],与本试验结果一致。马德华利用黄瓜幼苗获得了MDA含量与高温胁迫的正相关[20];孙保娟等人研究了番茄在持续高温下MDA含量的变化,也得到了一样的结果。超氧阴离子和H2O2是引发膜质过氧化的主要物质,随着高温胁迫的延长,其含量都会逐渐积累,对植物体造成进一步的伤害。脯氨酸(Pro)是重要的渗透调节物质,其调节作用被广泛研究,研究表明,高温胁迫下植物体内Pro含量会明显增加[12]。李威等[21]在茄子耐热性研究时,发现耐热性与脯氨酸含量的关联达到显著水平;另外耐热性强的水稻品种在高温胁迫下游离脯氨酸的增加幅度更大[22]。
综上所述,高温胁迫对马铃薯幼苗生长与生理生化代谢都造成一定程度的影响,植株生长萎焉、叶片生长缓慢,同时加剧植株水分丧失,膜质过氧化加剧,活性氧含量增加,导致叶片卷缩,甚至出现了损伤。马铃薯植株通过调节体内保护酶活性、提高脯氨酸含量等手段来缓解持续的高温胁迫。

图7 Pro含量随胁迫时间的变化
