秦川牛TRAF6基因多态及其与体尺性状的关联分析
2018-09-21张芷毓欧德琼邬明丽樊英智高源李世鹏赖振雨雷初朝党瑞华
张芷毓,欧德琼,邬明丽,樊英智,高源,李世鹏,赖振雨,雷初朝,党瑞华
(西北农林科技大学动物科技学院,陕西 杨凌712100)
TRAF6(TNF receptor associated factor 6)基因,是肿瘤坏死因子超家族(TNF)、Toll样受体家族(TRLs)和白细胞介素1受体超家族(TIR)信号通路调控途径中关键接头分子,是天然免疫信号通路的重要接头分子[1]。与此基因相关的功能包括功能性破骨细胞的形成与分化,和骨肉瘤细胞的增殖、凋亡和侵袭,一些研究已经强调该基因对小鼠的出生前和出生后存活率的重要影响,在TRAF6基因中诱导突变时,突变纯合的小鼠在出生时表型正常,但却过早地死于骨硬化,少数存活动物的体重也比健康动物减少20%~30%,体长也较短。这一证据表明,TRAF6基因可以参与牛早期发育阶段的调节,因此可以对牛的性状产生影响。位于该区域的数量性状位点与荷斯坦牛的产犊和Balman牛的日增重(ADG)相关[3]。
已知TRAF6基因可以在多种动物的多种类型细胞中表达,TRAF6的表达产物,肿瘤坏死因子受体相关因子6,是天然免疫信号通路中重要的接头分子。研究表明,人和模式生物小鼠等动物的TRAF6可调节多种细胞信号通路[4-5]。在免疫应答,炎症反应,应激反应和细胞分化与凋亡等生物学过程中起着重要作用[6-8]。目前对TRAF6的功能研究多见于人,小鼠,家禽及水生动物,这些研究表明TRAF6除具有抗病毒功能之外,可能还参与针对多种类型疾病的免疫应答,其中某些突变型还与类风湿性关节炎的诱发有关[9]。牛TRAF6基因位于15号染色体上,含有七个外显子,编码902个氨基酸,目前关于牛TRAF6基因多态性及抗病毒活性的研究主要集中在国外具有代表性的牛品种上。目前,对于TRAF6基因在各种动物中发挥的各种不同功能的研究尚在不断进行中,例如是否对猪的伪狂犬病病毒存在抑制效应等[2]。本试验旨在通过分析陕西地区秦川牛TRAF6基因的多态性,为下一步调查其多态性与牛体尺的关联性奠定基础,也为将来TRAF6基因在肉牛抗病育种中的应用提供基础资料。
1 材料与方法
1.1 材料
1.1.1 血样的采集 从陕西省秦川牛场、陕西省秦川牛良种繁育中心随机选取年龄、胎次及产犊时间相近;放牧、饲养和管理条件相同的健康秦川牛254头,从牛颈静脉采血20~40 mL/头,加ACD抗凝剂2 mL(V(ACD):V(血液)=1:6),置于冰壶中带回,-80℃保存。
1.1.2 主要试剂和仪器 蛋白酶K购自德国默克公司;含染料Reaction MIX、DNA Marker、去离子水;Tris饱和酚、EDTA、Tris、甲醛、硼酸、丙烯酰胺、N,N’-亚甲基双丙烯酰胺(Bis)、过硫酸铵、Temed、二甲基亚砜(DMSO)、N,N,N,N-四甲基乙二胺;十二烷基磺酸钠(SDS)、去离子甲酰胺、琼脂粉;HincⅡ酶。
紫外分光光度计、冷冻高速离心机(CENTURION,英国)、PCR仪、电泳槽、稳流稳压电泳仪、脱色摇床、凝胶成像系统(Bio-Rad公司,美国)。
1.2 秦川牛体尺及肉质性状的测量
实地测量所采血样秦川牛生长性状数据(体高、胸围、胸宽、胸深、腰高、体重、十字部高、体斜长、臂长、腰角宽、腹围)。
1.3 秦川牛基因组DNA的提取
采用传统的酚氯仿抽取法提取秦川牛的基因组DNA,提取后用1%的琼脂糖凝胶电泳检测DNA,用紫外分光光度计测定DNA样品的浓度,检测完毕后,取一定量的DNA样品稀释至50 ng/μL,保存于﹣20 ℃备用。
1.4 秦川牛TRAF6基因的PCR-RFLP分析
1.4.1 引物的设计与合成 参照GenBank提供的牛TRAF6基因序列(NC_007313),利用primer 5.0软件设计引物,引物由上海生工Sangon Biotech公司提供,引物信息见表1。

表1 牛TRAF6基因的引物信息
1.4.2 TRAF6基因的PCR扩增 PCR反应体系12.5 μL:6.25 μL康维Mix(包括PCR稳定剂和增强剂、Mg2+、dNTPs、Taq DNA Polymerase、PCR Buffer),4.75 μL ddH2O,0.5 μL上游引物F,0.5 μL下游引物R,0.5 μL DNA(50 ng·μL-1)。
PCR反应程序:95 ℃预变性5 min;94 ℃变性30 s,54 ℃退火30 s(退火温度见表1),72 ℃延伸30 s,36个循环;最后在72 ℃充分延伸10 min,4 ℃保存。PCR扩增产物用1%琼脂糖凝胶电泳30 min检测。
1.4.3 PCR产物测序 随机选取30个秦川牛DNA样本,每个样本取样2 μL,用移液器轻轻吸打混合均匀,构建DNA混池进行PCR扩增,PCR反应体系25 μL:12.5 μL康维Mix(包括PCR稳定剂和增强剂、Mg2+、dNTPs、Taq DNA Polymerase、PCR Buffer),9.5 μL ddH2O,1μL上游引物F,1 μL下游引物R,1 μL DNA(50 ng·μL-1)。扩增产物送至上海生工生物工程股份有限公司进行测序。测序结果使用SeqMan进行对比寻找突变位点。
1.4.4 PCR产物酶切及PCR-RFLP检测 酶切体系10.1 μL:TRAF6基因的PCR扩增产物5 μL、10×Buffer 1 μL、ddH2O 4 μL、限制性核酸内切酶HincⅡ0.1 μL。该酶切反应体系在37℃恒温箱中酶切4 h。取酶切产物2.5 μL,12%聚丙烯酰胺凝胶在 120 V电泳1.5 h。
1.5 数据统计分析

ym=μ+Markerm+em
式中,ym为个体表型值,μ为群体均值,Markerm是标记基因型效应,em为随机误差。
2 结果与分析
2.1 秦川牛TRAF6基因突变区域的PCR扩增结果
利用合成引物扩增样本牛TRAF6基因突变区域,所得PCR产物用1%琼脂糖凝胶电泳30min检测。如图1显示,扩增片段与目的片段一致,且条带清晰、特异性好,可直接用于后期的DNA测序。

图1 秦川牛TRAF6基因突变区域的PCR扩增结果
2.2 秦川牛TRAF6基因的测序及测序结果分析
使用SeqMan软件,分析秦川牛TRAF6基因突变区域的测序结果,发现了一个多态位点,见图2。对比分析结果表明:第六内含子17628 bp处发生由T到C的突变,将其命名为T17628C(B)。分析表明,B位点存在2种基因型TT和TC。由于该处突变发生在TRAF6基因的第六内含子区域,故未引起相应氨基酸的突变。
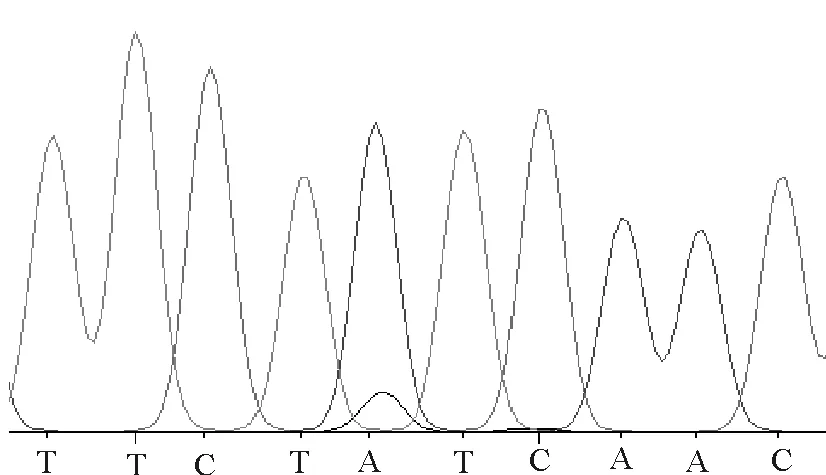
TC型
2.3 秦川牛TRAF6基因突变区域PCR-RFLP分析
PCR-RFLP分析结果表明,秦川牛TRAF6基因第六内含子区域存在TT、TC两种基因型,如图3显示,序列经酶切后应存在三条条带,但因为其中一条带较短只有38 bp,故电泳图谱上仅能体现其中较长的两条。HincⅡ酶切位点如图4显示。
图3 秦川牛TRAF6基因突变区域PCR-RFLP电泳图

图4 HincⅡ酶切位点
2.4 秦川牛TRAF6基因突变区域遗传多样性分析
如表2显示,从群体遗传学角度分析秦川牛TRAF6基因各突变位点的基因型频率、等位基因频率、纯合度、杂合度、有效等位基因数、多态信息含量等遗传指标。χ2结果表明,B位点 TT是优势基因型,T为优势等位基因,位点处于低度多态状态。检验表明,P﹥0.05,B位点处于Hardy-Weinberg平衡状态。

表2 秦川牛TRAF6基因B位点突变的遗传多样性
注:括号中的数据表示检测个体数。
2.5 秦川牛TRAF6基因突变位点不同基因型与体尺的关联性分析
如表3显示,运用SPSS(22.0)软件分析169头秦川牛不同基因型与体高、胸围、胸宽、胸深、腰高、体重、十字部高、体斜长、臂长、腰角宽、腹围的相关性。
由表3可知,TRAF6基因B位点不同基因型在体高、胸围、胸宽、腰高、体重、十字部高、体斜长、臂长、腰角宽、腹围方面无显著性差异,TT基因型的个体均值均高于TC基因型的个体均值,但无统计学差异(P>0.05)。T检验结果显示B位点不同基因型在腰高、十字部高、体斜长的P值分别为0.066、0.077、0.063,均大于0.05,在一般公认的统计意义上不显著,但非常接近显著。
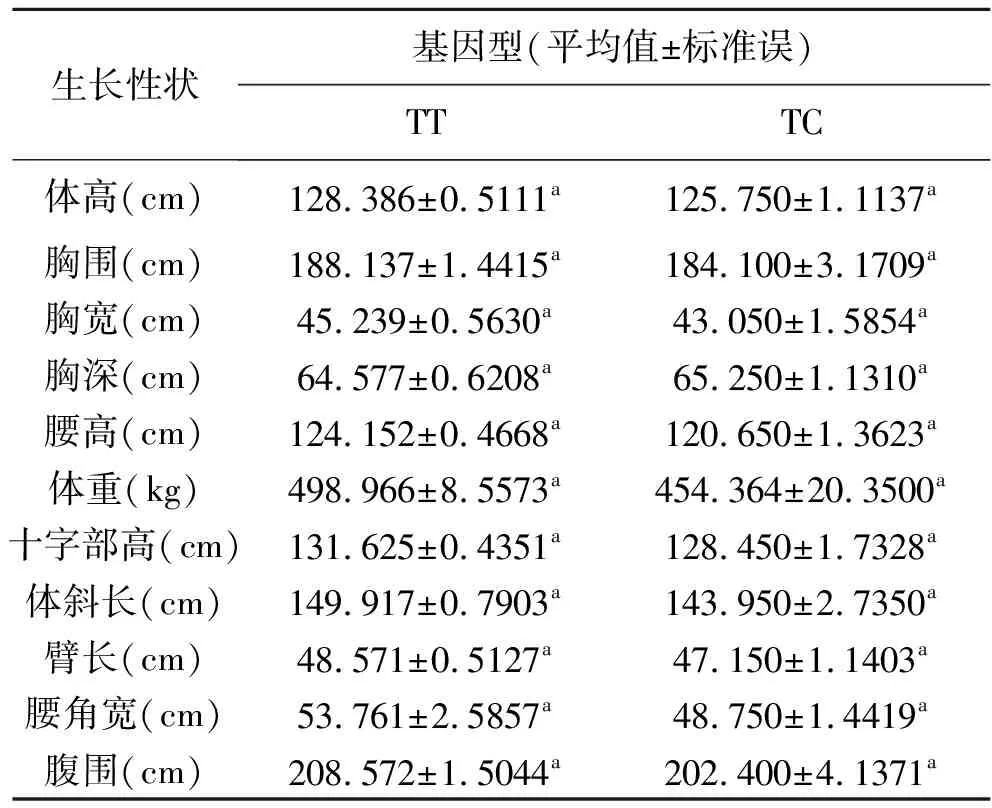
表3 秦川牛TRAF6基因各突变位点不同基因型与体尺性状的关联性分析
注:表中数值为平均值±标准误;同列不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01)
3.讨论
TRAF6基因不仅存在于脊椎动物中,比如猪牛等禽畜,也存在于许多非脊椎动物如青蛤[12]中,以及半滑舌鳎等鱼类[2.11]。说明TRAF6基因分布广泛,对机体生存具有重要作用。目前对于TRAF6基因的研究主要集中在其表达情况对于人类疾病的影响,关于其对禽畜健康及生长性能的影响的研究报道较少,且集中在对其编码区的克隆及序列分析工作,及其对密码子使用的偏好分析等工作上。已知牛TRAF6基因分别含有7个外显子与6个内含子,而人TRAF6基因有 2 个转录突变体(NM_145803.2)(NM_004620.3)与基因组DNA(NC_000011.10)发现所对应的 DNA分别含有 8个外显子和 7个内含子,可见牛和人TRAF6基因的基因结构存在较大差异,不可一概而论[4.8]。
针对TRAF6基因外显子设计引物,扩增产物通过PCR-RFLP检测分析,均未发现多态性,而扩增TRAF6基因内含子的TRAF6-4扩增片段在第6内含子17628bp处检测到T→C突变,可分为TT、TC两种基因型。该TRAF6基因多态性位点处于哈代-温伯格平衡状态,说明该群体在环境选择、人工选育等因素的影响下,其遗传结构并未发生改变,处于一种动态平衡状态。
本研究运用PCR-RELP方法,首次对秦川牛5’调控区,CDS区及部分突变频率高的内含子区域进行SNP筛查,利用PCR-RFLP技术在突变位点附近发现HincⅡ-RFLP酶切位点,使用HincⅡ酶对扩增条带进行切割,当此位点的T突变为C时可被HincⅡ酶切开,利用电泳技术可进行分型。突变位点多态性较丰富,关联分析显示该位点的杂合突变对牛体尺及体重等生长性状存在影响,因此可以作为遗传标记辅助选择的参考。
4 结论
陕西地区秦川牛的TRAF6基因存在较丰富的遗传多态性,在第六内含子17628 bp处检测到了SNP位点,形成两种基因型,可能作为一种遗传标记作为辅助选择的参考,对指导肉牛的育种工作具有实践意义。
