模拟酸雨和镉复合处理对秃瓣杜英生长和荧光参数的影响
2018-09-19项敬银伊力塔陈敏斯
项敬银,俞 飞,伊力塔,陈敏斯
浙江农林大学省部共建亚热带森林培育国家重点实验室, 临安 311300
随着经济社会的快速发展,酸雨污染已成为中国重要的环境问题[1]。目前,酸雨污染面积已占中国国土面积的40%,特别是华东地区已成为中国酸雨形势最严重区域之一,酸雨频率及酸雨覆盖面积不断增长和扩大[2- 3]。临安大气本地污染监测站的数据显示:2005—2010年,大气本地站降水各月pH均值均小于4.5,达强酸雨程度,且酸雨pH值总体呈逐年下降趋势[4]。相关研究指出,酸雨会破坏叶片角质层、表皮组织和气孔[5],改变植物体内酶活性而影响光合作用[6],抑制植物养分、水分的吸收和运输导致植物衰退[7-9]。除此之外,由于工业生产的迅猛发展以及农田污灌,大量镉进入生态系统,镉因其化学活性强、移动性大、毒性持久,严重危及动植物生存和人类健康[10-11]。目前在中国受到重金属污染的耕地面积近有2000 万hm2,约占总耕地面积的1/5[12]。杨楠楠[13]发现镉、汞、铅等元素严重超标的地区主要集中在浙江、上海、南京等发达城市中心,且区内中强度级以上的污染占80%左右,并且在中心城市出现了较大面积的局部污染。镉作为对植物产生主要毒害作用的重金属污染物[14],是植物生长的非必需元素,可使植物出现生长受阻的现象,使植株生长、产量及其在体内积累受到影响[15-16]。然而自然状态下,绝对意义上的单一污染是不存在的,污染多具有伴生性和综合性[17],各因素间可能会产生加和、协同或拮抗等作用。酸雨可以影响土壤中重金属的化学行为,二者共同导致植物生存条件进一步恶化[18]。已有资料表明酸雨会造成土壤中有毒重金属元素活性明显提高,溶解性增加,迁移能力提高[19],不仅形成对植物有害的土壤环境,而且使植物地上部分的茎叶中重金属含量明显提高,改变植物的理化性质,降低植物的抗性和品质[20]。但张丽华等[21]发现模拟酸雨作用于受重金属污染土壤时,污染土壤中不同重金属呈现不同的溶出规律,且溶出量与模拟酸雨pH相关。古昌红等[22]发现在酸雨作用下,酸度的增加对污染土壤中Cd、Zn、Cu的释放影响不大。
秃瓣杜英,杜英科(Elaeocarpaceae)杜英属(Elaeocarpus)常绿乔木,亚热带典型地带性树种,在我国福建、浙江及云南等省(区)海拔400—750 m的常绿阔叶林和针阔混交林中广泛分布[23]。中等喜光,生长迅速,在中性、微酸性的山地红壤、黄壤上均可生长;不仅抗风力强,也是城市绿化、庭园观赏和四旁绿化的优良树种。前期有研究发现秃瓣杜英的生物量积累、光合能力、PSⅡ原初光能转化效率和PSⅡ潜在的活性在模拟酸雨处理下均有所升高[23-24]。俞飞等[25]也发现秃瓣杜英对酸雨胁迫有较强的适应能力。本研究旨在相关研究的基础上模拟酸雨和镉复合环境条件下,秃瓣杜英的生物量积累、镉分配差异和荧光参数的响应特征,探究复合污染对亚热带树木的响应机制。
1 材料与方法
1.1 试验地概况
试验地位于浙江农林大学东湖校区试验温室(119°44′E,30°16′N)。该地属中亚热带季风气候区,温暖湿润,光照充足,雨量充沛,具有春多雨、夏湿热、秋气爽、冬干冷的气候特征。年平均气温16.6 ℃,年降水日158 d,年降水量1454.1 mm,年日照时数1825.7 h,无霜期年平均237 d,土壤为红黄壤[7]。
1.2 试验材料与试验设计
供试材料为浙江省农业科学研究院蚕桑所提供的1年生同种源实生秃瓣杜英幼苗,于2016年5月采用当地红黄壤将120棵幼苗移栽至高40 cm、直径30 cm的塑料花盆中,每盆1株,将所有幼苗编号、挂牌,置于通风、遮雨的塑料薄膜受控大棚内。缓苗期常规管理,用蒸馏水浇灌;试验期间及时除草、浇灌,以免造成干旱。缓苗两个月待秃瓣杜英长势稳定后,选取株高和地径基本一致的幼苗随机分成9组,每组12株,共108株。
试验采用随机区组设计,3次重复。设计4种类别处理方式:单一酸雨处理,包括pH 3.0和pH 5.6;单一镉处理,包括Cd 50和Cd 100;酸与镉复合处理,包括pH 3.0+Cd 50、pH 3.0+Cd 100、pH 5.6+Cd 50和pH 5.6+Cd 100;以及对照处理:CK(pH 7.0+Cd 0)。一共9个处理。查阅资料得大多数植株镉浓度耐受范围均小于50 mg/kg,杜天庆等[14]发现小麦耐受镉浓度为300 μmol/L,甜高粱幼苗、向日葵、香蕉耐受镉浓度分别为12 mg/kg、100 μmol/L和30 mg/kg[26- 28],较少植株能耐受100 mg/kg的镉浓度,因此本试验设置Cd 50和Cd 100两个梯度。
2016年7月,往花盆中加入氯化镉溶液(CdCl2),每3天加1次,共6次,最终使土壤中镉浓度接近67.71 mg/kg或126.97 mg/kg。模拟酸雨用浓度98%、密度1.84 g/mL的浓H2SO4和浓度68%、密度1.4 g/mL的浓HNO3配制(体积比1∶1)。喷淋前用蒸馏水稀释成pH 3.0和pH 5.6的溶液[7]。依据临安市近10年各月平均降雨量确定每月模拟酸雨喷淋量[29],并平均分配到每周,每周喷淋2—3次,每次300 mL,直至2016年9月下旬试验结束。
1.3 测定方法
1.3.1 生物量测定
生物量测定:2016年9月,将试验植株的根、茎、叶三部分分开,先用自来水冲洗粘附于植物样品上的泥土和污物,接着用超纯水冲洗,最后用去离子水冲洗,在烘干前先在105℃下杀青5 min,然后在60℃下于烘箱中烘至恒重,称量各部分的生物量[30]。
1.3.2 镉含量测定
用万分之一分析天平精确称取植株各部分样品干粉各0.2000 g,加入2 mL MillQ水和5 mL HNO3,隔日加入1 mL H2O2(HNO3和H2O2均为优级纯),放入微波消解仪(CEM-MARS 6,美国)中消解,赶酸后再用3% HNO3定容至50 mL,静置后用SOLAAR M6型原子吸收光谱仪(美国热电)测定根中的镉含量,用电感耦合等离子体质谱仪(XSERIES 2 ICP-MS,中国)测定茎和叶中的镉含量[31]。
1.3.3 荧光参数测定
于2016年7—9月利用调制叶绿素荧光成像系统(IMAGING-PAM,WALZ,德国)在晴朗天气的9:00—11:00时,选择植株中上部成熟叶片(每株3片)进行叶绿素荧光参数的测定,每个处理随机选取3株。每个处理的荧光参数值取3株植株的平均值。
首先,暗适应:用仪器专用暗适应叶夹DLC- 8夹住待测叶片,使其暗适应20 min后测定荧光参数,Fo(初始荧光)在暗适应后以弱调制测量光(0.05 μmol m-2s-1)诱导产生,随后用强饱和脉冲(6000 μmol m-2s-1)激发产生最大荧光Fm,闪光2 s。
接着,光适应:在自然光下直接测定,在光化光打开之前,测定一次Fo和Fm′(光适应的样品打开饱和脉冲时得到的最大荧光产量),然后在自然光照下系列饱和脉冲启动,施加多次强饱和闪光脉冲(6000 μmol m-2s-1,脉冲时间2 s),记录每次光适应下任意时刻的荧光产量(Ft)和Fm′,每隔20 s 测定一次,直至脉冲终止,取最后6次闪光的平均值(因闪光10次后基本达到稳态)。测定指标包括Fo、Fm、Ft,其中光系统Ⅱ最大光化学效率(Fv/Fm)、非光化学淬灭系数(NPQ)和光化学淬灭系数(qP)的计算公式如下:
Fv/Fm= (Fm-Fo) /Fm
NPQ = 1 - (Fm′-Fo′) / (Fm-Fo)
qP= (Fm′-Ft) / (Fm′-Fo′)
1.4 数据处理
利用SPSS 19.0统计分析软件中单因素方差分析对所有数据进行正态性和齐性检验,并采用Tukey或Duncan多重比较检验法进行差异显著性检验(P<0.05);利用双因素方差分析进行酸雨强度和镉浓度的影响进行检验;采用Excel绘制图表。
2 结果与分析
2.1 生物量积累与分配
从表1可以看出,单独的酸雨处理显著(P<0.05)增加了秃瓣杜英的总生物量积累。其中,pH 3.0和pH 5.6处理下的生物量与对照组相比分别提高了30.58%和16.31%,随着酸雨强度增加,植株的总生物量积累显著(P<0.05)增加,且根冠比提高。结合图1,酸雨处理改变了生物量的分配,pH 3.0促进了植株叶片生物量的分配,叶所占总生物量的比例较对照提高了40.40%;pH 5.6促进了植株根生物量的分配,根所占总生物量的比例较对照提高了37.73%,两者茎占生物量的比例较对照组显著(P<0.05)下降。
单独Cd处理下,Cd 50总生物量与对照组相比显著(P<0.05)增加,提高了约20.37%,结合图1,Cd 50促进了植株根及叶片生物量的分配,降低了茎干生物量的分配。而Cd 100的总生物量积累显著(P<0.05)低于对照组,但生物量分配并未受太大影响。随着Cd浓度的升高,植株根生物量、叶生物量和总生物量的积累均显著(P<0.05)下降。
复合处理下,pH 5.6+Cd 100显著(P<0.05)增加了总生物量积累,与对照组相比提高了48.84%,其余处理方式总生物量与对照组相比差异不显著(P>0.05)。所有处理均促进了植株根及叶片生物量的分配,降低了茎干生物量的分配(图1),植株的根冠比也均下降。
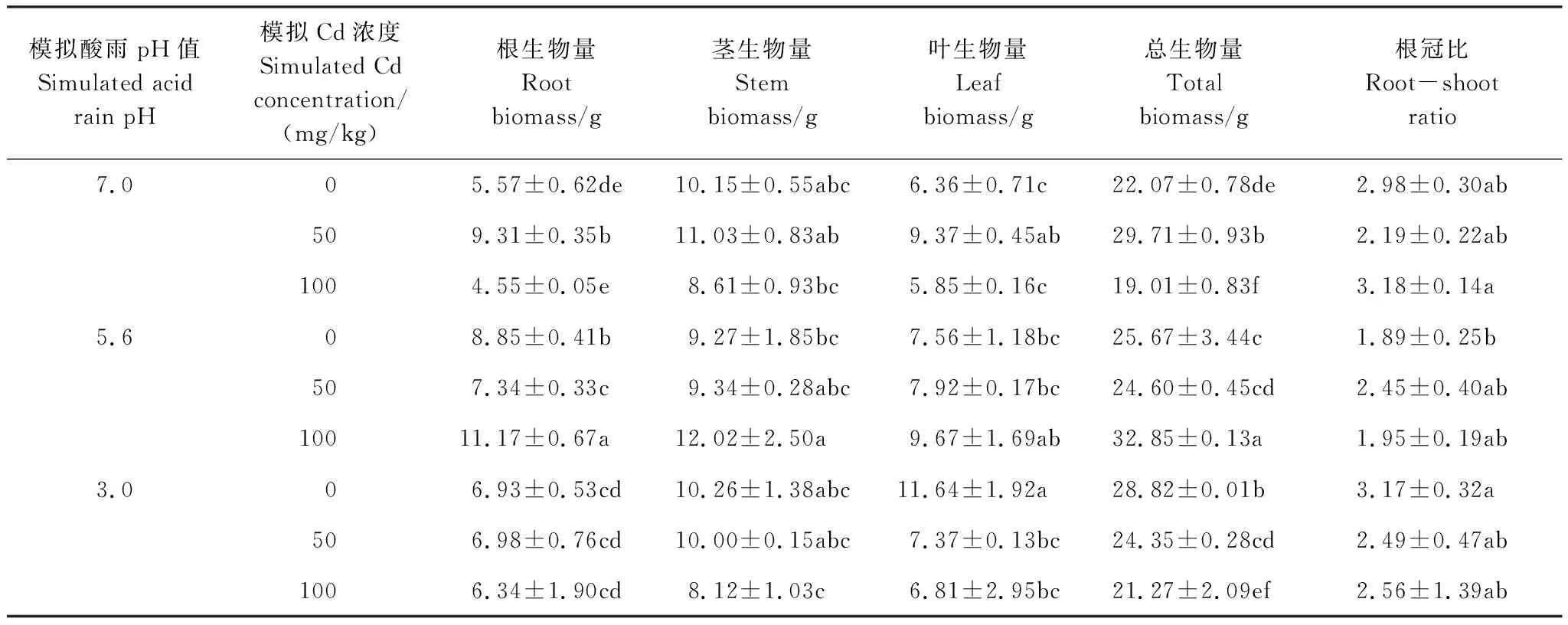
表1 酸雨和镉复合处理下秃瓣杜英生物量积累
注:表内数据为平均值±标准误差;同列不同字母表示差异显著(P<0.05)

图1 复合处理下秃瓣杜英生物量分配 Fig.1 The effects of simulated acid rain and Cd on the biomass allocation of Eleocarpus glabripetalus
2.2 镉积累
从表2可以看出,单独酸雨处理下,植株总的镉含量与对照组无显著差异(P>0.05),但显著降低了叶中的镉含量,特别是pH 3.0和pH 5.6处理中叶的镉含量与对照组相比分别减少了64.06%和32.81%。随着酸雨强度的增强,植株根、茎、叶及总的镉含量均下降,pH 3.0条件下最明显。
单独的镉处理必将显著增加植株器官的镉含量的积累,Cd 50和Cd 100总的镉含量分别是对照组的19.67倍和26.33倍。随着镉浓度的升高,植株根、茎、叶及总的镉含量均显著增加(P<0.05),Cd 100总的镉含量与Cd 50相比增加了33.86%。
而在复合处理下,与对照组相比,4种处理方式均显著(P<0.05)增加了植株根、茎、叶及总的镉含量的积累。pH 3.0+Cd 50、pH 3.0+Cd 100、pH 5.6+Cd 50和pH 5.6+Cd 100总的镉含量分别是对照的16.80、25.24、22.42和16.31倍。
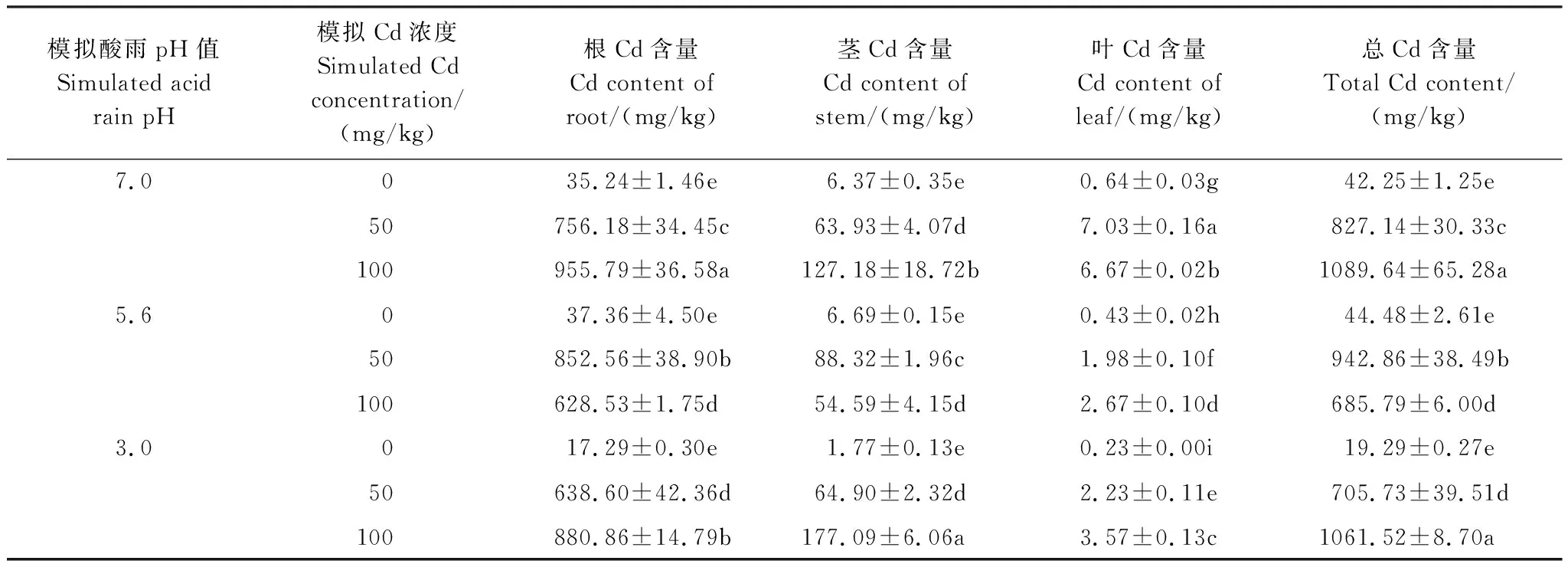
表2 酸雨和镉复合处理下秃瓣杜英镉的积累
2.3 荧光参数
从图2可看出,单独酸雨处理下各荧光参数与对照组无显著差异(P>0.05),可见单独酸雨处理对秃瓣杜英PSⅡ反应中心并没有显著(P>0.05)影响。随着酸雨强度的增加,不同处理间的各荧光参数也无显著差异(P>0.05)。
单独镉处理的各荧光参数与对照均没有显著差异(P>0.05),表明单独镉处理对秃瓣杜英PSⅡ反应中心没有显著(P>0.05)影响。随着镉浓度的增加,两种处理间的各荧光参数无显著差异(P>0.05)。
与对照组相比,复合处理未对植株的各荧光参数产生显著性(P>0.05)影响,说明复合处理对秃瓣杜英PSⅡ反应中心没有显著影响(P>0.05)。各复合处理间,pH 5.6+Cd 100的F0显著(P<0.05)低于pH 5.6+Cd 50,而NPQ显著(P<0.05)高于pH 5.6+Cd 50;pH 5.6+Cd 50的qP值显著(P<0.05)高于pH 3.0+Cd 100。
2.4 酸雨强度、镉浓度及交互作用对秃瓣杜英生理参数的影响
从表3可以看出,酸雨强度对秃瓣杜英的根生物量,总生物量,根、茎、叶、总镉含量和NPQ均有显著(P<0.05)影响;镉浓度对根、茎、叶、总镉含量和qP有显著(P<0.05)影响;酸雨强度和镉浓度的交互作用对根、茎、叶、总生物量,根、茎、叶、总镉含量和F0有显著(P<0.05)影响。
3 讨论
本研究发现,经过短期的酸雨和镉复合处理模拟试验,单独的酸处理显著促进了植株总生物量的积累,具体表现在对根和叶片生物量的提高;发现植株总生物量的提高可能是酸雨中的氮素进入土壤后的施肥效应所导致的。我国亚热带地区的土壤普遍缺氮[32-33],而氮沉降可在土壤缺氮的条件下,提高土壤的有效氮水平。
表3酸雨强度、镉浓度及交互作用对秃瓣杜英生理参数的双因素方差分析
Table3Two-wayANOVEfortheeffectsofpHvalueofsimulatedacidrain,cadmiunconcentration,andtheirinteractionsonthephysiologicalparametersofEleocarpusglabripetalus

参数Parameter酸雨强度Acid rain intensityCd浓度Cd concentration酸雨强度×Cd浓度Acid rain intensity × Cd concentrationFPFPFP根生物量Root biomass29.247*0.000*2.239 0.153 22.364*0.000*茎生物量Stem biomass0.723 0.507 0.374 0.696 4.021*0.030*叶生物量Leaf biomass2.275 0.149 1.180 0.343 7.163*0.004*总生物量Total biomass16.824*0.000*3.382 0.072 38.126*0.000*根冠比Root-shoot ratio3.504 0.066 0.580 0.576 1.772 0.205 根生物量占总生物量百分比The percentage of root biomass to tatal bio-mass3.199 0.080 0.325 0.729 1.854 0.189 茎生物量占总生物量百分比The percentage of stem biomass to tatal bio-mass3.308 0.075 0.067 0.935 2.368 0.116 叶生物量占总生物量百分比The percentage of leaf biomass to tatal bio-mass0.706 0.515 0.298 0.748 0.968 0.470 根镉含量Cd content of root 18.107*0.000*2047.686*0.000*59.244*0.000*茎镉含量Cd content of stem31.450*0.000*428.122*0.000*64.469*0.000*叶镉含量Cd content of leaf2116.708*0.000*3406.707*0.000*496.502*0.000*总镉含量Total Cd content18.630*0.000*1814.122*0.000*72.734*0.000*Fv/Fm1.525 0.244 2.331 0.126 1.349 0.313 F01.768 0.199 0.696 0.511 8.512*0.000*NPQ6.065*0.010*1.350 0.284 1.201 0.345 qP3.243 0.063 7.450*0.004*1.861 0.161
*为显著度为P<0.05
此外,氮素的输入所引起的生物量增加在植物体内分布是不均匀的。因为氮素的输入主要引起叶片中氮含量的上升或增加了植物叶片面积及数目,促进了叶片生物量积累[34-35],与本试验研究结果一致。而相关研究发现在酸雨浓度较低的情况下,低浓度的酸雨对植物的株高、胸径以及叶绿素含量、光合速率、生物量等影响较小甚至有一定的促进作用[36-40]。Thomas等[41]发现在自然条件下,酸沉降在个体水平上促进了24种美国常见植物的生物量的积累,与本研究结果相符。
相同或不同物种的不同生态型或同一植株的不同部位对某一金属的积累能力不同[42- 43]。进入秃瓣杜英体内的镉主要富集在根部,其次在茎,最后为叶。陆志强等[44]也发现镉在白骨壤幼苗中的含量为根>茎>叶>子叶。超积累植物(Hyperaccumulator)指可以超量累积某些化学元素的野生植物。理想的重金属超积累植物一般具有以下特征[45]:(1)可以耐受高水平的重金属;(2)地上部超量积累某种或几种重金属时,不影响植物的正常生长,通常超出普通植物的100倍以上,比如超积累植物积累的镉含量可达100 μg/g (干重)以上。(3)生长迅速;(4)生物量大;(5)根系发达。从秃瓣杜英生长的情况来看,Cd 50显著促进了总生物量的积累,Cd 100虽显著降低了植株总生物量的积累,但生物量分配并未受太大影响。秃瓣杜英在长期的进化过程中对胁迫环境有了较强的适应性,镉浓度较低对植株体无直接伤害作用,反而被作为“养分”而吸收。我们可以看出,秃瓣杜英的镉耐受浓度较高,因此Cd 50促进了总生物量积累,Cd 100虽抑制了总生物量积累,但植株通过稳定生物量分配进行自我保护。复合处理均显著增加了植株根、茎、叶及总的镉含量的积累,但植株的生物量积累与对照组并无显著差异。植株通过根部吸收水分和营养,虽然地上部分也能吸收一部分营养,但根部吸收能力大于地上部分,因此植株可能是通过降低根冠比,增强根部吸收能力来自我调节。而pH 5.6+Cd 100显著促进了总生物量的积累,其总的镉含量显著低于Cd 50,表明其在植株承受范围内。张帆等[46]发现在高浓度的镉处理下,有效氮供应充足,能显著促进一些植株生长及叶绿素含量的增加,与本研究相符。
此外,所有处理的Fv/Fm、F0、NPQ以及qP与对照组无显著差异(P>0.05),表明秃瓣杜英PSⅡ反应中心未受到显著影响。pH 5.6+Cd 100的Fv/Fm值达到最高,即PSⅡ原初光能转化效率达到最高,且NPQ值高于对照,表明该处理对PSⅡ反应中心的影响仍处于植物自身调节范围内,可以通过热耗散的形式进行调节,这可能也是其生物量积累达到最高的原因。pH 5.6+Cd 100的F0显著低于pH 5.6+Cd 50,而NPQ显著高于pH 5.6+Cd 50,说明pH 5.6+Cd 100通过热耗散的手段减少PSⅡ反应中心所受的影响,从而使其生物量积累显著高于pH 5.6+Cd 50。pH 5.6+Cd 50的qP值显著高于pH 3.0+Cd 100,表明pH 5.6+Cd 50的电子传递活性显著高于pH 3.0+Cd 100,因此其生物量积累显著高于pH 3.0+Cd 100。Cd 50的Fv/Fm值和qP值高于对照,说明植株的PSⅡ原初电子受体的氧化还原状态较高、PSⅡ开放中心的数目较多[22],PSⅡ具有高的电子传递活性和原初光能转化效率,因此其生物量的积累显著高于对照。Fv/Fm值和qP值的降低可能是造成Cd 100生物量积累显著下降的原因,但植株通过提高NPQ值,把热耗散作为自我保护的一种手段,稳定了生物量的分配,减缓Cd 100的伤害作用。
所有处理的荧光参数与对照组相比并无显著差异,是什么造成了植株生长的差异呢?推测可能土壤酸化增大了植株根系细胞膜透性,促进镉向细胞的吸附及跨膜运输,增大胞内镉含量,进而加剧对根系的伤害。此外,镉处理亦能增加植株根系细胞膜透性,抑制H+-ATP酶活性,增大细胞外H+输入,降低胞内H+外排,导致胞质更加酸化,加剧对根系的伤害[47]。这导致了植株根系有丝分裂受到抑制,阻碍伸长生长,降低生物量和活力[48]。本实验中,所有处理均使植株根、茎、叶中的镉含量占植株总镉含量的比例发生变化,均提高了根中镉含量所占的比例、降低了叶片中镉含量所占的比例,其中Cd 50及pH 5.6+Cd 100显著降低了茎中镉含量所占的比例。相关研究发现镉能通过影响植物水分及营养元素的吸收,降低作物光合色素的含量和光合速率,抑制光合产物的输送,降低气孔导度,影响叶绿素酶活性,进而抑制光合作用,最终导致植株矮小,生物量下降[49-50]。且经双因素方差分析表明,酸雨强度和镉浓度的交互作用对根、茎、叶、总生物量,根、茎、叶、总镉含量和F0有显著(P<0.05)影响。说明两者之间存在一个“最优组合”,本试验发现pH 5.6+Cd 100处理的镉含量积累是复合处理中最低的,且其生物量积累达到最高。以上可能是导致植株生长产生差异的原因。
本实验初步探究了酸雨和镉及其复合环境处理条件下秃瓣杜英的生长、生理的响应特征,但由于实验时间较短,还不能充分量化不同处理的作用效果,未能明确处理之间的相互关系。且相关试验表明,在相同pH条件下,自然酸雨比模拟酸雨对植物的伤害更大[51]。所以需经过进一步长期观测与测量,才能获得其复合处理环境条件下的响应特征。
4 结论
本研究结果显示:经过短期的酸雨和镉复合处理的模拟受控试验,单独的酸雨处理显著增加了植株总生物量的积累,促进了根和叶片生物量的分配,降低了茎干生物量的分配,且显著降低了叶片中的镉含量。随着酸雨强度的增加,植株器官镉含量积累下降,且生物量积累增加。单独镉处理下,Cd 50显著增加了植株生物量的积累,促进了根和叶片生物量的分配,Cd 100虽抑制了植株生物量的积累,但生物量分配几乎不受影响。复合处理促进了植株根和叶片生物量的分配,降低茎干生物量分配,且降低了根冠比,其中pH 5.6+Cd 100显著促进了植株生物量的积累,其植株镉含量的积累为复合处理中最低。所有处理均未对植株的PSⅡ反应中心产生显著(P>0.05)影响,镉处理和复合处理显著增加了植株镉含量的积累,镉在植株体内的积累为:根>茎>叶。同时,双因素方差分析表明,酸雨强度和镉浓度的交互作用对根、茎、叶、总生物量,根、茎、叶、总镉含量和F0有显著(P<0.05)影响。
