坡位对乳白香青种群特征及雌雄株形态可塑性的影响
2018-09-19刘航江宗人旭刘金平游明鸿
刘航江,宗人旭,刘金平,游明鸿,苟 蓉
(1.西华师范大学西南野生动植物资源保护省部共建教育部重点实验室,四川 南充 637009;2.四川省草原科学研究院,四川 成都 611731)
雌雄异株植物是陆地生态系统的重要组成部分之一,对物种多样性的保护和维持生态系统的稳定性有着积极作用[1]。由于性别的分株表达,雌雄异株植物维持种群稳定性的能力较薄弱,光合作用能力、生物量分配、水分利用效率及开花授粉等方面均易受环境胁迫的影响[2]。面对胁迫时,雌雄个体的生长、形态、生理、生殖及抗逆性等方面存在明显的性别差异[3-5],常引起种群内性比失衡和繁殖能力下降[6-7],导致种群组成、结构和功能发生改变[8]。雌雄异株草本植物具有植株低矮、生活期及寿命短的特征,种群特征和个体发育更易受生境的影响,要求雌雄个体及时、同步、协调、匹配地采取适应生境条件的应对措施,依胁迫因子和胁迫强度调整构件性状和生物量结构,使物质和能量在生长、生殖和抗性等功能间合理分配[9],保证生活史的顺利完成,且产生充足、健康的种实后代,才能在残酷的自然选择中保证物种的存活与延续,避免物种的濒危或灭绝。
乳白香青(Anaphalislactea)为菊科香青属的多年生雌雄异株草本植物,又名大矛香艾和大白矛香,生于海拔2 000~3 400 m的亚高山草地及灌木丛中,是重要的药用植物,性味辛、苦、寒,具活血散瘀、平肝潜阳、祛痰及外用止血功效。因全株密被灰白色绒毛,苞片多层的头状花序常聚成复伞房状,具有特殊的观赏价值[10]。目前仅对其内生溶磷细菌[11]、绿原酸[12]及黄酮成分[13]有相关研究,对其种群特征、雌雄异株特性及雌雄个体形态差异研究较少,尤其对于不同生境下种群特征的变化规律及雌雄个体表型可塑性差异未见报道。
在全球及区域尺度上,气候带是决定植被分布格局的主要因素[14],在景观及更小尺度上,地形等非地带性因素是植被分布格局的主导因素[15]。坡向和坡位是植物群落重要的立地因子,坡向主要影响光照,坡位主要影响土壤养分和水分等的再分配[16-17]。种群是物种存在、物种进化、种间关系的基本单位,种群大小取决于物种的生物学特性和繁殖能力,更依赖于其必需的自然资源和生存空间所允许的限度[18]。本研究以青藏高原东缘亚高山草地中同一山丘不同坡位的雌雄异株草本植物乳白香青为研究对象,通过观测生境特点、种群特征、个体构件性状和生物量累积及分配,分析不同坡位下乳白香青种群特征及雌雄个体表型的差异,研究坡位对种群数、种群密度、年龄结构及性比的影响,探讨雌、雄株个体应对或适应不同坡位的生长及生殖策略,以期为乳白香青资源保护利用及生产扩繁提供依据,为雌雄异株草本植物研究提供借鉴。
1 材料与方法
1.1 试验材料
于2016年7月中旬,以自然分布于四川省阿坝藏族羌族自治州红原县月亮湾4A级景区观景平台所处山丘,处于开花期的野生乳白香青种群为材料。在东北坡向的40°坡的不同坡位,沿水平方向设置20 m×2 m采样带,样带情况如表1所列。
1.2 试验地概况
试验地为大陆性高原温带季风气候,年均温1.1 ℃,极端高温23.5 ℃,极端低温-33.8 ℃,年降水量738 mm,相对湿度71%,≥0 ℃年积温865 ℃·d,年辐射量2.524×107kJ·m-2,土壤为草甸土。
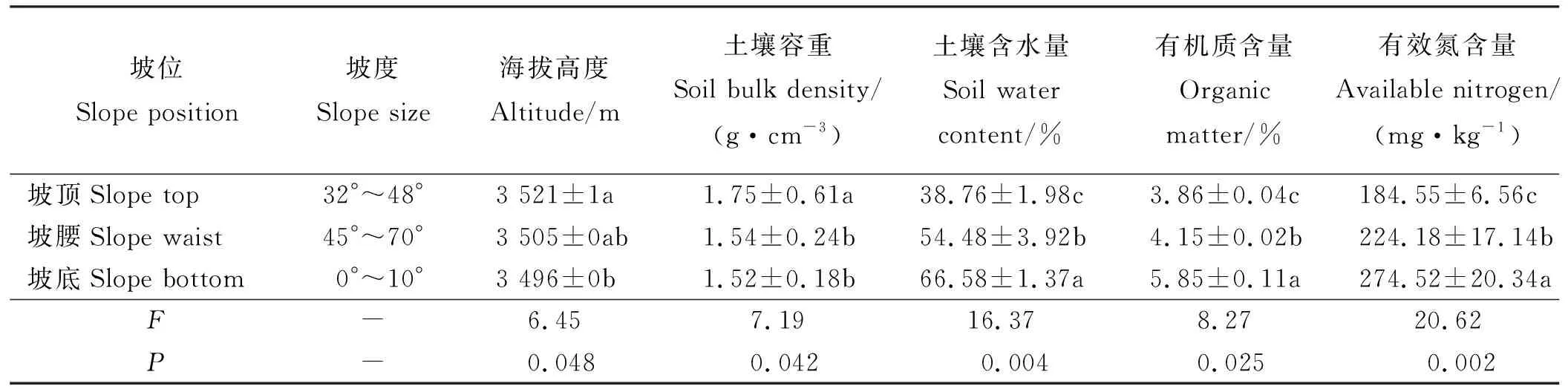
表1 样地基本情况Table 1 Basic characteristics of the three sample habitats
同列不同小写字母表示坡位间差异显著(P<0.05)。下表同。
Different lowercase letters within the same column indicate significant differences between different slope positions at the 0.05 level; similarly for the following tables.
1.3 测量项目及方法
种群数量:随机沿山丘等高线在坡顶、坡腰、坡底用样绳划定20 m×2 m的样地,以株距大于2 m 为判定不同种群依据,测定样方内的种群数量。3次重复。
种群空间格局:在样地内随机30次抛置0.5 m×0.5 m样方,测定样方内株数。
Cx=S2/M。
式中:Cx为分布系数,S2为方差,M为平均值。Cx=0属均匀分布,Cx<1属规则分布,Cx=1属随机分布,Cx>1属集群分布。
种群面积:随机选取种群,沿4个方向测定植物分布的宽度,以均值为直径,拟合圆形,计算种群面积,3次重复。
种群密度:在样地内随机选取3个种群,测定种群面积与植株数,计算单位面积内的株数为密度,3次重复。
种群年龄结构:随机选取3个种群测定种群内幼年(生殖前期)、成年(生殖时期)、老年(生殖后期)的个体数,依据个体数结构判断增长型、稳定型或衰退型。
性比:随机选取3个种群,依据花序性状,测定雌(因雌花序中央偶有雄花,但雄花败育,故两性花植株归为雌株)、雄株个体数,雌株数与雄株数的比值为性比。
株高:随机测定样方内雌雄各20株乳白香青的自然高度。
叶、茎、根性状:随机选雌雄各20株,用直尺法测量其中部成熟茎生叶的叶长、叶宽,用叶厚计测量其叶厚,用计数法测出其叶数;用直尺法测量其茎长,用游标卡尺测其茎径;挖取、清洗、拭干后,测定根的80%长度为根长,用游标卡尺测量其根径。
花性状:随机选雌雄各20株,测花序数、花序宽、花序高,小花数、小花宽、小花高,苞片层数、苞片长、苞片宽。
生物量:随机挖取雌雄各20株完整植株,清洗、拭干后,将根、茎、叶、花器分离,分别装袋,105 ℃下杀青0.5 h,75 ℃下烘至恒重,称干重为生物量,计算含水量:
含水量=(鲜重-干重)/鲜重×100%;
分配比=某部分生物量/总生物量×100%;
水分配比=某部分含水量/植株总含水量×100%。
1.4 数据处理
用SPSS 19.0进行方差分析和多重比较,分析坡位对乳白香青种群特征、构件性状和生物量等指标的影响。
2 结果与分析
2.1 坡位对乳白香青种群特征的影响
不同坡位海拔与坡度的差异致使坡位间草甸土的理化性状差异显著(P<0.05),坡顶土壤含水量、有机质及有效氮含量显著低于坡底 (表1)。坡位对单位面积种群数有显著影响(P<0.05) (表2),坡腰种群数最少,仅为坡顶和坡底的31.02%和23.68%,坡底种群数是坡顶和坡腰的1.31和4.22倍。乳白香青种群空间格局均为集群分布,坡位对分布系数有显著影响,坡底的分布系数显著高于坡顶和坡腰(P<0.05),坡顶和坡腰的分布系数差异不显著(P>0.05)。坡位越低种群面积越大,坡底的种群面积分别约为坡顶和坡腰的3和2倍。
坡位对乳白香青种群密度有极显著影响(P<0.01)(表2),坡底种群密度是坡顶和坡腰的2.44和4.44倍。坡腰种群密度仅为坡顶和坡底的54.96%和22.50%。乳白香青种群年龄结构均为衰退型,且坡位对衰退程度有极显著影响(P<0.01),坡位越高种群衰退越严重,坡顶、坡腰和坡底种群中,老年个体分别是幼年个体的32.5、6.33和3.4倍。坡位对种群性比有极显著影响(P<0.01),坡底种群雌株数是雄株数的5.02倍,其性比显著高于坡顶和坡腰。坡顶的种群性比近于1,随坡位降低性比显著增加,引起性比严重失衡。
坡位对乳白香青种群特征有显著影响(P<0.05),根据P和F值可判断,坡位对种群特征参数影响大小表现为性比>种群密度>年龄结构>种群数>分布系数>种群面积(表2)。
2.2 坡位对乳白香青构件性状的影响
2.2.1叶性状 乳白香青叶长和叶厚在性别间差异较小(P>0.05),雄株叶宽显著大于雌株,而叶数显著小于雌株(P<0.05)(表3)。坡位对乳白香青叶数、叶长、叶宽、叶厚均有显著影响(P<0.05),随坡位降低叶数减少,而叶长、叶宽和叶厚增加。坡底种群植株叶数显著低于坡顶和坡腰,而叶长、叶宽和叶厚显著高于坡顶和坡腰。坡位对叶性状影响有性别差异,由F值可判断,雌株的叶数、叶长和叶宽受坡位影响大于雄株,而雄株叶厚比雌株更易受影响。
2.2.2茎性状 乳白香青雌株株高显著大于雄株 (P<0.05)(表2),坡位越低株高越大,雌株比雄株更易受坡位影响。雌雄株的茎长和茎径有显著性别差异(P<0.05)(表4),坡位对雌雄株茎长均有影响,随坡位降低茎长增加;坡位对雌株茎径有极显著影响(P<0.01),对雄株茎径影响较小。坡底雌株茎长和茎径是坡顶的1.81和2.33倍,雄株则为1.22和0.85倍。由F值可判断,雌株茎长、茎径和株高更易受坡位影响,受影响大小为茎径>茎长>株高。
2.2.3根性状 乳白香青雄株根长和根径显著大于雌株(P<0.05)(表4),坡位对雌、雄株根长和根径均有影响,坡位越低根长越长而根径越细,雌株根长、雄株根径更易受影响。坡底的雌、雄株根长比坡顶增加了37.38%和26.86%,而根径比坡顶降低了19.24%和38.89%。由F和P值结果显示,雌株根长和雄株根径极显著受坡位影响(P<0.01)。
2.2.4花性状 乳白香青的花序高、花序数和小花高无性别差异(P>0.05)(表5),雌株的花序宽、小花数、小花宽和苞片宽显著大于雄株,而苞片层数和长度显著小于雄株(P<0.05)。坡位对雌株花性状影响大于雄株,雌株花序数、花序宽和小花数及苞片宽显著受坡位影响,而雄株花序数、苞片层数和苞片宽易受影响。
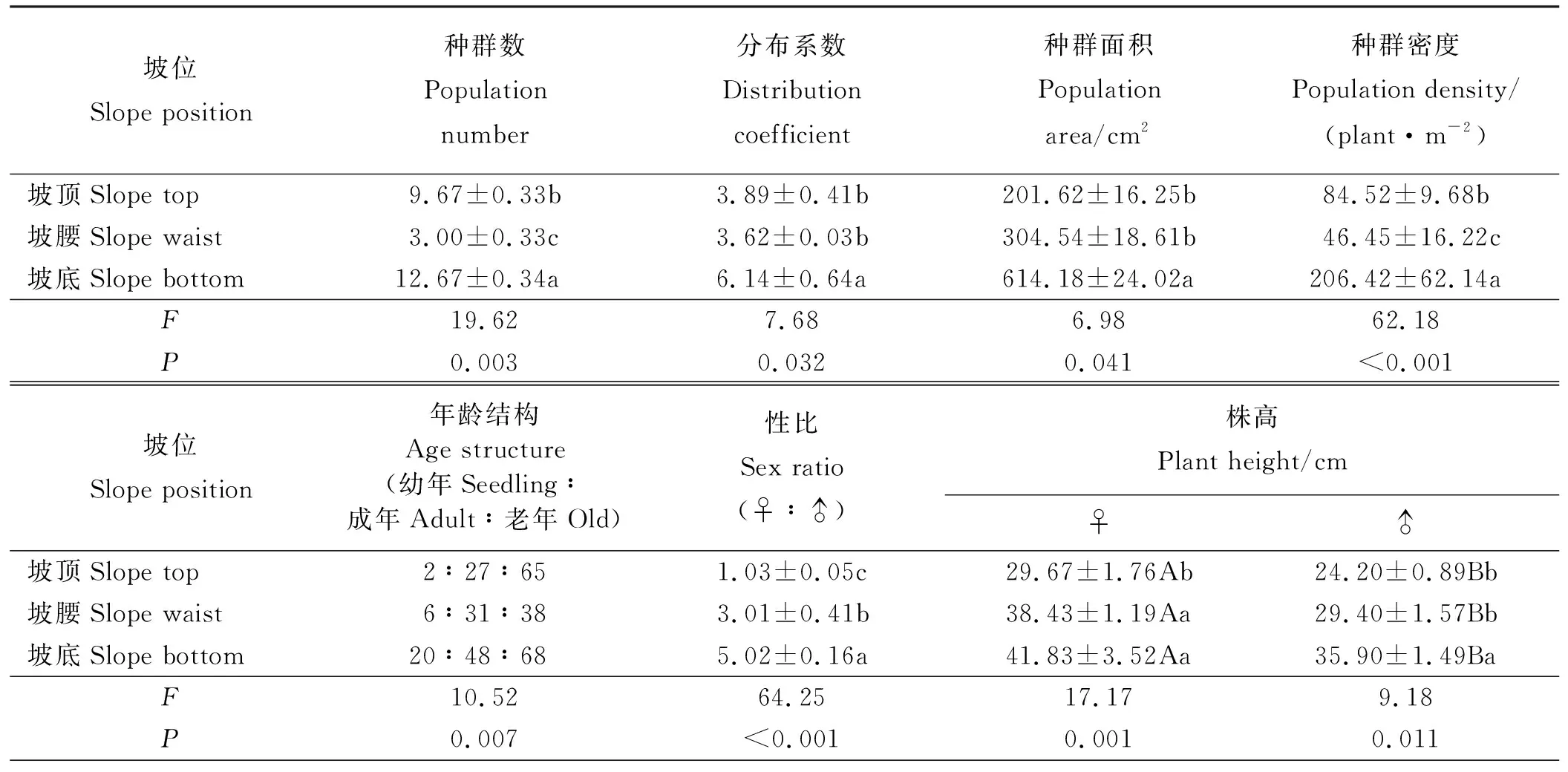
表2 坡位对种群特征的影响Table 2 Effect of slope position on the population characteristics
同行不同大写字母表示性别间差异显著(P<0.05),同列不同小写字母表示坡位间差异显著(P<0.05)。下同。
Different capital letters within the same row for the same parameter indicate significant differences between two genders at the 0.05 level, and different lowercase letters within same column indicate significant differences among different slope positions at the 0.05 level; similarly for the following tables.
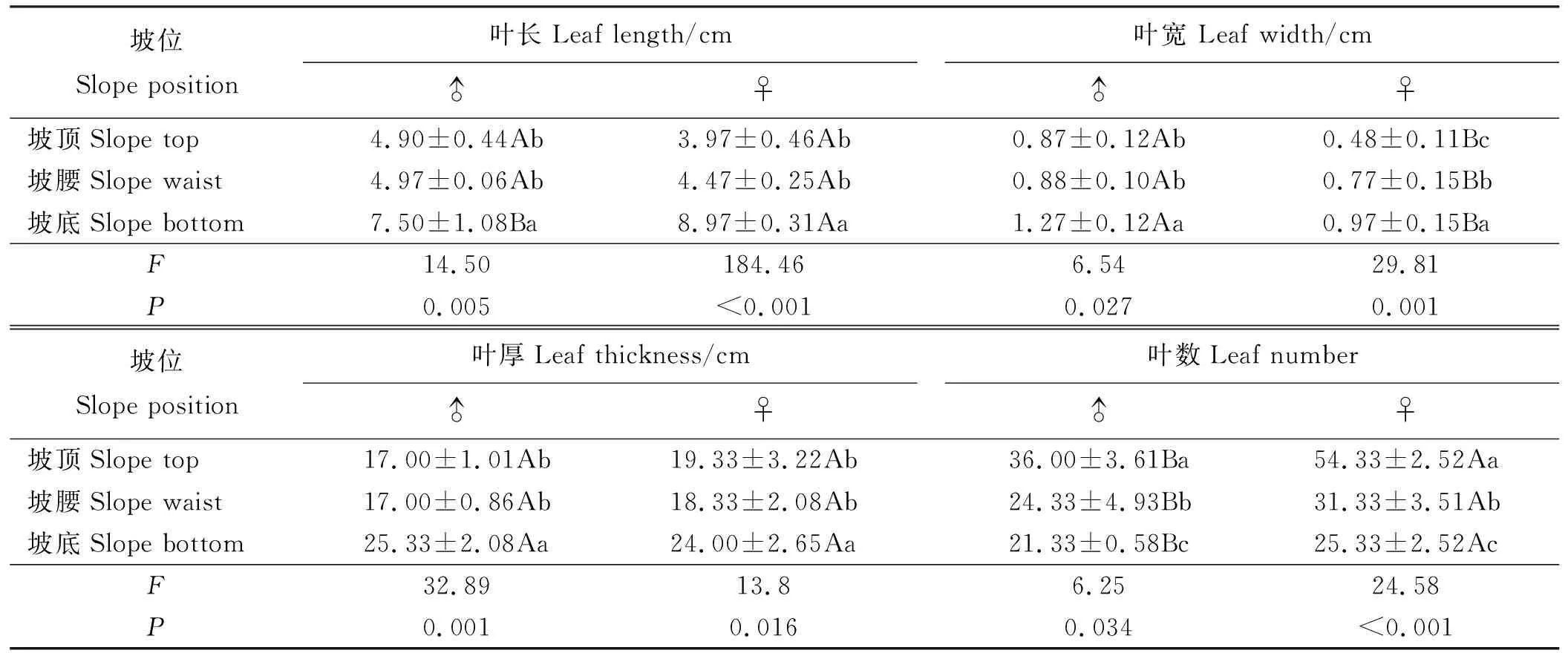
表3 坡位对叶性状的影响Table 3 Effect of slope position on leaf traits
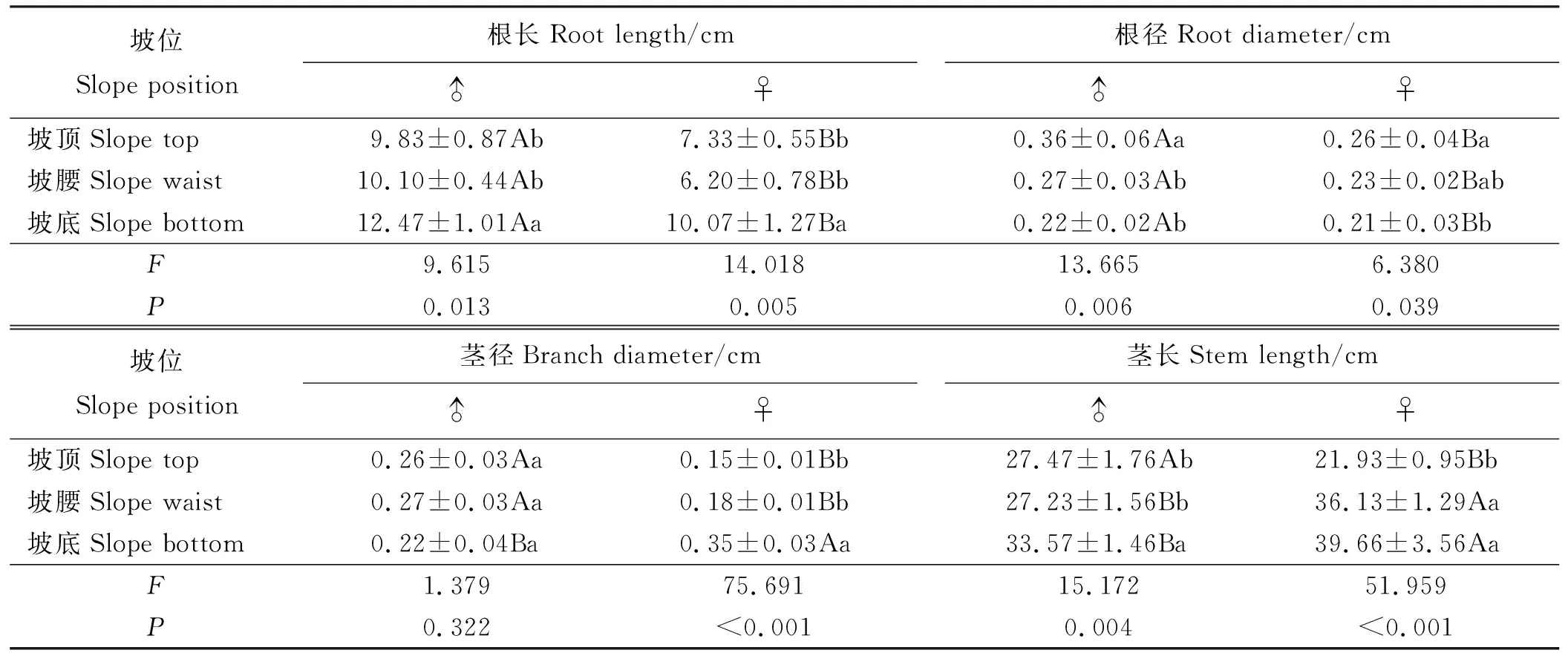
表4 坡位对茎和根性状的影响Table 4 Effect of slope position on stem and root traits
乳白香青的花序高、小花高和小花宽较稳定,受坡位影响较小。
2.3 坡位对乳白香青生物量的影响
2.3.1生物量累积 乳白香青的根和花生物量累积有显著的性别差异,3个坡位上雄株根和花生物量均高于雌株(P<0.05)(表6),而茎、叶生物量和总生物量变化无固定性别差异。坡位对叶、根生物量和总生物量均有显著影响,随坡位降低根生物量降低;雄株叶生物量和总生物量在坡腰下最大,雌株则在坡底最大。坡位对雄株花生物量有显著影响,对雌株花生物量影响较小。坡位对雌株茎生物量有极显著影响(P<0.01),对雄株几乎无影响。坡顶和坡腰的雄株根、茎和花生物量均显著大于雌株(P<0.05),坡底雄株根和花生物量大于雌株,而茎、叶生物量和总生物量显著低于雌株。
2.3.2生物量分配 乳白香青的根、茎和叶分配比的性别差异因坡位而不同,而3个坡位雄株花分配均高于雌株(P<0.05)(表7)。坡位对雌、雄株根分配有显著影响,坡顶和坡腰的根分配显著高于坡底(P<0.05),且雌株在坡顶和坡腰的根分配显著大于雄株,而坡底雌株根分配仅为9.09%,不足雄株的1/2。坡位对雄株茎分配有极显著影响(P<0.01),对叶分配无显著影响;而雌株茎和叶分配受坡位显著影响(P<0.05)。雌株花分配受坡位极显著影响(P<0.01),而雄株花分配相对稳定。
2.3.3坡位对乳白香青水分的影响 乳白香青根含水量在性别间和坡位间均无显著差异(P>0.05)(表8)。雄株茎、叶含水量显著高于雌株(P<0.05),而花序含水量无显著性别差异。坡位对乳白香青茎、叶和花含水量有极显著影响(P<0.01),雌雄株的茎、叶含水量随坡位降低均逐步增加。坡底的茎和叶含水量比坡顶和坡腰增加15%左右。雌株的花含水随坡位降低显著增加,而坡腰的雄株花含水量显著低于坡顶和坡底。
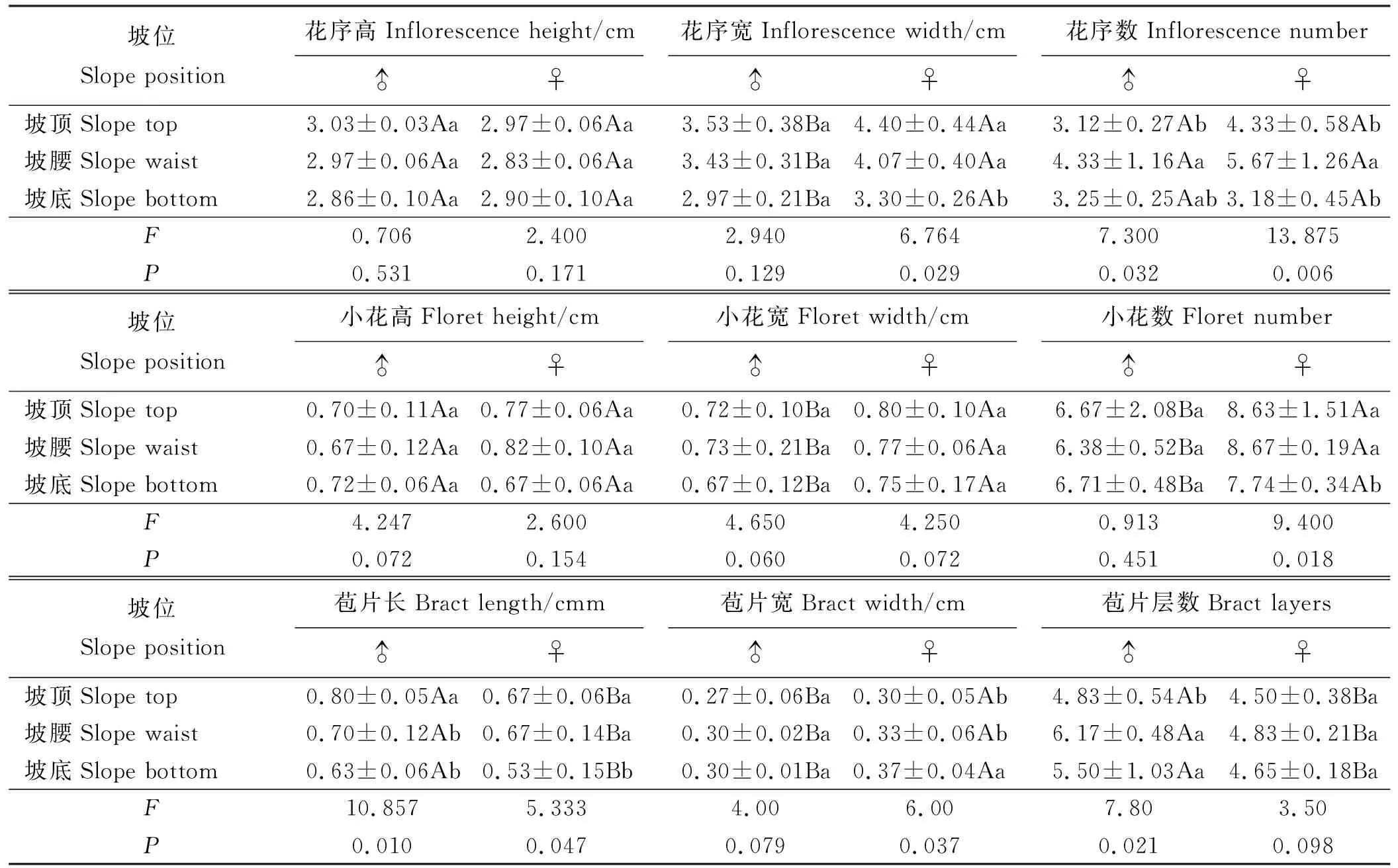
表5 坡位对花性状的影响Table 5 Effect of slope position on flower traits
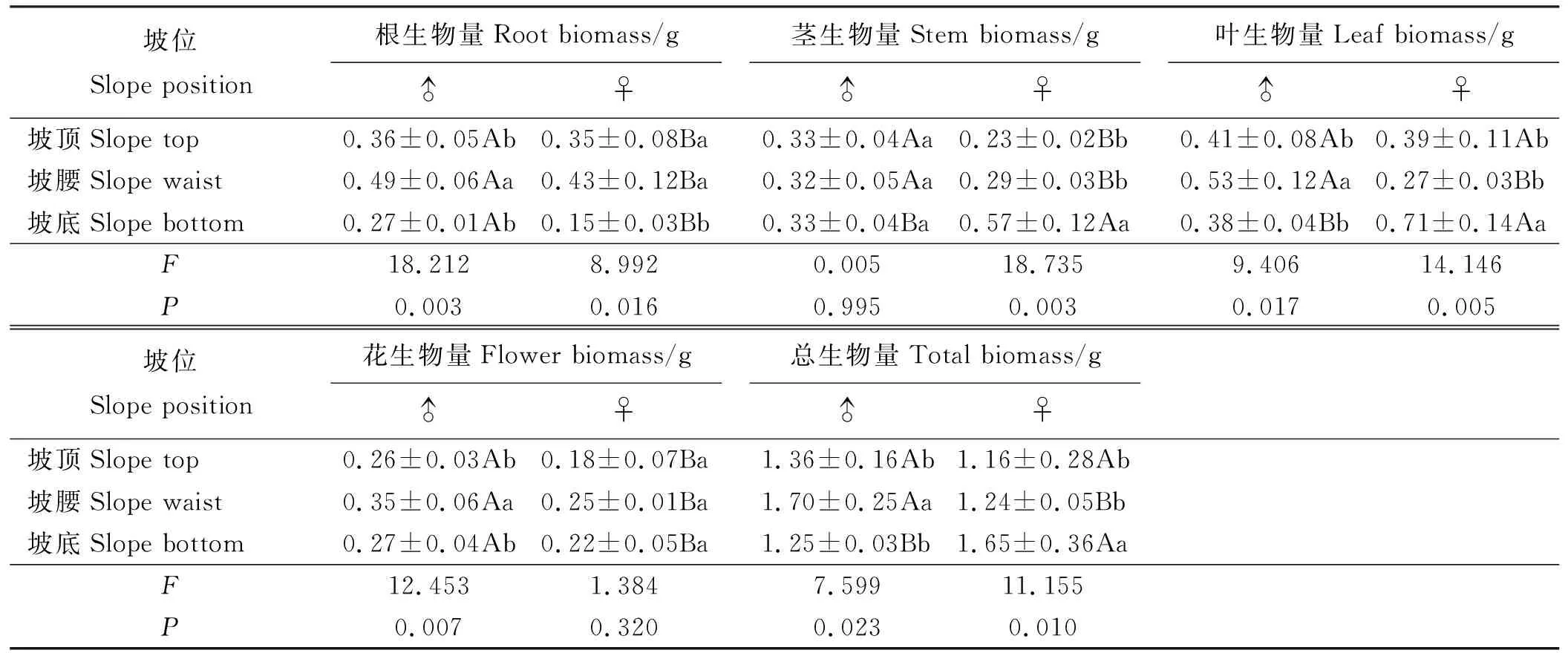
表6 坡位对单株生物量累积的影响Table 6 Effect of slope position on single plant biomass accumulation
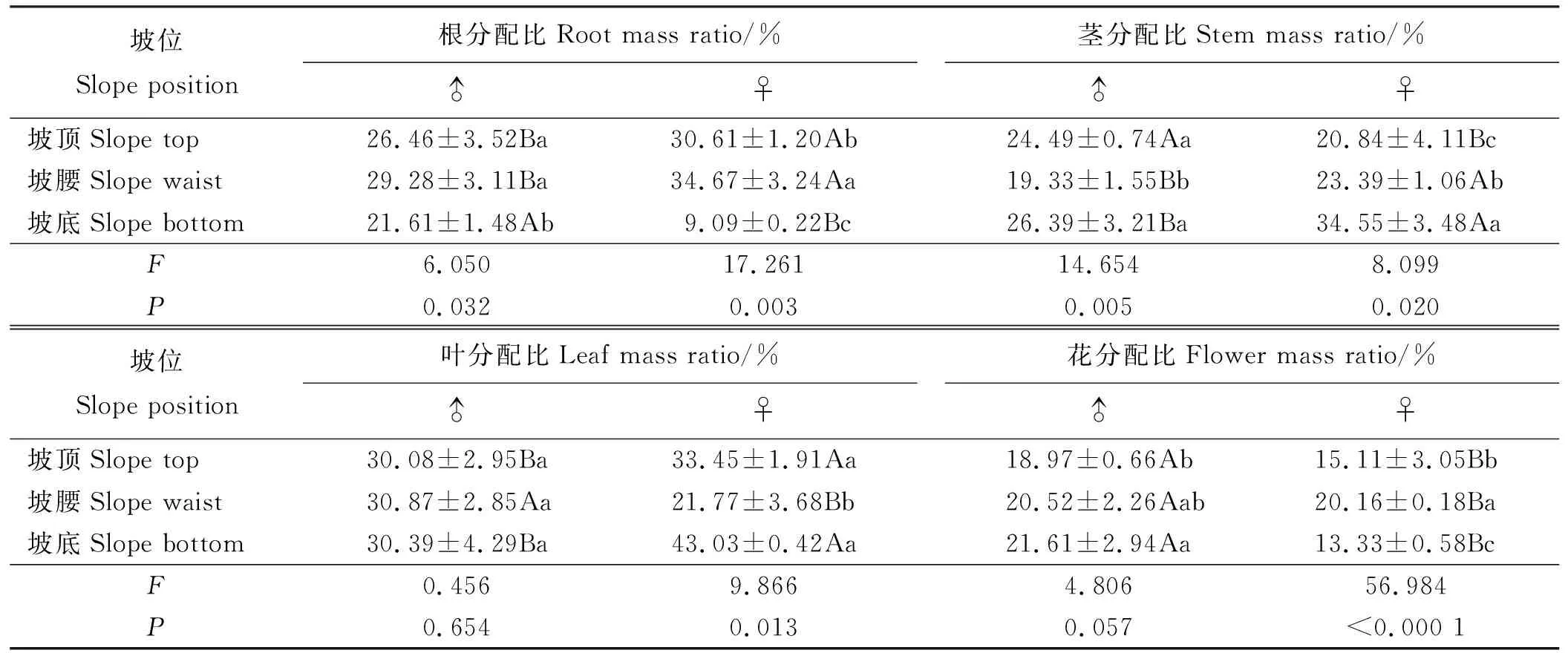
表7 坡位对生物量分配的影响Table 7 Effect of slope position on biomass allocation
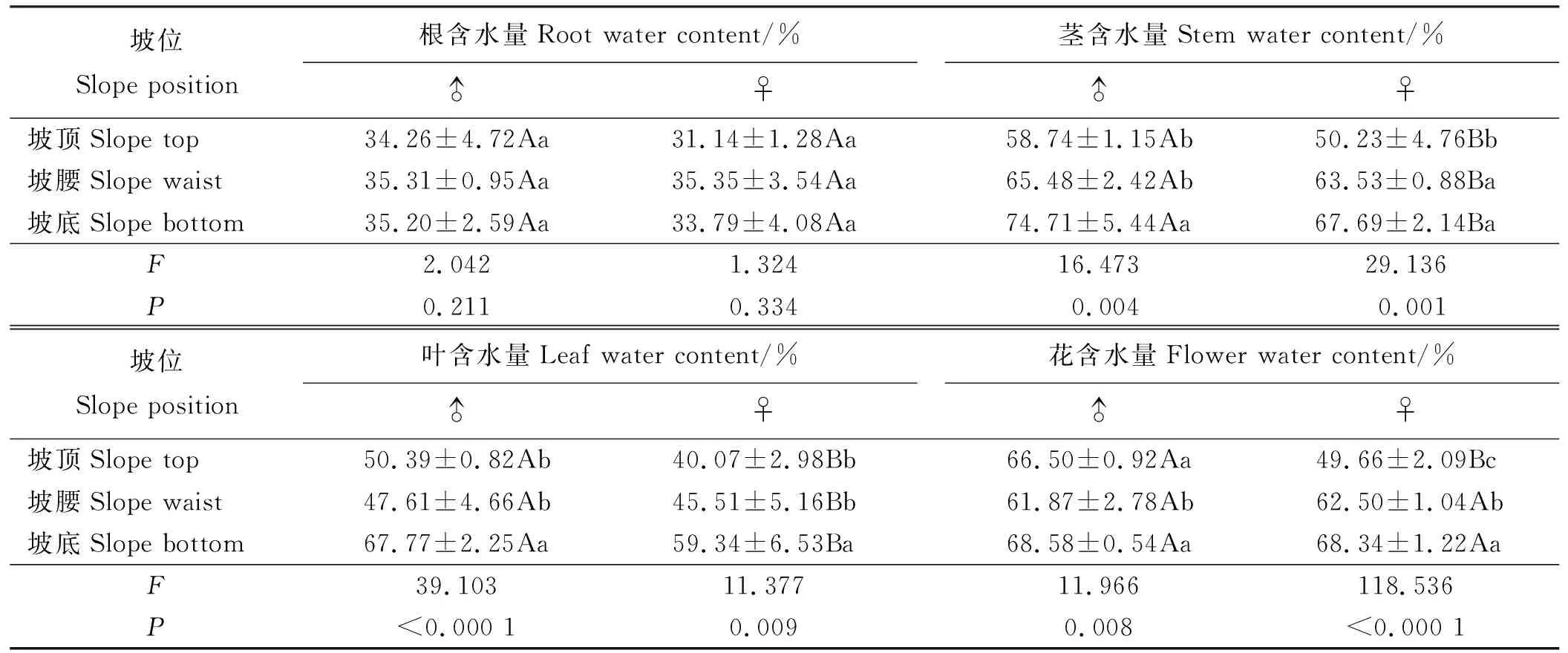
表8 坡位对构件含水量的影响Table 8 Effect of slope position on water content of components
3 讨论
不同坡位因海拔和坡度的差异,地貌过程和起伏变化,风蚀、水蚀强度不同,其表层土壤具有异质性[19-20],表层土壤紧实度、养分和水分显著差异,直接影响物种沿坡面的分布格局,使不同坡位的建群种或优势种差异较大[21],间接通过资源因子的空间再分配影响植被分布,导致不同坡位植被类型明显分异,形成坡顶以灌丛为主,坡腰以菊科(Asteraceae)和玄参科(Scrophulariaceae)为主,坡底以禾本科(Gramineae)和莎草科(Cyperaceae)为主的群落复合体。本研究中,坡底演替为典型的高寒草地群落,测定结果与“上部坡面发育为该区气候顶级群落(climatic climax),下部坡面发育为地形群落”不同,或许受取样尺度与景区人为干扰的影响有关。坡位导致的环境异质性为小尺度范围内生物多样性的形成与维持提供了一种重要机制[22]。本研究仅关注坡位对乳白香青种群特征的影响,把群落组成差异及资源因子(光、热、水、土壤养分)分配当成坡位固有特征,采用了沿等高线样带取样法,一定程度上避免了群落内植物种类、株丛类型及发育状况对乳白香青种群产生的直接或间接影响,关于群落内植物与乳白香青种群形成及个体发育的关系,有待于深入研究。
不同坡位生境限度使乳白香青种群特征表现出显著的差异,坡腰土壤水肥状况优于坡顶,其种群数显著小于坡顶,而单个种群面积随坡位降低而增加。或许坡顶灌丛有利于截留种子,坡腰45°~70°坡面及外在风力影响了种子截留与散布,种子仅在凹陷背风处滞留与萌发,水肥对幼苗的存活和生长促进作用致使种群面积随坡位降低而增大。乳白香青由其花序构造和种实脱落方式决定了种群空间格局为集群分布,高于小花的覆瓦状排列的多层总苞片随种子发育紧裹头状花序,阻挡了瘦果的风播能力,头状花序秋冬季随地上部分干枯才整体脱落,种子在苞片与托盘腐烂后才能散布,致使乳白香青成集群分布。土壤水肥差异对种子萌发及幼苗生长的影响,致使坡位对乳白香青种群密度及分布系数有极显著影响。因草本植物在春季发芽或返青,使花期的乳白香青种群年龄结构为衰退型,坡位越高种群衰退越严重,表明一定范围内水肥条件越差开花结果越早。自然状况下雌雄异株植物所产后代性比为1,乳白香青种群性比随坡位降低而显著增大,良好生境下减少雄株数量可降低种内竞争,提高存留雄株的传粉效率,增加种群产生更多种子的几率。关于乳白香青种群密度调控及自疏机理、引起雌雄个体竞争的生境因子、造成雄株竞争力低于雌株的原因及同性植株间与异性植株间竞争差异等问题还有待于深入研究。
表型可塑性是植物对环境条件或刺激的重要反应特征,植株通过表型可塑性和异速生长响应环境变化带来的压力[23],调整生存策略以增加种群的竞争能力和对胁迫环境的适应能力[24],植株通过构件间权衡与分配的相互作用,形成了适应异质生境的形态和生理特征[25]。在高寒地区,植物面临复杂且不稳定的多种外界环境因素的干扰[26],雌雄异株植物乳白香青在长期的自然选择中得以物种延续,表明雌、雄株能依据立地条件同步、协调、匹配地调整生长占繁殖的策略。本研究发现,乳白香青构件形态存在一定的固有性别差异,雌株叶数多于雄株、雄株叶宽大于雌株,说明雌雄株在捕获光能及散失水分方面采取了不同的策略。种群中雌株显著高于雄株,且花序宽、苞片宽和小花宽大于雄株,而苞片层数和长度小于雄株,使雌花花型更大、开放度更高,更易吸引传粉昆虫和外来花粉,一定程度上避免了种群内的自交频度,增加了种群间基因的交流。乳白香青的根、茎和叶等营养器官的性状都对坡位变化展示出表型可塑性,且表现出一定的性别差异,雌株的叶数、叶长和叶宽、根长、株高和茎径,比雄株更易受坡位影响,而雄株叶厚、根径更易受影响,表明雌株更具张力的表型可塑性及对生境的适应能力,而雄株采取较为“保守”的生长策略,致使雄株比雌株更易受生境胁迫的危害,或许是造成乳白香青种群性比失衡的原因之一。乳白香青的花器官对坡位变化也表现出一定的可塑性, 雌株花性状可塑性大于雄株,雌株通过调节花序数和小花数等决定配子数量的性状适应生境,雄株通过改变苞片层数和苞片宽等附属结构而不减少提供配子数量的方式应对胁迫,因性别分工差异且雌株比雄株有更高的繁殖成本[22],雌、雄株花性状大相径庭的适应表现,殊途同归地为产生更多的后代采取了最佳的繁殖策略。同时,决定细管状小花大小的小花高和小花宽均较稳定,保证了传花授粉正常进行及种实的发育,为生产健康饱满的种子及种群的延续提供了保障。
生物量是体现植物能量积累的主要指标,生物量在各器官中分配差异可直观反映植物适应环境的生长策略[24],生物量分配常受光照、温度、水分、营养等生境因子的影响[25]。植株构件生物量和分配格局是植物与环境共同作用的结果,反映植物对环境的适应能力及环境对植物的影响程度[27],生物量合理的分配使植物形态上更快的适应环境,且满足植物生存、生长、繁殖及抗逆等功能的需要。坡位对水肥资源的再分配,使乳白香青生物量累积和配比对坡位变化有明显的响应。坡位对雌、雄株的叶、根生物量和总生物量均有显著影响,而对雄株的茎生物量和雌株的花生物量累积几无影响,表明乳白香青叶和根对生境敏感度较强,尤其随坡位变化根生物量分配显著变化,使植株根系吸收水肥能力随生境条件进行自我调控,从而增加了乳白香青抗旱性、抗涝性和耐贫瘠性,致使乳白香青在沼泽地、沙化地及草甸中均有自然种群的分布。构件生物量分配大小实质是植物获取资源难易度及资源在生理间或功能间权衡的结果,雄株茎生物量、叶分配比和花分配比相对稳定,雌株仅花生物量受生境影响较小,而雌株的根、茎、叶和花分配比显著受坡位影响,表明雌株对胁迫生境具有更高的适应策略。植物繁殖分配与个体大小呈线性负相关关系,繁殖代价可用繁殖器官数量或生物量进行表达,乳白香青繁殖分配与雌雄株个体大小未呈线性关系,雄株的花生物量及花分配比均高于雌株,表明雄株比雌株把更多的物质和能量用于生殖器官,便于提供足量的花粉,完成雄株在种群繁殖中的作用。而雌株依据生境适合度决定生殖投入比例,且用于生殖的物质总量趋于稳定,雌株灵活又控制总量的生殖投入,是开花结实养分消耗与生境可供资源及植株各项功能权衡的结果,使种群依据生境条件产生适量发育完熟的种子。
4 结论
坡位对乳白香青空间分布格局影响较小,对种群数、种群大小、种群密度、分布系数、年龄结构及性比等种群特征参数有显著影响,影响大小表现为性比>种群密度>年龄结构>种群数>分布系数>种群面积。坡底种群数、种群面积和密度大于坡顶和坡腰,且随坡位降低,种群衰退度减慢,而雌株比例不断增加,使不同坡位的种群表现出不同的适应性种群特征,使生长期延长和性比越失衡。坡位对乳白香青构件性状、生物量累积与分配、水分分配有显著影响,其表型性状、资源分配格局和生物量配比具有极强的可塑性。虽雌、雄株在构件性状和生物量分配有一定的性别差异,但雌、雄株均能依据坡位做出同步且匹配性的响应,合理使物质和能量在生存、生长、生殖及抗性等功能间的进行分配,为顺利完成生活史和延续种群奠定了基础。
