人工调控对三江源区多年生禾草混播草地生产性能的影响
2018-09-19施建军马玉寿董全民邵新庆武高林王彦龙张春平王晓丽
施建军, 马玉寿, 董全民, 邵新庆, 武高林, 王彦龙, 刘 玉,张春平, 王晓丽*
(1. 青海畜牧兽医科学院, 青海 西宁 810016; 2. 青海大学, 青海 西宁 810016; 3. 三江源区高寒草地生态省部共建教育部重点实验室, 青海 西宁 810016; 4. 中国农业大学, 北京 100193; 5. 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西 杨凌 712100)
调查表明,三江源区退化草地面积已占草地总面积67%以上[1-2],其中高寒草甸极度退化后形成的黑土滩面积达到了7 363.42万亩[2],占高寒草甸总面积的30.97%。因黑土滩退化草地植物群落中原生植被莎草科植物几乎消失,自然繁殖更新能力极低,加之特殊的高寒气候条件,植被恢复非常艰难漫长且水土流失严重,必须通过人为干预,采用重建和改建的方法恢复其植被[2-4]。研究表明,选择适宜多年生禾本科牧草建立人工草地是恢复黑土滩退化草地的生产、生态功能的快速和有效途径之一[4-7],并得到广泛推广。目前黑土滩退化草地上建立的人工草地,主要以垂穗披碱草(ElymusnutansGriseb)、青海中华羊茅(FestucasinensisKeng ‘Qinghai’)和青海冷地早熟禾(PoacrymophilaKeng‘Qinghai’)为主[7-10],缺乏耐牧的根茎型草种,草皮形成速度较慢[9-11]。同时,黑土滩退化草地优势种主要是阔叶型杂类草,地下根系发达,加之人工草地建植后重利用而轻管理,人工群落逆向演替较快[8-10]。因此,如何维持多年生禾草混播人工草地的生产稳定性,是建植高寒人工草地的关键[12-14]。适宜的品种组合是实现混播群落产量和组分稳定性的基础,合理的人工调控措施是维持混播群落生产力的主要途径[15-18]。我们前期的研究结果表明,选用6种当地野生驯化草种,垂穗披碱草、青海草地早熟禾(PoapratensisiL.‘Qinghai’)、青海中华羊茅、青海冷地早熟禾、碱茅(PuccinelliadistansL.)和西北羊茅(FestucarylcviaanaRererd.)[5-7],按照上繁草和下繁草、丛型与根茎型搭配的原则[7-10,13],可以建植较为稳定的多年生人工草地。因此,本研究以混播组合为基础,采用杂草防除+施肥的人工调控和对照2个处理,探讨不同草种组合和调控措施下人工群落的生产力与稳定性,阐明草地群落生产力稳定性的合理调控措施,为黑土滩退化草地的治理提供技术支撑。
1 材料与方法
1.1 试验地概况
试验地位于果洛州玛沁县大武镇大武一大队的冬春草场上,地理位置为34°27.821′ N,100°12.367′ E,海拔3 760 m。年均温-3.9℃,≥5℃积温850.3℃,1月的平均气温为-12.6℃,7月的平均气温为9.7℃。牧草生长季为150天左右,无绝对无霜期。年均降水513.2 mm,年蒸发量2 471.6 mm。
试验区建植人工草地前是黑土滩退化草地,土壤为高山草甸土,主要建群植物为伏毛铁棒锤(AconitumflavumHand.-Mazz.)、西伯利亚蓼(PolygonumsibiricumLaxm.)鹅绒委陵菜(PotentillaanserinaL.)、光稃香草(HierochloeglahraTrin.)等。鼠害严重,牧用价值已近丧失。
1.2 试验设计与方法
1.2.1试验设计 试验设置人工调控(杂草防除+施肥)和对照2组处理,每个处理6个混播组合(表1)。所有处理的人工草地均进行灭鼠和围栏管理。
1.2.2试验草种 试验选用的6种牧草均为当地野生驯化草种[4-6]。其中,垂穗披碱草和青海中华羊茅为疏丛型上繁草,青海冷地早熟禾和碱茅为疏丛型下繁草,西北羊茅密丛型下繁草,青海草地早熟禾为根茎-疏丛型下繁草。青海草地早熟禾原种来源于青海黑土滩草业科技有限公司,其余原种来源于青海省牧草良种繁殖场。
1.2.3混播群落组合配置 试验以当地种植面积最广的垂穗披碱草为基础,依次添加其它草种,共组成6个组合,设为A、B、C、D、E、F和Ack、Bck、Cck、Dck、Eck、Fck(表1)。
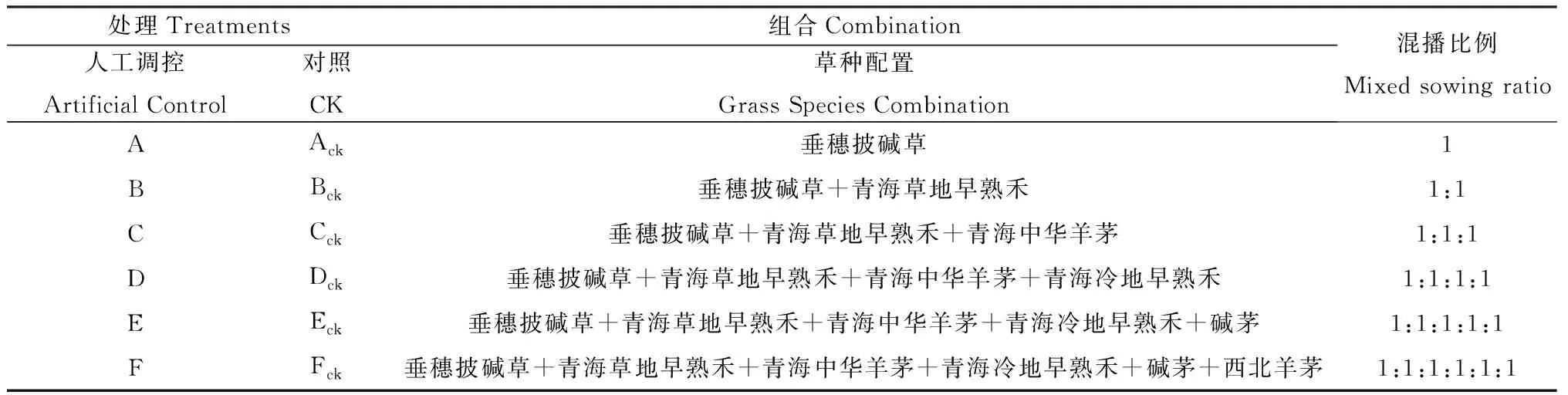
表1 混播组合配置表Table 1 Different Treatments and Mixed Combinations
1.2.4试验小区设计与播种量 小区面积为3 m×4 m,完全随机区组排列,三次重复。播前土壤深翻25 cm,机械耙平,人工开沟条播,覆土深度为2~3 cm。2006年5月8日播种,播前施磷酸二铵15 g·m-2,尿素7.5 g·m-2作底肥。组合中各种牧草的播种量=该种牧草的单播播种量/n,n为各组合中牧草组成种数。垂穗披碱草的单播播种量为3 g·m-2,青海草地早熟禾、青海冷地早熟禾、碱茅和西北羊茅的单播播种量为0.75 g·m-2,青海中华羊茅的单播播种量为2.25 g·m-2。
1.2.5田间管理 建植前试验地及周围200 m以内彻底灭鼠,之后每年3月对试验区进行害鼠防控,达到无鼠害状态。试验区围栏保护。播种后,每年4月~11月严格禁牧,其余时间放牧利用。人工调控试验区于2006年7月上旬人工除杂1次。2007年6月10日用10%甲磺隆可湿性粉剂和72%的2,4-D丁酯乳油配合防除阔叶型杂草,并在6月下旬进行一次人工除杂;于6月5日和7月8日追施尿素两次,施肥量分别为15 g·m-2和7.5 g·m-2。2008年6月中旬药物除杂一次(除草剂和用量同上年),7月初以尿素22.5 g·m-2追肥1次。2012年6月初用10%苯磺隆可湿性粉剂0.075 g·m-2兑水防除阔叶型杂草,7月初以尿素22.5 g·m-2追肥1次。2014年调控同2012年一致。2015年和2016年7月初以尿素22.5 g·m-2分别追肥1次。对照试验区播种后不施肥也不灭杂,仅进行灭鼠和围栏处理。
1.2.6生物量测定 每年8月初至9月上旬测定,每小区内随机设1 m×1 m的样方(边际不取)1个,样方内牧草当年齐地面刈割,第2年起留茬5 cm刈割,测定鲜重,取500 g带回实验室在65℃下烘48 h以上至恒重为干重,折算群落干物质。
2 结果与分析
2.1 人工调控对草地群落生物量的影响
2.1.1人工调控对群落总生物量的影响 施肥和杂草防除干扰了群落的生物量构成,杂草防除降低了群落生物量,施肥提高群落生物量,与对照之间存在显著差异。年际间比较,建植当年,人工调控处理下各混播组合生物量均显著低于相应对照(P<0.05),主要是人工调控处理通过杂草防除,杂类草被铲除,导致生物量降低。生长第二年,受植物特性和施肥的影响,人工调控下各混播群落的生物量最高,极显著高于对照,是对照的2倍左右。通过施肥和杂草防除,人工调控下各组合群落生物量维持在一个较高的生产稳定状态,显著高于相应对照(P<0.05),对照的生物量总体呈现降低的趋势。自生长第2年至第11年,人工调控下组合A、B、C、D、E、F群落总生物量年际间分别比相应对照Ack、Bck、Cck、Dck、Eck、Fck提高34.41%~113.21%、55.43%~122.72%、56.36%~188.39%、97.88%~131.74%、93.78%~139.21%、85.78%~141.27%,差异显著(P<0.05)(图1)。人工调控对群落生物量的影响与群落组成关系十分密切,不同混播组合对人工调控措施的影响具有差异性,随组分种数的增加,群落生物量在人工调控下更趋于稳定,表明人工调控措施对禾草混播群落维持生产稳定状态作用显著,合理的人工调控措施可使高寒人工草地保持在高生产力的生产稳定状态下,取得较高的生产经济效益(图1)。从年际间变化来看,人工调控中施肥的效应要大于杂草防除,2014—2016连续3年施肥,群落生物量始终维持在高水平上,而对照却呈现平稳的降低趋势。而在2010—2011连续2年未施肥的情况下,2012年虽然进行了杂草防除和施肥管理,但群落生物量明显下降,尤其是2~3组分的群落,生物量下降显著(P<0.05)(图1)。

图1 不同混播组合群落生物量的年际变化Fig.1 Yearly variation of aboveground total biomass of community in grass mixtures
2.1.2人工调控对群落禾草生物量的影响 禾草是群落中的主要功能群,是群落生物量的主要贡献者。人工调控下禾草生物量占总生物量的90%以上,直接决定着群落生物量的变化,其年度变化与群落生物量变化一致。对照的禾草生物量占群落生物量的比例在50%以上.随着生长年限延长,占群落生物量的比例逐步降低。建植当年,人工调控下群落禾草生物量略高于对照,但差异不显著。第2年,人工调控下禾草生物量达到生长高峰,极显著(P<0.01)的高于对照,分别比对照提高79.80%~140.28%。生长3年后,人工调控对群落禾草生长促进作用逐步增强,与对照之间的差异逐步加大,差异极显著(P<0.01),生长3、7、9、10、11年,人工调控下各混播组合禾草生物量分别较对照提高151.69%~174.174.76%、90.12%~167.03%、152.54%~193.22%、158.41%~196.41%、218.21%~379.74%。人工调控下组合A、B、C、D、E、F与对照Ack、Bck、Cck、Dck、Eck、Fck比较,不同年限呈禾草生物量分别提高90.12%~218.21%、79.80%~293.74%、87.18%~379.74%、140.28%~258.61%、119.52%~222.9%、127.36%~200.46%,增产效果显著,提高比例随生长年限呈线性增加(图2)。因此,持续的人工调控措施可促进群落中禾草的生长生殖,抑制杂类草的生长生殖,使混播群落能维持在高生产性能的稳定状态。同时表明,在黑土滩退化草地上建植多年生禾草人工草地,因其土壤养分贫瘠,要维持人工草地的高产性能,持续的养分输入和杂类草防控是十分必要。
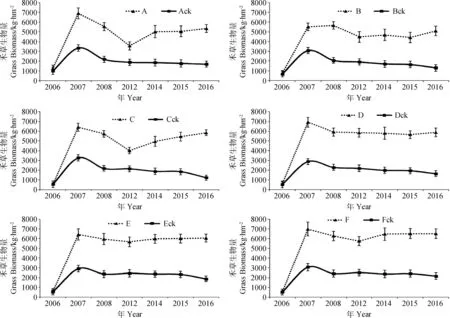
图2 不同混播组合群落禾草生物量的年际变化Fig.2 Yearly variation of aboveground grass biomass of community in grass mixtures
2.1.3人工调控对群落杂类草生物量的影响 建植当年,通过人工除杂措施,调控组合中90%以上的杂类草被清除,组合Ack、Bck、Cck、Dck、Eck、Fck与A、B、C、D、E、F比较,对照组合杂类草生物量是相应调控组合的11.6~32.8倍。第2年,通过人工与药剂防除,杂类草进一步得到控制,对照组合杂类草生物量是相应调控组合的1.7~12.0倍。自第3年起,群落趋于稳定,杂类草以多年生杂类草为主,通过药物防除和施肥干扰后,调控组合群落中的杂类草相比于对照只是降低了高度和抑制了生殖生长,对照组合杂类草生物量比相应调控组合增加25.12%~202.31%(图3)。
2.2 不同混播组合群落生物量对人工调控的响应
2.2.1不同混播组合群落总生物量对人工调控的响应 人工调控下,建植当年人工调控下组合A群落总生物量显著高于其他组合(P<0.05),其余组合之间差异不显著,大小顺序为A>B>D>C>E>F(图4)。对照组合与人工调控基本一致,大小顺序为Ack>Bck>Dck>Eck>Cck>Fck(图4)。生长第2年,人工调控下组合F群落生物量最高,显著(P<0.05)高于其它组合,排序序为F>D>A>C>E>B(图4)。而对照组合Ack群落总生物量显著高于其他组合(P<0.05),排列为Ack>Cck>Bck>Fck>Dck>Eck(图4),与人工调控不同,少组分群落高于多组分群落。自生长第3年开始,人工调控下群落总生物量最高的是组合F,最低的是A或B,两者之间差异极显著(P<0.01)。而对照在第3年群落生物量最高的是垂穗披碱草单播Ack,且与其它组合之间差异不显著(图4)。对照群落在7~11年生物量最高的组合是Fck,最低的是Bck或Cck,两者之间差异显著(P<0.05),但与其它组合之间差异不显著。人工调控提高了混播群落的总生物量,混播组分分化合理,生长性能得到发挥。
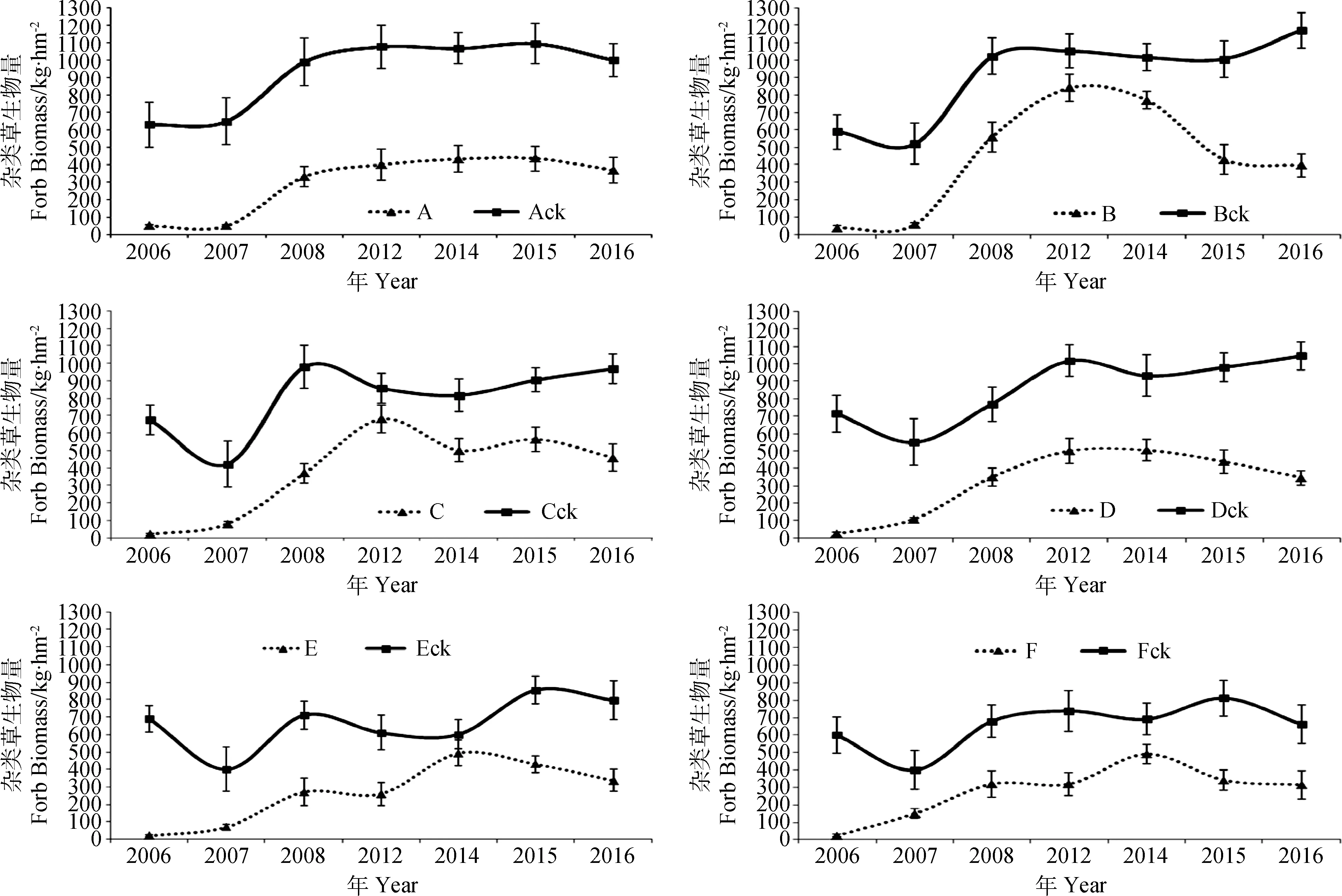
图3 不同混播组合群落杂类草生物量的年际变化Fig.3 Yearly variation of aboveground forb biomass of community in grass mixtures
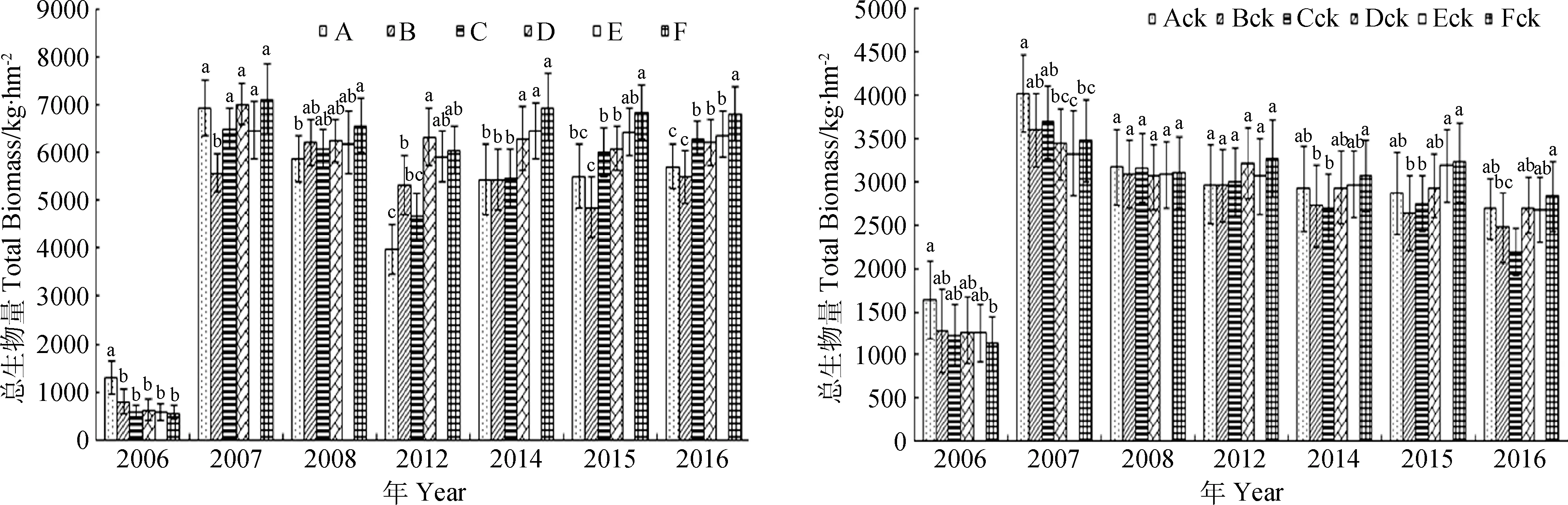
图4 不同处理下群落生物量的年际变化Fig.4 Yearly variation of aboveground total biomass of community in grass mixtures under weedremoval+fertilization and CK
2.2.2不同混播组合群落禾草生物量对人工调控的响应 人工调控下组合不同组合群落禾草生物量变化趋势与总生物量一致,只是随着生长年限增加,多组分群落禾草生物量与少组分群落之间差异显著性增加,变化趋势与混播组分呈正相关(图5)。对照各组合群落禾草生物量与总生物量变化趋势也一致,随生长年限增加,4组分以上群落禾草生物量显著高于其他组合(P<0.05),3组分以下差异不大,但与人工调控的相应组合有差异(图5)。
结果表明,随着生长年限增加,人工调控提高了群落生物量,而混播组合也逐步体现出高矮禾草搭配的优势,5组分以上群落显现出较好的生产性能。
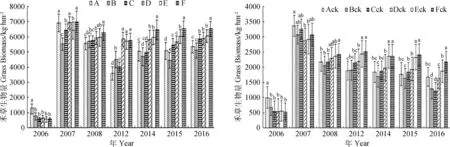
图5 不同处理下群落禾草生物量的年际变化Fig.5 Yearly variation of aboveground grass biomass of community in grass mixtures underweed removal+fertilization and CK
2.3 人工调控对群落组成的影响
人工调控下,组合A中垂穗披碱草比例均在80%以上,对照组合A中垂穗披碱草比例人工较调控低25%以上,通过杂草防除和施肥干扰,能有效提高垂穗披碱草生长生殖,使之处于绝对优势地位。人工调控下组合B中垂穗披碱草比例较对照提高3.58%~40.39%,青海草地早熟禾比较对照组中增加,随生长年限延长呈线性增长。组合C、D、E、F与Cck、Dck、Eck、Fck比较,混播组分种在群落中的比例分别提高10.18%~51.71%、6.31%~52.75%、10.35%~51.72%、7.36%~48.64%,人工调控保持了群落组分种的绝对优势地位,比例维持在90%以上,而对照中组分种的比例在70%以上。
人工调控下各组分种占群落生物量的比例随组分种增加逐步降低,随生长年限增加逐步降低,而杂类草占群落生物量比例随生长年限增加逐步增加。垂穗披碱草是群落生物量的主要贡献者,在群落中所占比例随各组合组分种增加逐步降低,并随着生长年限增加,其所占比例也逐步下降,群落总生物量的高低已不完全跟受控于垂穗披碱草的播种比例。对照组各混播组合垂穗披碱草占群落生物量比例较相应人工调控下降10%~30%,杂类草占群落生物量比例较人工调控显著增加,总体变化趋势与人工调控一致(图6)。人工调控下群落中的下繁草生物量比例随组分种增加逐步增加,并随着生长年限增加逐步增加,它们对群落生产能力的贡献率逐步增大,草种合理搭配的优势逐步展现。人工调控下西北羊茅在前3年对群落生产力的贡献率一直很低,到第9~11年,其所占群落生物量的比例已达到1/3强,表现出很好的生产潜力,可作为一个重要组分种来考虑。对照群落中,随群落生长年限延长,垂穗披碱草的比例逐步下降,杂类草比例逐步增加。青海草地早熟禾和青海冷地早熟禾比列随群落生长年限逐步增加,第9~11年,已趋于稳定。对照群落中,青海中华羊茅在第2、3年较高,之后逐步下降,尤其是在组分较多的组合中,到第9~11年比例下降更多。对照西北羊茅变化趋势和人工调控一致,随群落生长年限延长而增加,已接近垂穗披碱草比例(图6)。

图6 不同处理下下群落组分比例变化Fig.6 Changes of the proportion of species compositions of plant community under weed removal+fertilization and CK
2.4 人工调控措施对牧草群落茎叶比的影响
茎叶比是衡量牧草品质和生产性能的重要指标之一,牧草叶含量的比值越大,就会提高牧草的粗蛋白含量,并促使粗纤维含量降低,从而提高牧草的营养价值。即茎叶比越小,牧草品质越好。人工调控下各混播组合的茎叶比显著低于对应的对照组合,表明人工调控改善了群落结构,优化了高矮禾草叶片的分布。不同混播组合比较,茎叶比以人工调控措施组合F最低,为0.84,表明叶含量大于茎含量,草群叶量丰富,营养价值较高,牧草的品质也较高(表2)。

表2 不同混播组合茎叶比Table 2 The stem-leave ratio of different mixed combinations
3 讨论
人工草地作为草地经营的高级形式,其建植和长期利用的关键就是维持草地群落的稳定性[15-18]。群落稳定性是衡量人工草地质量的重要标准和草地长期保持生产力的基础,也是合理有效的建植、利用、管理及改良人工草地的基本依据[10,17-18]。环境压力和干扰活动是影响人工草地生产稳定性的重要因子,通过降低干扰活动和调节环境压力,去除影响草地健康生长的动物与人为干扰因素(不合理放牧、鼠害和刈割等),添加养分(施肥)和阻止入侵种侵入(杂类草防除),可使人工草地维持较高的生产性能,达到人工草地可持续利用的目的[19-21]。在黑土滩退化草地上建植人工草地,由于土壤基质和肥力较差,牧草定植和生长需要的养分原有土壤无法全部持续的供给,通过施肥来补充土壤养分是维持草地生产力的关键[9-11]。同时,黑土滩退化草地原有的优势种以根系发达的阔叶型杂类草为主,侵占能力很强,尤其是在肥力较好条件下,其生长发育能力要强于优良牧草,防除杂类草有利于播植的优良牧草生长发育[8-9,22]。试验结果表明,在施肥和杂草防除的调控下,群落总生物量比对照提高56.36%~188.39%,且差异显著性随生长年限延长而增加。随生长年限增加,群落中杂类草比例逐步增加,至第11年,杂类草比例已达到30.86%,表明草地在退化。人工草地退化实质上是前茬植物群落的恢复演替,显著特征就是人工种植的优良牧草种类随生长年限增加在草地群落中渐次消失,初级生产力和草地经济利用价值降低[11-12,19,23]。高寒多年生禾草人工草地建植后,在混播群落种间种内竞争、气候生态因子和放牧压力胁迫逐步退化,种植的禾本科牧草在草地群落中渐次消失,向原来的黑土滩植被演替。而在人工调控措施下,人工草地群落中优良牧草生物量随生长年限延长,比对照提高79.80%~379.74%。
垂穗披碱草是三江源区高寒草地的广布种和优势种,试验中从垂穗披碱草单播到6种牧草混播,随群落组分增加,垂穗披碱草在群落中的比例下降,但群落生物量的主要贡献者仍然是垂穗披碱草,印证了种间竞争的能力取决于种的生态习性、生活型和生态幅等,与前人的研究一致[20-21,24]。
当混播组合中组分种达到4种以上时,群落生物量显著高于单播生物量,且随生长年限增加,差异显著性也增加,呈正相关。许多研究都表明,种间竞争促使两物种的生态位分离,各种群在群落中具有各自的生态位,避免直接的竞争,保证了群落的稳定[15-18]。一个相互起作用的、生态位分化的种群系统,各种群对群落的时间、空间和资源利用方面以及相互作用的可能类型方面,都趋向于互相补充而不是直接竞争[13,23,24-26]。因此,由多个种群组成的群落要比单一种群的群落更能有效地利用环境资源,群落净生产力随物种数增加而增加,种群个体之间存在生态位互补,维持长期较高的生产力和更大的稳定性,即资源互补效应[19,21,25,26]。即物种多样性对群落生产力存在正效应,群落净生产力随物种数增加而增加,种群个体之间存在生态位互补,这就是混播人工草地在群落稳定性和生产力方面优于单播人工草地的原因[27-29],本试验也验证了这一点。随着生长年限增加多组分群落产生力高于少组分群落,群落中禾草的竞争力显著增强,群落稳定性提高,混播群落中下繁草生长优势逐步增强,生产贡献率已逐步接近上繁草,群落结构趋于合理,尤其是根茎型品种(青海草地早熟禾)的生物量稳步增加。通过合理的品种组合和人工调控措施,使人工草地维持在高产可持续亚稳定状态,是高寒人工草地追求的结果和需要解决的科学问题。如何以适当的农业措施,便干预程度恰可保持人工草地在高生产水平上的稳定存在[30-32],即如何使建立的人工群落抵御恢复演替的生态压力,而保持在亚稳态状态[21,26],是黑土滩退化草地上建植多年生禾草人工草地可持续利用追求的目标。
4 结论
人工调控下,混播组合群落生物量与优良牧草比例比相应对照组合显著提高,随着组合草种增加,生长年限的延长,人工调控和品种组合优势合力体现,4组分及以上的高矮禾草混播组合的生产性能和群落稳定性显著高于2~3组分混播组合,群落生产力维持在较高的稳定状态。人工调控保持了群落组分种的绝对优势地位,比例维持在90%以上,对照中组分种的比例70%以上。
