黄土高原雨养农业区不同种植模式 土壤温室气体排放特征
2018-09-18邓长芳罗珠珠李玲玲牛伊宁蔡立群张仁陟谢军红
邓长芳,罗珠珠,*,李玲玲,牛伊宁,蔡立群,,张仁陟,谢军红
(1.甘肃农业大学资源与环境学院,甘肃 兰州 730070;2.甘肃省干旱生境作物学省部共建国家重点实验室,甘肃 兰州730070)
近年来,由温室效应带来的海平面上升、生物多样性减少及淡水资源减少等生态环境问题已成为全球性问题,受到人们广泛关注。作为主要温室气体的CO2、CH4、N2O控制着陆地生态系统中碳的数量、分布以及循环速度[1],对温室效应的贡献率约占80%[2],CO2排放量的增加更是导致全球变暖的主要因素[3]。与CO2相比,CH4的增温潜势大约是其23倍[4],而N2O的增温效应更高,约达到了CO2的150~200倍[5],具有更强的增温潜势。目前,国内外对温室气体的关注在工业化、城市化方面较多,忽视了农业低碳减排的必要性和紧迫性,而农田土壤是温室气体的重要排放源,约占温室气体总排放量的20%[6]。
甘肃省紫花苜蓿(Medicagosativa)占全国种植面积的1/3,一直居全国第1位[7],连作是其主要的种植方式,但长期的苜蓿连作会使土壤干燥化程度不断加剧,形成土壤干层且逐年加深[8],也会导致根系生物量减少,降低对土壤养分的利用强度[9-10],进一步造成土壤肥力下降进而影响苜蓿生产力。徐坤等[11]对不同年限苜蓿土壤CO2排放的研究表明,CO2排放通量随苜蓿种植年限的增加而增加。因此,在陇中黄土高原地区寻求合理的苜蓿种植方式,不但能够改善土壤质量,而且对控制农田土壤温室气体的排放也有重大意义。近年来,随着对黄土高原地区生态环境的关注,有关草田轮作的研究日益增多,合理的草田轮作方式不仅能改善土壤结构,调节土壤肥力,还能促进土壤生物化学过程。研究发现,苜蓿-作物轮作能降低土壤容重,增加土壤孔隙度,提高土壤饱和导水率,增加土壤入渗[12],且较之苜蓿连作可以增加土壤速效磷、速效钾、硝态氮、铵态氮等养分含量[13]。此外,通过调整种植方式对控制农田温室气体的排放也有巨大潜力[14]。目前,陇中黄土高原地区苜蓿-作物轮作系统的研究主要侧重于不同轮作模式对土壤理化性质的影响[15-16],对土壤温室气体排放的研究也大多是针对单一作物分别进行的[17],而同时测定不同作物土壤温室气体排放的研究匮乏。因此,本研究以陇中黄土高原旱作农田为研究对象,分别在作物生育期和休闲期进行温室气体排放观测,旨在探明该地区苜蓿后茬轮作不同作物条件下温室气体排放的季节变化规律,并探求同步观测的生物因子对气体排放通量变化的影响,以期为探明农田土壤温室气体排放特征、预测农田土壤温室气体排放的未来趋势和提高雨养农业系统可持续性提供理论参考。
1 材料与方法
1.1 研究地区概况
试区位于黄土高原半干旱丘陵沟壑区的定西市安定区李家堡镇。该区属于中温带半干旱区,平均海拔2000 m,年均太阳辐射592.9 kJ·cm-2,日照时数2476.6 h,年均气温6.4 ℃,≥0 ℃年积温2933.5 ℃,≥10 ℃年积温2239.1 ℃;无霜期140 d,年均降水390.9 mm,年蒸发量1531.0 mm,干燥度2.53,为典型的雨养农业区。该区土壤为典型的黄绵土,土质疏松,土层深厚,质地均匀,贮水性能良好。作物生育期内降水量见图1。
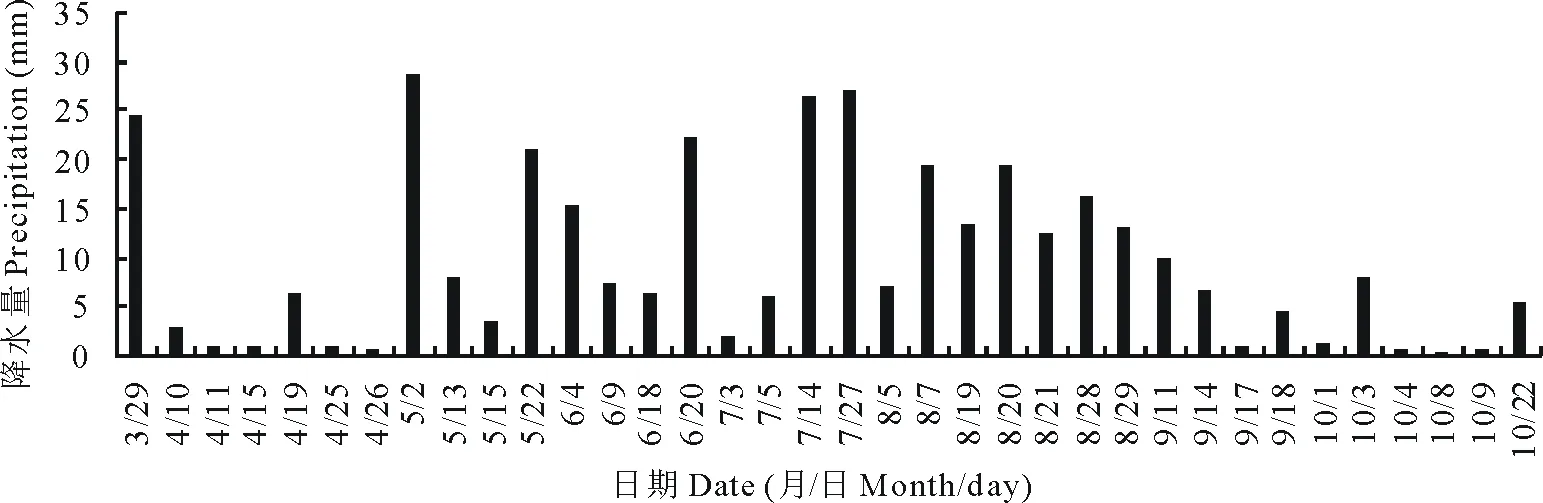
图1 作物生育期内降水量分布Fig.1 Distribution of the precipitation during crop growing season
1.2 试验设计
本试验共设6个处理(表1),3次重复,小区面积3.0 m×7.0 m,随机区组排列。供试作物为紫花苜蓿(L)、玉米(Zeamays)(C)、春小麦(Triticumaestivum)(W)、马铃薯(Solanumtuberosum)(P)、谷子(Setariaitalica)(M)。L-W、L-P、L-M均施纯氮肥105 kg·hm-2,纯P2O5105 kg·hm-2;L-C施纯氮肥200 kg·hm-2,纯P2O5105 kg·hm-2,L-L和L-F不施肥。所有肥料均在播种时施入,生育期没有追肥。小麦于2017年3月23日播种,7月22日收获;谷子于2017年4月17日播种,10月23日收获;玉米、马铃薯均于2017年4月28日播种,10月23日收获;苜蓿于2017年6月25日和10月23日刈割两次,各处理见表1。

表1 试验处理描述Table 1 Treatments discription
所有用地均是在2003年开始连续种植苜蓿8年。Alfalfa began to be planted in 2003 for 8 years.
1.3 气体样品采集及测定方法
1.3.1温室气体的采集与测定 气体采样时间为2016年10月至2017年10月,每隔15 d进行一次采样,气体采集时间均在上午8:30-11:30完成。N2O、CH4用静态箱密闭法采集,采样箱用1 mm 厚,304 K薄不锈钢板制成,箱体直径38 cm,高35 cm,底座内径36.5 cm,埋入各处理小区,整个采样期内不再移动。箱体外覆反光铝箔保温膜,箱顶有一胶塞测温口用于插入温度计读取箱内温度,箱内壁装有一个100 mm风扇用来混匀气体,箱体一侧安有三通阀采气孔并连接一根橡胶管用于注射器采样。采样时将采样箱扣入底座凹槽加水密封,箱顶插入温度计并接通风扇电源后用注射器分别采集0、10、20 min 时的气体样品注入150 mL铝塑复合气袋中,密封保存,每次采样结束后立即将样品带回实验室分析。N2O、CH4使用安捷伦7890A 型气相色谱仪(7890A GC System,USA)进行气体样品分析,色谱条件为色谱柱:Porapark Q 15 m×0.53 mm×25 μm,进样口150 ℃,分流进样,N2O检测器的检测温度为300 ℃,柱温45 ℃,柱流速3.3 mL·min-1,载气为高纯N2;CH4检测器的检测温度为200 ℃,柱温55 ℃,柱流速7.7 mL·min-1,载气为高纯N2;气体采集的同时,采用碳通量测量系统LI-8100(LI-COR,USA)及其自带20 cm直径呼吸室测定CO2呼吸速率,每小区测定3次,每次测定时间为90 s[18]。
1.3.2土样的采集及酶活性的测定 分别在2017年3月25日、7月25日、9月25日采集气样的同时取土样,取样深度为0~30 cm,每小区取3点,充分混合后带回实验室内进行测定。脲酶活性的测定采用苯酚次氯酸钠比色法,过氧化氢酶活性的测定采用高锰酸钾滴定法[19]。
1.3.3土壤水分 每次采集气体的同时通过烘干法(105±2) ℃测定 0~5 cm,5~10 cm,10~30 cm 三个土壤层的水分含量。
1.3.4土壤温度 在各小区分别安置一套地下5,10,15,20及25 cm地温计,每次采样时同步观察土壤温度。
1.4 主要计算方法
温室气体排放通量的计算:
F=(C2×V×M×273/T2-C1×V×M×273/T1)/[A×(t2-t1)×22.4]
(1)
式中:F代表温室气体(N2O,CH4)的排放通量(mg·m-2·h-1);A为采样箱的底面积(m2);V为采样箱体积(m3);M为被测气体的分子量;t1、t2分别为测定开始和测定结束的时间;T1、T2分别为采样箱密闭时和采样箱开启前的箱内温度(K);C1、C2分别为采样箱密闭时和采样箱开启前箱内被测气体的体积浓度。
温室气体累积排放量的计算:
M=∑(FN+1+FN)/2×(tN+1-tN)×24
(2)
式中:M为整个测定期内温室气体(N2O,CH4)的累积排放量(kg·hm-2);F为温室气体(N2O,CH4)的排放通量;N为采样次数;t为采样时间即初次采样的天数。
CO2累积排放量计算公式如下[20]:
CE=∑[1/2×(ti+1-ti)×(Rsi+1-Rsi)×0.1584×24]
(3)
式中:Rs为CO2排放速率;i+1与i表示两次相邻的测量;t表示播种后天数,0.1584为将碳排放数值单位μmol(CO2)·m-2·s-1转换为g(CO2)·m-2·h-1的系数,24为将碳排放数值单位由g(CO2)·m-2·h-1转换为kg(CO2)·hm-2的系数。
增温潜势(GWP)计算公式如下:
GWP=CO2的排放量×1+N2O的排放量×265+CH4的排放量×28
(4)
本研究以1 kg CO2的增温潜势为1,1 kg N2O和CH4的增温潜势分别为1 kg CO2的265倍和28倍[21],从而得出各处理下 CO2、N2O 和 CH4全年的综合增温潜势。
1.5 数据分析
采用Microsoft Excel 2007软件处理数据和制图,SPSS 19.0 统计软件对数据进行统计分析,差异显著性采用Duncan法,显著水平设置为0.05。
2 结果与分析
2.1 不同种植模式下土壤CO2排放通量年动态变化
图2是不同种植模式下土壤CO2排放通量的全年动态变化曲线。如图所示,各处理的变化趋势较为一致,均呈现出夏秋季高,春冬季低的季节性变化特征。从10月进入休闲期开始,随着土壤温度逐渐降低,CO2排放通量逐渐减少,在翌年1月达到全年排放量的最低值,这可能是在较低的土壤温度下土壤微生物的活跃程度下降引起的。至作物播种期,CO2排放通量缓慢增加,随着春季气温的回升以及作物播种后,CO2排放通量总体上呈逐渐上升的趋势,到6月5日L-L、L-W、L-C、L-F这4个处理出现了生育期内的第1个“峰值”,随后呈下降趋势。这是因为这一时期3种作物基本进入了快速生长期,营养生长与生殖代谢活动加快,植物光合作用和呼吸作用加强,其中,L-L处理下的CO2排放量更是达到了全年排放量的最高峰,可能是因为苜蓿一年内刈割2次,第1茬的苜蓿生物群体最大,土壤呼吸速率也最高,CO2的排放量也更多。到9月5日,除L-F和L-W处理外,其他几种作物出现了生育期内的第2个“峰值”,可能是由于这一阶段作物进入成熟期,生物量多,土壤微生物活动剧烈而导致CO2排放量较高。此后,随着作物的收获和温度的下降,土壤CO2排放通量逐渐减少,年均CO2排放量表现为L-L>L-M>L-C>L-P>L-W>L-F。
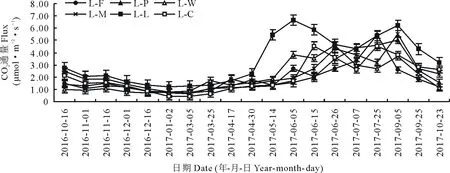
图2 不同种植模式下土壤CO2排放通量动态变化Fig.2 Dynamic variations of soil CO2 fluxes under different planting patterns
2.2 不同种植模式下土壤CH4吸收通量年动态变化
图3是不同种植模式下土壤CH4吸收通量的全年动态变化曲线。与土壤CO2排放通量的季节变化相似,CH4吸收通量也表现出夏秋季高,春冬季低的变化特征。夏、秋季节,土壤温度高、通气性好,CH4氧化菌的活性高,土壤吸收能力增强;冬、春两季土壤温度低,CH4氧化菌的活性降低,土壤吸收CH4能力明显减弱。从2016年休闲期至翌年播种前,由于受到土壤温度的限制,CH4吸收量保持在0.0078~0.0421 mg·m-2·h-1的较低水平,休闲期各处理吸收通量大小表现为L-W>L-M>L-P>L-C>L-L>L-F。至3月25日播种前,各处理CH4吸收量有明显降低,这可能是播种对土壤的扰动引起土壤气体的外排过程强于吸收过程,也可能是施肥导致土壤中氮含量增加,抑制了CH4的氧化过程,从而降低了土壤对CH4的吸收。随着春季气温回暖,土壤中CH4氧化菌活性增强,各处理下CH4吸收量开始逐渐升高,6、7月,各处理CH4吸收通量达到全年最大值,总体保持在0.0507~0.1348 mg·m-2·h-1的较高水平,主要原因在于这一时期温度高蒸发量大,土壤处于好气状态,促进了土壤对CH4的吸收。6月5日吸收量急剧降低,这可能是因为前一天降水15.29 mm,厌氧条件下CH4菌活跃,氧化菌活性降低,从而抑制了CH4的吸收过程,6、7月各处理的吸收总量表现为L-W>L-M>L-C>L-P>L-L>L-F。进入成熟期后(9月5日),土壤温度降低,微生物活动减弱,土壤CH4吸收通量逐渐减少,各处理年均CH4吸收量表现为L-W>L-M>L-P>L-C>L-L>L-F。
2.3 不同种植模式下土壤N2O排放通量年动态变化
不同种植模式下N2O排放通量变化趋势一致,总体呈现出降低-升高-降低的单峰变化趋势(图4),整个生育期内,苜蓿连作表现出高吸收高排放的特点,这应该与其发达的根系有较大关系。10月收获后进入越冬期,土壤N2O呈吸收排放状态,各处理下的N2O排放量逐渐降低并保持在-0.0314~0.0456 mg·m-2·h-1的范围内,休闲期内各处理N2O排放通量表现为L-F>L-C>L-M>L-P>L-W>L-L。3月随着气温回升,N2O排放量逐渐升高,至7月水热适宜,作物生长旺盛,N2O排放量达到了全年峰值,各处理排放量表现为L-L>L-P>L-M>L-W>L-C>L-F,随后呈明显的下降趋势,这可能是因为作物成熟后,对土壤养分、水分消耗过多,抑制了土壤中N2O的扩散与释放。各处理年均N2O排放量表现为L-C>L-M>L-F>L-L>L-W>L-P。

图3 不同种植模式下土壤CH4吸收通量动态变化Fig.3 Dynamic variations of soil CH4 fluxes under different planting patterns

图4 不同种植模式下土壤N2O吸收通量动态变化Fig.4 Dynamic variations of soil N2O fluxes under different planting patterns
2.4 不同种植模式对CO2、CH4和N2O累积排放量和增温潜势的影响
从表2可以看出,不同种植模式对CO2、CH4和N2O 3种温室气体的排放(吸收)量以及增温潜势有显著地影响。从3种气体的累积排放(吸收)量方面看,CO2的累积排放量在L-L处理下最高,苜蓿-作物轮作的处理里L-W最低,较L-L处理显著减少了42.43%;CH4的累积吸收量在L-F处理下最低、L-L次之,而轮作的作物中L-M最高,分别较L-F和L-L增加了62.36%和31.69%,L-M与L-W处理间差异不显著;N2O的累积排放量在L-C处理下最高,L-M处理次之,L-P和L-W处理最低,且处理间没有显著差异。这说明与苜蓿-作物轮作相比,苜蓿连作处理的CO2排放量高,CH4吸收量少,而苜蓿-小麦可有效减少CO2的排放,增加CH4的吸收。
从对大气的增温潜势方面看,苜蓿连作(L-L)处理CO2的增温潜势显著高于苜蓿-作物轮作的其他处理,且提高了49.57%~81.16%;在轮作处理中,L-F处理的CO2增温潜势最低,较最高的L-M处理下降了17.44%,L-F与L-W处理间差异不显著;N2O增温潜势最低的是L-P处理,较最高的L-C处理下降了19.44%,L-P与L-W处理间差异不显著;CH4增温潜势最低的是L-F处理,而在轮作处理中增温潜势最低的是L-C,L-C与L-W处理间差异不显著。综合增温潜势表现为L-L>L-M>L-C>L-P>L-F>L-W,这说明苜蓿多年连作后,对大气温度的变化有明显的增温效应,而苜蓿后茬轮作粮食作物可有效缓减大气温度的升高,并以苜蓿后茬轮作小麦效果最佳。
2.5 不同种植模式对土壤酶活性的影响
不同生育期内0~30 cm土层各处理脲酶和过氧化氢酶活性如表3所示。两种酶活性在不同生长阶段变化趋势基本一致,均呈现出先升高后降低的趋势。播种期(3月25日)脲酶活性均值为2.29 mg·g-1,苜蓿-作物处理中L-W处理显著低于L-L和L-C处理,分别较之降低了10.29%和7.63%;至作物生长期(7月25日),脲酶活性均值比播种期提高了20.12%,苜蓿-作物处理中,L-M处理最高,较L-P、L-W处理分别增加4.43%、8.02%,差异显著;至作物成熟期(9月25日),L-C处理的脲酶活性略低于L-L处理,较L-P、L-W、L-M分别提高2.33%、4.76%、8.20%。过氧化氢酶活性在播种期均值为1.34 mL·g-1,L-P处理较L-M处理提高17.97%,与L-C、L-W无显著差异;生长期L-L处理过氧化氢酶活性较其他处理提高5.56%~26.67%,与L-P、L-W、L-M处理差异显著;成熟期,苜蓿连作的过氧化氢酶活性显著高于其余处理。
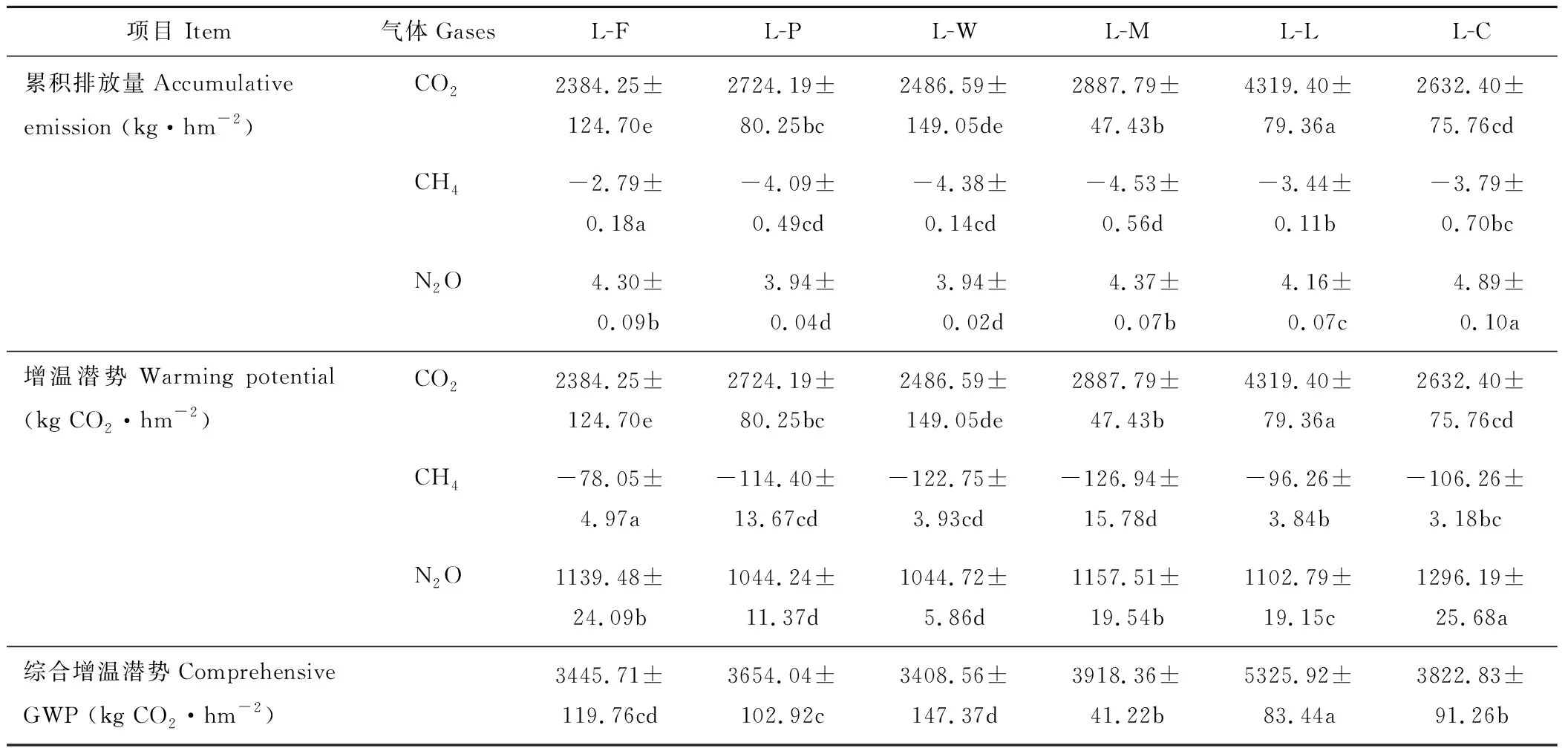
表2 不同种植模式下温室气体累积排放量及增温潜势Table 2 Greenhouse gas accumulative emission and comprehensive global warming potential (GWP) under different planting patterns
同行不同小写字母表示处理间差异显著(P<0.05)。下同。
Different small letters in the same row indicate significant difference among treatments at 0.05 level. The same below.

表3 不同种植模式下0~30 cm土层的酶活性Table 3 Soil enzyme activities under different planting patterns in 0-30 cm
2.6 不同种植模式对土壤水分和温度的影响
从测定的温度数据可以看出(图5),在完整的一个年周期内,0~30 cm土层的温度受季节变化的影响,呈夏秋高、春冬低的变化规律,对比不同轮作处理全年0~30 cm土层温度变化可以看出,在整个休闲期内,各轮作处理之间温度变化的趋势以及温度范围基本上保持一致。不同种植模式对土壤温度的影响主要集中在各作物的生育期内,从3月25日(播种期)至9月25日(成熟期),各轮作处理的土壤温度均保持在较高范围,这一阶段0~10 cm土层温度分别在6月5日和7月7日达到两个波峰,6月5日的平均气温在28~32 ℃,7月7日的平均气温在26.5~32.5 ℃;而在10~30 cm土层下,L-L处理的温度明显高于其他处理。从全年范围内不同土层平均温度来看,0~5 cm土层温度均表现为L-M>L-C>L-W>L-F>L-P>L-L,5~10 cm和10~30 cm土层温度表现为L-C>L-M>L-F>L-W>L-P>L-L。到10月下旬,各处理间温度变化的差异逐渐变小,10~30 cm土层温度基本没有差异。
从0~30 cm土层的水分含量动态变化来看(图5),各处理0~5 cm土层水分含量在4月中下旬达到全年最小值,变化范围在1.95%~9.72%,其中L-C处理最高,L-P处理最低,这与该地区这一时期春旱频次高、强度大的季节特征有关,而L-C处理水分较高是因为地膜具有极强的保墒作用,抑制了土壤水分的蒸发;各处理土壤水分含量较高的时期出现在9月5日,保持在19.41%~23.27%,其中L-F处理最低,L-P处理最高,这是因为8月降水较多,尤其是8月28、29日降水量达到29.5 mm。从土层深度上看,0~10 cm的水分由于受气象因素的影响波动较大,10~30 cm变化幅度较小,而各处理的土壤水分在相同土层内的变化趋势较为一致,0~30 cm土层的全年平均含水量表现为:L-C>L-F>L-P>L-M>L-W>L-L。

图5 不同种植模式下0~30 cm土层的土壤温度和土壤水分Fig.5 Soil temperature and water content in 0-30 cm soil layer under different planting pattern
2.7 土壤酶及环境因子对温室气体排放通量的影响
表4和图6是0~30 cm土层土壤温度、土壤水分、土壤酶与温室气体排放量的回归分析。结果表明,土壤温度与CO2、CH4、N2O排放通量之间都呈现极显著正相关关系,CO2排放通量与0~10 cm土层土壤含水量显著正相关(r=0.54~0.56),CH4排放通量与0~5 cm土壤含水量显著负相关(r=-0.50),N2O排放通量与5~10 cm土层水分显著正相关(r=0.51);温室气体排放通量与土壤酶活性之间有极显著线性相关关系,脲酶与CO2、N2O、CH4的排放通量相关系数r分别为0.84、0.85和-0.78(P<0.01),过氧化氢酶与CO2、N2O和CH4的排放通量相关系数r分别为0.71、0.57和-0.70(P<0.05),这说明土壤温度升高促进温室气体的排放,不同土层的水分对温室气体排放的影响效果有差异,而CO2和N2O排放通量随脲酶和过氧化氢酶活性的升高而增加,CH4排放通量随酶活性的升高而减少,吸收通量增加。
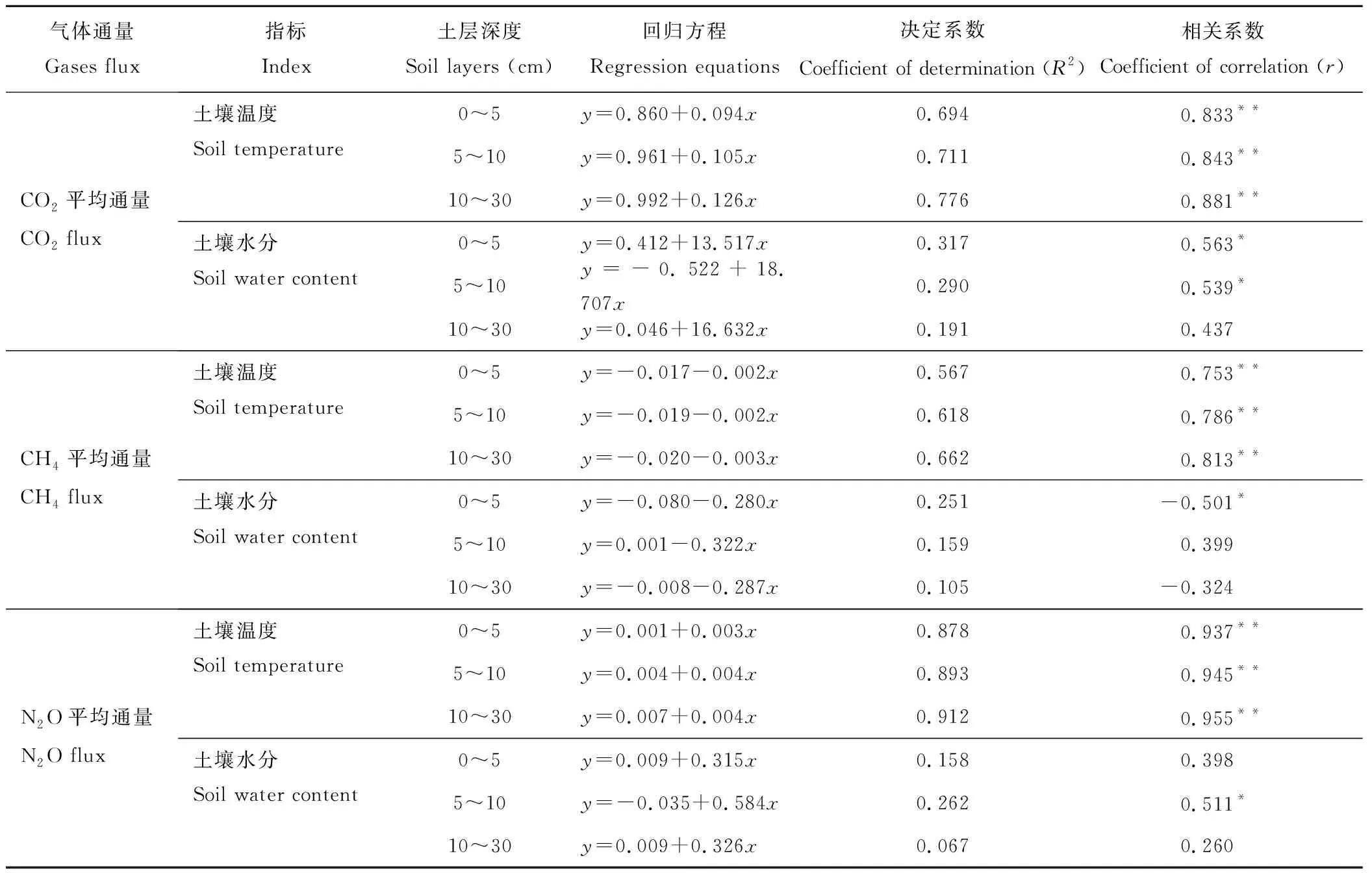
表4 不同种植模式下土壤温室气体与不同土层土壤温度、土壤水分的回归方程Table 4 Regression equations between greenhouse gases (y) and soil water content and temperature of different layers (x) under different planting patterns
*:P<0.05;**:P<0.01.
表5是利用逐步回归分析法建立了0~30 cm土壤温度、土壤水分、土壤酶影响温室气体排放通量的最优回归模型。结果表明,土壤温度、过氧化氢酶对CO2和CH4排放量的影响较大;在N2O排放量的回归模型中,土壤生物因子相对而言对气体排放的影响较小,环境因子对N2O的排放有极显著影响。这说明农田温室气体的排放是诸多因素共同作用的结果,主导不同气体排放量的因素也不尽相同,本研究中CO2和CH4的排放受到环境因子和生物因子的共同影响,而N2O的排放则与土壤温度、水分关系较为密切。
3 讨论
土壤温室气体的产生是土壤微生物等在众多环境因子综合作用下分解土壤有机物质和植物根系等共同作用的结果,其强度受到土壤微生物类群数量及活性、土壤有机质数量及矿化速率、土壤动植物呼吸等的共同影响[22]。本研究中,不同种植模式下温室气体排放量呈夏秋高,春冬低的季节性变化特征,这与宋敏等[23]对旱作农田温室气体的研究结果较为一致,春冬休闲期内土壤温度低,微生物活性以及土壤呼吸速率降低,夏秋季节光温条件适宜,微生物呼吸和植物根系呼吸加强,加速了土壤中有机质的分解和微生物活性,而且大量根系分泌物及植物凋落物输入到土壤, 为微生物提供了足够的底物反应基质。越冬期内, 不同种植模式下土壤对N2O呈吸收排放态,这与蔡立群等[14]研究黄土高原旱作区土壤N2O在整个测定期内均为排放源的结果不一致。虽然两研究位于同一试区,但种植作物不同,土壤环境存在较大差异,苜蓿根系发达且为固氮植物,冬季温度低、根瘤菌活性弱, 可能需要从土壤中吸收一定的矿质氮来满足植物体对氮的需求, 这与其他苜蓿地土壤N2O排放研究的结果[24-25]相似。由此可见,不同种植模式下作物所处的温度水分及根际环境的差异会引起土壤气体排放的变化,有研究发现,陇东黄土高原不同作物CO2排放量表现为苜蓿>玉米>草地>小麦[26]。本研究结果表明,苜蓿-苜蓿连作处理的CO2累积排放量最高,苜蓿-小麦处理较之降低了42.43%,CH4的吸收量也显著低于苜蓿-作物轮作处理,这说明苜蓿-作物轮作系统较苜蓿-苜蓿连作能不同程度地降低土壤CO2排放量,提高CH4的吸收量。 裴淑玮等[27]对玉米-小麦轮作农田N2O的研究Y1、Y2、Y3分别是CO2、CH4、N2O的排放通量;X1、X2、X3、X4分别是土壤温度、土壤水分、脲酶、过氧化氢酶。Y1,Y2andY3are emission fluxes of CO2, CH4and N2O respectively;X1,X2,X3andX4are soil temperature, soil moisture, urease activities and catalase activities respectively.

图6 不同轮作模式下土壤酶活性与温室气体排放通量的关系Fig.6 Relationship between soil enzymes activities and greenhouse gas fluxes under different planting patterns

表5 0~30 cm土壤水分、土壤温度、土壤酶与 温室气体排放量的逐步回归分析Table 5 Multiple stepwise liner analysis between soil moisture, soil temperature, soil enzymes of 0-30 cm and greenhouse gases emissions
表明玉米季N2O的排放量占全年排放量的57%~86%。本研究中,L-C处理的N2O累积排放量显著高于其他处理,比L-W处理提高24.11%,玉米生长期正值高温多雨季,土壤温湿度高于小麦生长期,为土壤反硝化营造了良好的条件,促进了反硝化过程的发生,提高N2O的产生速率。温室效应的加剧主要依赖于CO2、 N2O和CH4等温室气体的排放,本研究不同处理下的综合增温潜势表现为L-L>L-M>L-C>L-P>L-F>L-W,由此可见,苜蓿-小麦轮作能更有效缓减大气的增温效应,因为小麦生长期较短,避免了夏季部分高温多雨的天气,大大减少了生育后期CO2和N2O的排放。此外,小麦进入收获期较早,避开了降水充沛期,土壤CH4氧化菌活性高,削弱了CH4的排放,强化了小麦土壤对CH4的吸收。
农田温室气体的排放过程极为复杂,是土壤内环境各个部分的综合效应向外界反馈的过程。土壤酶是微生物活性的表征,不仅影响着土壤中生化反应速率及各种氧化还原反应,又与作物根系生长发育及土壤温室气体排放密切相关[28],土壤温度、水分的变化对温室气体的排放也有重要的影响。许多学者都做了关于土壤环境因子、生物因子对温室气体排放方面的研究,一些研究认为,土壤温度升高时不仅会加快土壤CO2的释放[29-30],还会通过影响土壤中微生物活性、化学反应来影响CH4和N2O的吸收和排放[31]。本研究中,温室气体的排放与0~30 cm土壤温度显著正相关,各处理中L-L处理土壤温度最低,这说明草粮轮作有一定的增温效应,但轮作小麦和马铃薯增温效果相对较低;0~10 cm土壤水分越高越会促进CO2和N2O的排放,0~5 cm水分增加会降低土壤CH4的外排。而本试验处理中,轮作小麦较之轮作其他作物能明显降低土壤水分含量;土壤中的脲酶和过氧化氢酶过高时虽然能减弱CH4的排放过程,但也会显著增加CO2和N2O的排放量,本研究中整个生育期内苜蓿连作处理的两种酶活性均高于其他处理,差异显著,这是因为苜蓿根系较轮作处理根系发达,有机质转化较为强烈的原因,苜蓿的固氮作用进一步导致了土壤氮素的增高。因此,较之轮作其他作物,轮作小麦能更有效降低两种酶活性。此外,本研究还发现,影响不同温室气体排放的主导因子也不尽相同,CO2和CH4的排放受土壤温度和过氧化氢酶的影响较大,而N2O的排放与环境因子的关系更加密切。
4 结论
在陇中黄土高原雨养农业区,与苜蓿连作相比,苜蓿种植多年后翻耕轮作粮食作物可明显降低农田综合增温潜势,其中以轮作小麦的效果最好,这主要是由于苜蓿-作物轮作后能促进土壤温度和土壤水分的增加,降低土壤中脲酶和过氧化氢酶活性,这有利于降低土壤中CO2和N2O的排放量,减少土壤中CH4的外排和对大气中CH4的吸收,有效减弱了农田温室气体的增温效应。因此,连续多年种植苜蓿后应该及时进行倒茬轮作,不仅有利于改善土壤水温特性,提高土壤质量,而且对节能减排及改善大气环境都有重大意义。
