生长素对荠菜心皮发育的影响
2018-09-11刘晓柱李银凤张学文
刘晓柱,李银凤,赵 燕,张学文*
(1.贵州理工学院 食品药品制造工程学院,贵州 贵阳 550003; 2.湖南农业大学 生物科学技术学院,湖南 长沙 410000)
植物花器官发育是植物生长周期的重要过程,对于大多数农作物而言不仅是决定其产量和经济价值形成的重要阶段,也是植物学界研究的重要领域[1]。对于植物而言,器官形态的最终形成取决于建立起形态模式后对细胞分裂方向和分裂频次的控制和分化,而生长素(Auxin)在这一过程中起决定性作用[2]。
近年来,关于拟南芥雌蕊发育及模式建成的研究十分活跃,已从遗传分析和激素调控两方面获得了深入的认识[3]。Nemhauser等[4]发现并提出了雌蕊中顶端至基部生长素浓度梯度模型,在该模型中雌蕊顶端存在高浓度的生长素,促进了柱头和花柱的伸长和分化,子房中存在中间浓度的生长素,雌蕊底端的花托中存在低水平的生长素。提高顶端生长素浓度可弥补一些花柱促因子的缺失。心皮是构成雌蕊的基本单位,说明心皮发育过程中存在生长素浓度梯度的分布。Sundberg等[5]利用DR5∶∶GFP系统对拟南芥花器官发育过程生长素的分布研究指出,在雌蕊发育过程中,柱头产生高浓度的生长素,并维持一种顶端至基部的极性分布,子房中存在中间浓度的生长素,胚珠中产生低水平的生长素,保证胚珠的发育成熟。受精完成后,胚胎中生长素迅速增加。
生长素通过运输载体进行极性运输实现其在植物体内的差异分布,而这种差异分布是调控植物生命过程的必要先决条件[6]。PIN家族是目前一种公认的生长素输出载体,极性分布在细胞膜上,负责生长素的内外运输,调控生长素浓度梯度动态平衡[7-8]。拟南芥、水稻、苹果、麻疯树、狗蔷薇等的PIN基因已被克隆, 并进行了深入研究,证实了其参与植物器官的生长发育、向性运动、胚胎发育等过程[9-11]。
荠菜为十字花科1年生草本植物,在生理特性和遗传背景上与拟南芥非常接近,但心皮形态却差异巨大:荠菜为心形心皮结构,而拟南芥为柱形心皮结构。造成荠菜心皮形态的独特性机制还不清楚,推测可能与生长素的极性分布有关,因此构建生长素报告系统DR5∶∶GFP,探析生长素在荠菜心皮发育中的极性分布,利用RT-PCR克隆荠菜PIN3(CbPIN3)基因编码区,并进行生物信息学分析以及表达分析,旨在为揭示荠菜心皮形态建成奠定分子基础。
1 材料和方法
1.1 菌株、质粒及植物材料
大肠杆菌(Escherichiacoli)DH5α、根癌农杆菌(Agrobacteriumtumefaciens) GV3101、质粒pEGAD、pBI121均由湖南农业大学细胞生物学实验室保存。
野生型荠菜采自湖南农业大学耘园教学实习基地。
1.2 试剂及试剂盒
蛋白胨、甘油、琼脂、常用抗生素购自上海生工生物有限公司;磷酸氢二钾、硫酸镁、氯化镁、无水乙醇、冰醋酸等均为分析纯,购自广州化学试剂有限公司;TaqDNA聚合酶、T4 DNA 连接酶、各种限制性内切酶、DNA分子质量标准均购自宝生物工程(大连)有限公司;质粒提取试剂盒、胶回收试剂盒购自天根生化科技(北京)有限公司。
各种抗生素贮存液均按《分子克隆实验指南》[12]配制。
1.3 培养基
本研究所用LB、YEB、1/2 MS培养基的配制均参考《分子克隆实验指南》[12]。
1.4 试验方法
1.4.1 生长素报告系统DR5∶∶GFP的构建 参照Ulmasov等[13]方法,设计了生长素特异响应启动子DR5,该启动子序列由3部分组成:生长素响应元件DR5、35S minimal promoter、TMV leader(Ω′)。DR5启动子全序列设计如下:
AAGCTTCTATACTAAGTTCATGATAATAGTTGCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCCTTTTGTCTCCAAGACCCTTCCTCTATATAAGGAAGTTCATTTCATTTGGAGAGGATATTTTTACAACAATTACCAACAACAACAAACAACAAACAACATTACAATTACTATTTACAATTACATCTAGA。
DR5启动子由上海英骏生物公司合成。
采用HindⅢ、XbaⅠ双酶切DR5以及载体pBI121,回收酶切产物,T4 DNA连接酶16 ℃过夜反应,然后转化DH5α感受态细胞,涂布在含有50 μg/L卡那霉素(Kan)的LB培养基上,挑选白色克隆。碱法小批量抽提法抽提质粒,酶切检测后送测序,构建正确的载体命名为pBI121-DR5。
采用高保真酶扩增GFP基因,BamHⅠ、SacⅠ双酶切GFP扩增产物以及载体pBI121-DR5,回收纯化酶切产物,T4 DNA连接酶16 ℃过夜反应,然后转化DH5α感受态细胞,涂布在含有50 μg/L Kan的LB培养基上,挑选白色克隆。碱法小批量抽提法抽提质粒,酶切检测后送测序,构建正确的载体命名为pBI121-DR5∶∶GFP。本研究所用引物见表1,由北京华大基因生物有限公司合成。
1.4.2 生长素报告系统DR5∶∶GFP转化野生型荠菜 利用冻融法将载体pBI121-DR5∶∶GFP转化至根癌农杆菌GV3101中,通过菌落PCR检测后,接种至新鲜YEB培养基中(利福平 20 mg/L+庆大霉素30 mg/L+Kan 50 mg/L),培养24 h。当野生型荠菜植株生长至主花序10 cm左右,次花序在莲座开始形成时,剪掉已开花或将要开花的花苞,进行荠菜的遗传转化,收取成熟转基因荠菜种子进行筛选检测。
1.4.3CbPIN3基因的克隆 严格按试剂盒说明书步骤提取荠菜叶片RNA,采用甲醛变性胶电泳与核酸紫外分光光度计检测其质量与浓度,然后反转录成cDNA。根据拟南芥PIN3基因(NM_105762.3)核苷酸序列,设计PCR克隆引物(表1)。
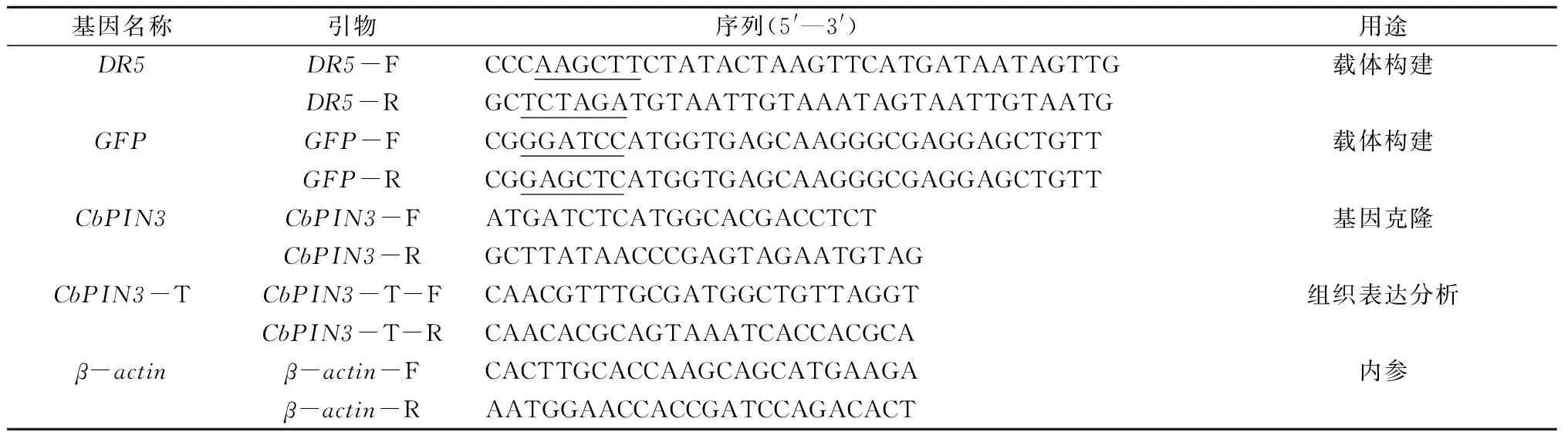
表1 研究所用引物
注:下划线部分为酶切位点。
以荠菜cDNA为模板进行PCR扩增,PCR反应体系为:cDNA模板1 μL, PCR Buffer(10×) 2.5 μL,dNTPs (10 mmol/L) 1 μL,引物CbPIN3-F/R (10 μmol/L)各1 μL,TaqDNA聚合酶1 U,补水至总体积25 μL。PCR反应程序为: 95 ℃预变性3 min;94 ℃变性1 min,55 ℃退火1 min,72 ℃延伸2 min,35个循环;最后72 ℃终末延伸10 min。将扩增产物切胶回收纯化后,连接T载体,转化DH5α感受态细胞,涂布在含有IPTG和X-gal的氨苄青霉素LB培养基上,挑选白色克隆。抽提质粒,经酶切检测后送测序。
1.4.4CbPIN3生物信息学分析 采用NCBI BLAST在线分析CbPIN3基因核苷酸序列同源性;采用DNAStar软件分析CbPIN3开放阅读框、氨基酸序列组成、理化性质;采用蛋白质分析系统(http://www.expsy/ch/tools)分析CbPIN3蛋白结构特征;采用ClustalX与MEGA软件构建CbPIN3蛋白系统进化树。
1.4.5CbPIN3组织表达分析 提取荠菜根、茎、叶、花、种子部位总RNA,反转录成cDNA。qRT-PCR检测CbPIN3基因表达水平。扩增体系:cDNA 2 μL,上下游引物(10 μmol/L)各0.8 μL,SYBR Green Mix(2×)10 μL,补水至总体积20 μL。扩增程序:95 ℃预变性 3 min;95 ℃变性10 s,60 ℃退火30 s,35个循环,最后72 ℃终末延伸10 min。以β-actin为内参,按照2-ΔΔCt方法分析CbPIN3相对表达量。
2 结果与分析
2.1 生长素报告系统DR5∶∶GFP的构建
鉴于荠菜与拟南芥心皮形态差异显著(图1),为分析生长素在荠菜心皮发育中的作用,构建了生长素报告系统DR5∶∶GFP,该系统包含DR5启动子与GFP报告基因2个部分。首先人工体外合成DR5,连接到载体pBI121上,替换35S启动子,然后将GFP基因构建到载体pBI121上,双酶切检测GFP基因,最终成功构建了生长素报告系统DR5∶∶GFP(图2)。
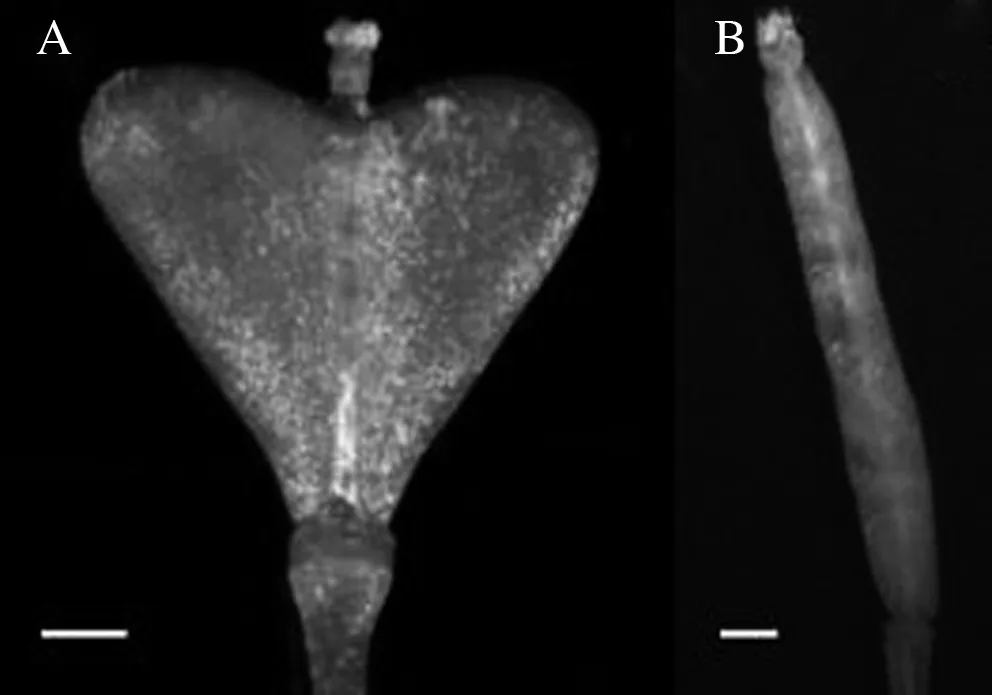
A:荠菜成熟期心皮;B:拟南芥成熟期心皮;bar=1 mm
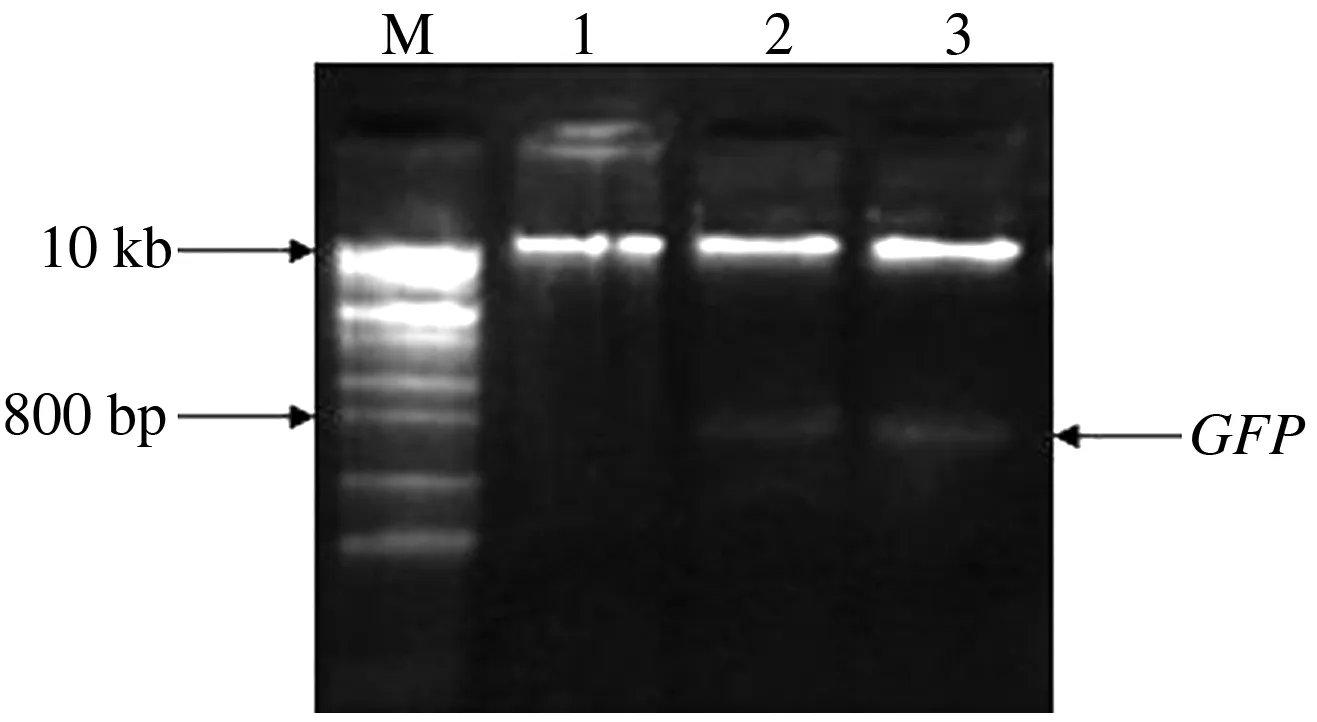
M:1 kb plus DNA分子质量标准;1—3:pBI121-DR5∶∶GFP酶切结果
2.2 转生长素报告系统DR5∶∶GFP荠菜植株的获得
将构建好的生长素报告载体pBI121-DR5∶∶GFP通过冻融法转化至根癌农杆菌GV3101中,农杆菌通过花序侵染方式转化野生型荠菜,收取转基因荠菜种子,播种于盆装培养土中,待长到2片真叶时喷洒50 mg/L的Kan进行筛选,得到具有抗性的转基因植株(图3),温室内培养至成熟结种,单株收种子得到T1代种子。T1代种子经消毒,春化后再次种植,经过性状分离筛选,得到T2代纯合荠菜植株。提取Kan抗性荠菜总DNA,PCR法检测GFP基因,结果4株扩增出GFP目标条带(图4),最终成功获得转生长素报告系统DR5∶∶GFP荠菜植株。

图3 转基因荠菜Kan抗性筛选
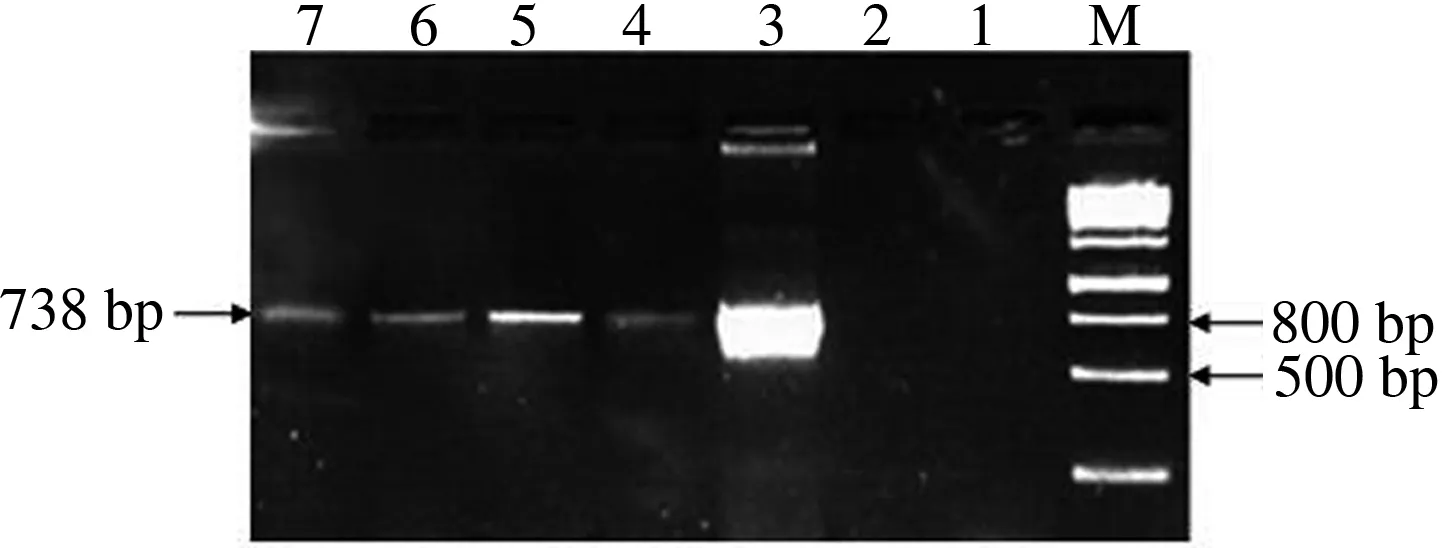
M:1 kb plus DNA分子质量标准;
2.3 转基因荠菜心皮发育过程中生长素的检测
荧光显微镜观察转基因荠菜花器官的GFP荧光,结果发现,荠菜花器官发育过程中生长素分布的特点如下:花瓣中观察到较低的荧光信号;雄蕊产生较强的荧光信号;柱头产生高强度的荧光信号;胚珠中有较弱的荧光信号(图5)。
2.4 荠菜生长素输出载体PIN3基因的克隆
在拟南芥的向性反应中,PIN3动态分布参与生长素的极性运输,包括侧向运输[14]。由于荠菜心皮发育中生长素的极性分布特点,通过RT-PCR方法获得了1条2 000 bp的特异性条带(图6)。测序结果表明CbPIN3基因编码区全长1 950 bp,BLAST比对发现,所克隆的序列与拟南芥、印度芥菜的PIN3基因及其他的PIN基因具有较高的同源性,表明已成功克隆了荠菜PIN3基因,命名为CbPIN3,并提交GenBank(JF966673)。
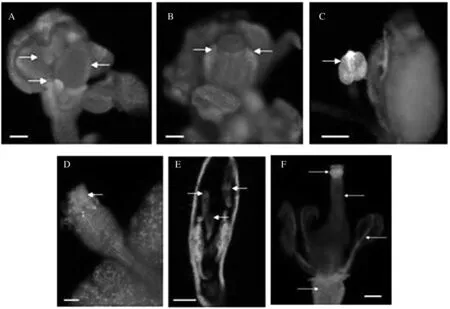
A:GFP在荠菜花瓣中表达;B:GFP在荠菜心皮两侧上端表达;C:GFP在荠菜雄蕊中表达;

M:1 kb plus DNA 分子质量标准;
2.5 CbPIN3蛋白结构分析
DNAStar软件翻译结果显示,CbPIN3可编码649个氨基酸(图7),蛋白质分子质量为70.00 ku,等电点为8.14。碱性氨基酸(K、R)个数为48;酸性氨基酸(D、E)个数为46;疏水氨基酸(A、I、L、F、W、V)个数为247;极性氨基酸(N、C、Q、S、T、Y)个数为172;脂肪族氨基酸(I、L、V)个数为145;芳香族氨基酸(F、W、Y)个数为61。
在拟南芥中PIN家族关键位点的磷酸化修饰,对其发挥生长素极性运输作用至关重要[15]。因此利用NetPhosBac 1.0 Server在线预测CbPIN3蛋白磷酸化情况,结果如图8所示。CbPIN3蛋白存在13个丝氨酸磷酸化位点,1个苏氨酸磷酸化位点,说明CbPIN3蛋白功能的发挥可能也需要磷酸化修饰。
跨膜结构的存在对于生长素运输载体发挥功能起着重要作用,因此预测了CbPIN3蛋白跨膜结构情况。结果表明,CbPIN3具有9个跨膜结构域,其中N端5个,C端4个。在C端还有一个整合蛋白,定位于质膜的细胞外表面(ES)(图9)。在线预测了CbPIN3亚细胞定位,结果显示,CbPIN3在细胞膜上进行表达。
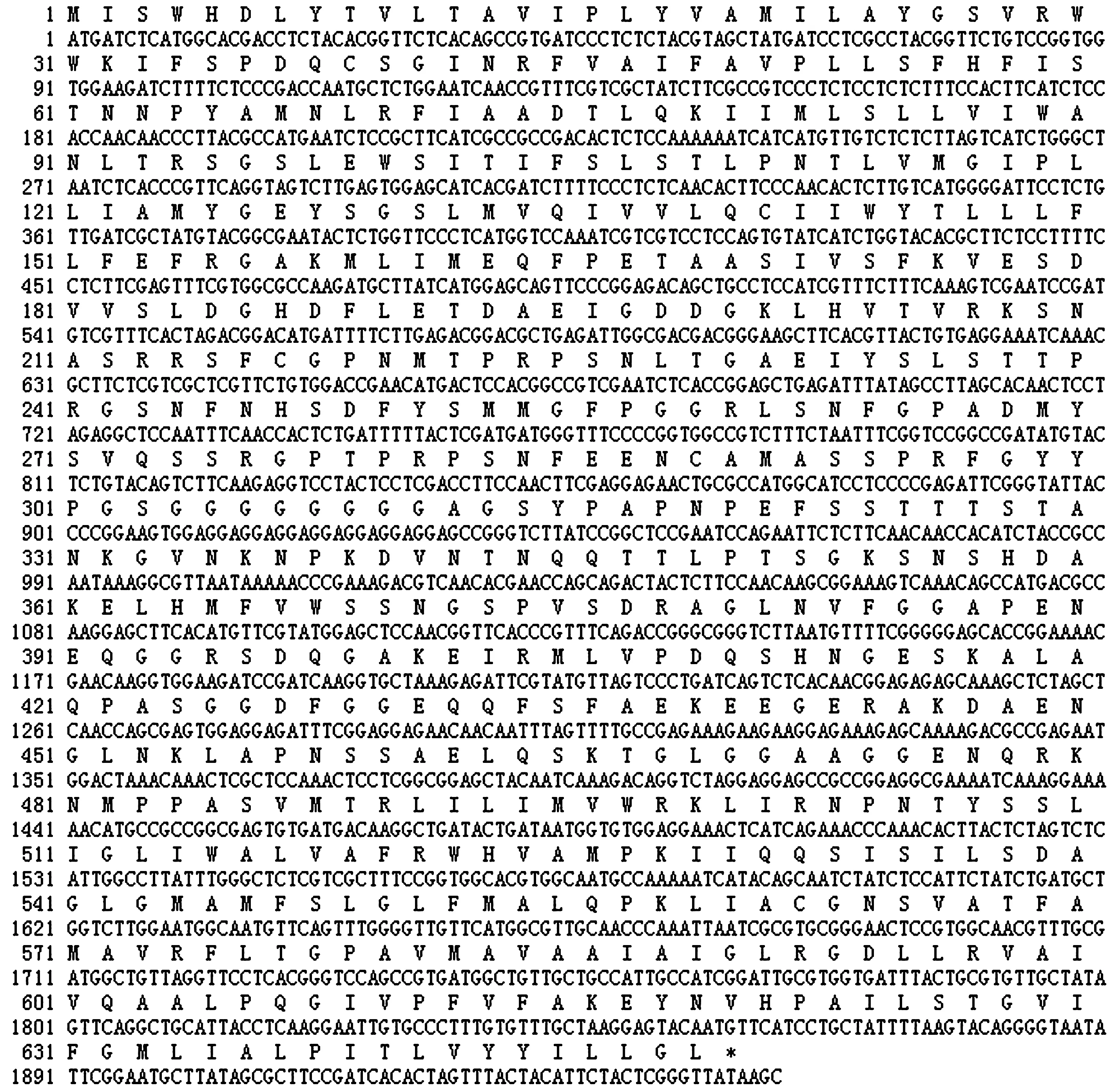
图7 CbPIN3基因核苷酸序列及推测氨基酸序列

图8 CbPIN3蛋白磷酸化位点预测
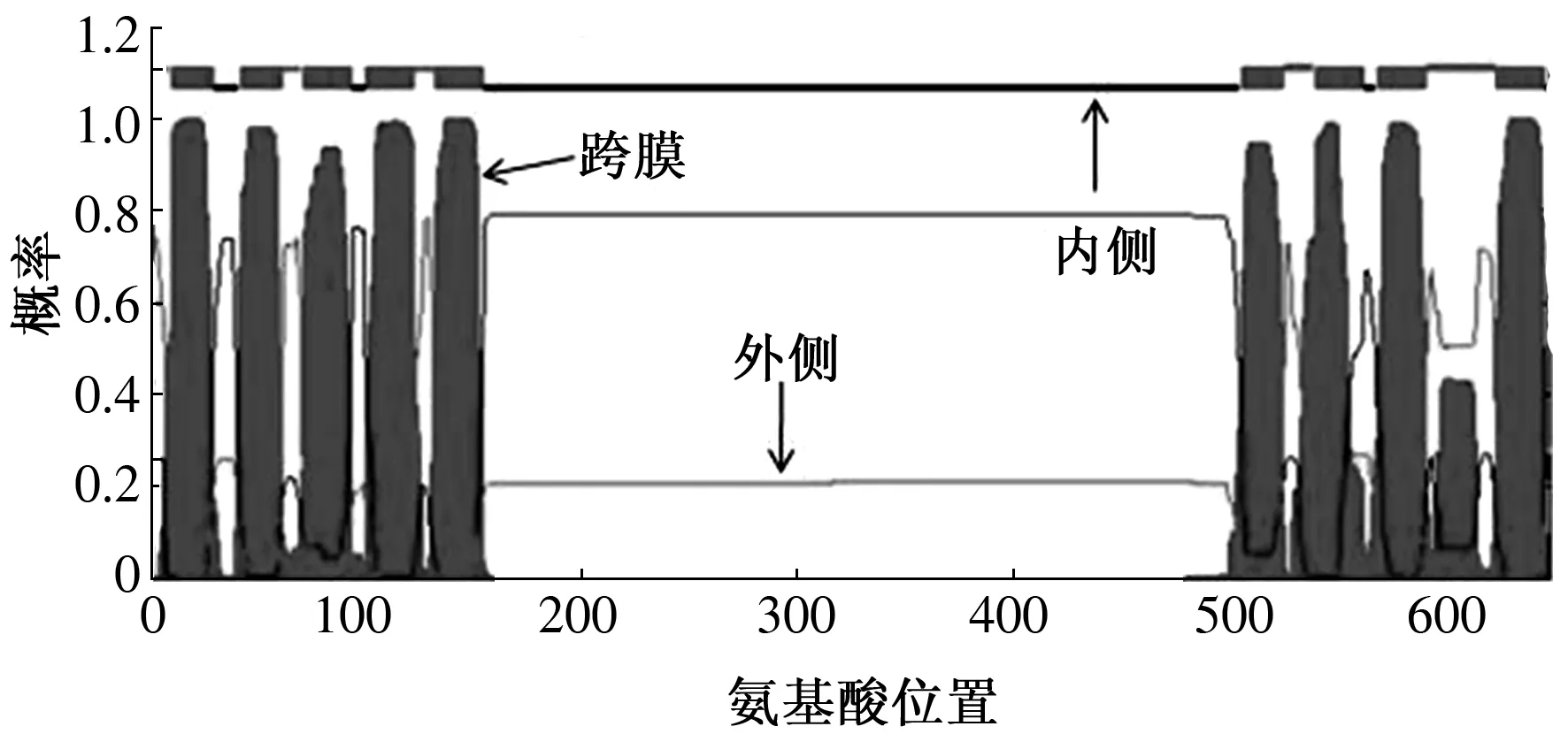
图9 CbPIN3蛋白跨膜区预测分析
2.6 CbPIN3蛋白进化分析
为进一步分析CbPIN3蛋白与其他植物PIN3蛋白的进化关系,利用MEAG 5.1构建PIN3系统进化树,结果如图10A所示,CbPIN3与拟南芥PIN3(AtPIN3)亲缘关系最近,进化树上属同一支;亲缘关系最远的是菠萝PIN3(AcPIN3)。
进一步分析CbPIN3蛋白与拟南芥PIN3蛋白结构,发现二者同源性高达90%以上,二者N端和C端都具有几乎一致的保守跨膜结构域,区别主要在于中间环状结构。根据亲水环数目的差异将拟南芥PIN蛋白分为长PIN蛋白与短PIN蛋白2个亚家族。在长PIN蛋白家族中,CbPIN3与AtPIN3亲缘关系最近,在进化树上同属于一支;其次与AtPIN7关系较近;与AtPIN6亲缘关系最远(图10B)。
2.7 CbPIN3组织表达分析
为分析CbPIN3基因表达模式,以qRT-PCR分析荠菜不同组织CbPIN3表达情况。如图11所示,CbPIN3基因在荠菜根、茎、叶、花、果荚及种子中均有表达,但表达量具有组织差异性。其中,表达量最高的是茎,其次是根、叶及花,果荚中表达量最低,因此,推测CbPIN3可能参与调控荠菜茎、根、叶、花、果实等的发育过程。
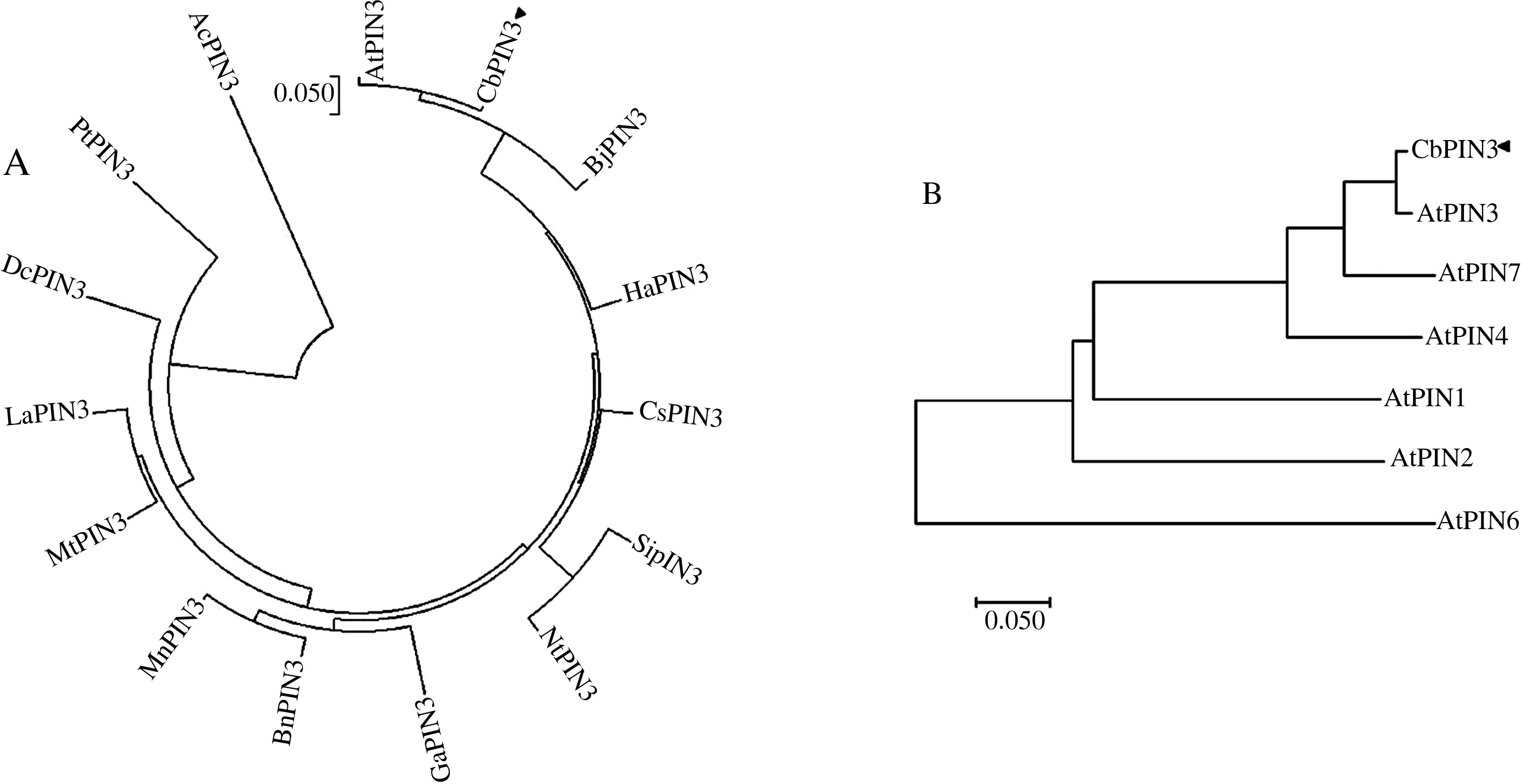
A.CbPIN3蛋白进化树分析;B.CbPIN3蛋白与拟南芥PIN蛋白进化关系
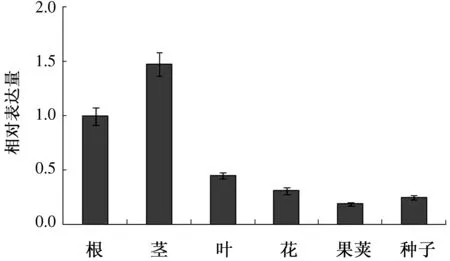
图11 CbPIN3组织表达分析
3 结论与讨论
作为植物的一种关键激素,生长素几乎参与调控植物生长发育的各个方面,包括植物对光和重力的向性反应、主根和侧根的发育、顶端优势的维持、器官的建构、维管组织的发育以及组织培养中的细胞生长等[16-18]。长久以来,由于无法直接显示生长素在植物体内的分布情况,因此影响了研究的进展。随着分子生物学技术的进步,生长素诱导基因的发现是生长素研究史上一大重要突破,生长素早期响应基因的启动子区域包含多个生长素反应元件,这些顺式作用元件赋予其生长素响应能力。通过对大豆GH3基因生长素反应元件改造,人工构建了生长素反应元件DR5[19-20],研究表明,DR5比天然的生长素反应元件具有更强的生长素响应能力,可提高5~10倍。DR5可调控报告基因GUS与GFP的表达,因此生长素报告系统DR5∶∶GUS、DR5∶∶GFP可用来在时间和空间上监控植物体内生长素的分布。DR5∶∶GUS/GFP已广泛用于研究拟南芥、烟草、小立碗藓、棉花、豌豆等多种植物生长发育过程,取得了非常显著的效果[21-24]。
通过对转基因荠菜花器官GFP荧光观察,发现在荠菜花瓣中荧光信号较低,表明存在着低水平的生长素;雄蕊中具有较强的荧光信号,表明生长素在花粉囊中大量合成;柱头上存在高强度荧光信号,推测雄蕊的伸长将生长素富集到花粉粒中,当花粉授到柱头上时使柱头维持高浓度的生长素水平;胚珠荧光信号较弱,表明生长素水平较低。这些结果均与拟南芥花器官发育中生长素分布情况类似,暗示荠菜和拟南芥花器官发育过程中生长素的分布有着较大的同源性。但二者也有不同:荠菜早期心皮上端两侧荧光信号较中间部分与下部强,而拟南芥的心皮中除柱头外信号处于均匀分布。由此推测,在荠菜心皮的形态建成中,存在生长素向两侧上端横向运输的机制,导致生长素的极性分布,加快细胞横向分裂,最终发育成心形心皮形态。
生长素发育调控作用的发挥需要生长素的极性分布。PIN3作为PIN家族的一员,调控生长素的横向运输,参与拟南芥的向光、重力等向性反应。本研究推测在荠菜的心皮发育中也存在生长素的横向运输,是否CbPIN3也参与了这一过程还未知,因此首先克隆了CbPIN3基因,分析了其蛋白结构特征。结果发现,CbPIN3与拟南芥PIN3高度同源,具有类似的磷酸化位点、跨膜结构域等。为进一步分析PIN3在荠菜生长发育中的作用,qRT-PCR分析了PIN3基因在荠菜不同组织的表达特征。结果发现,CbPIN3基因在荠菜多种组织中均表达,但表达量具有组织差异性。其中,茎中表达量最高,果荚中表达量最低,说明CbPIN3可能参与调控荠菜茎、根、叶、花、果实等多种器官的发育过程。
本研究利用生长素报告系统初步证实生长素参与了荠菜心形心皮的发育过程,其调控模式与Aloni等[25]、Sundberg等[5]利用DR5∶∶GUS/GFP系统研究拟南芥花器官发育过程生长素的分布结果较为一致,表明了荠菜与拟南芥心皮发育中生长素的极性分布差异导致了二者心皮形态迥异。此外,克隆了CbPIN3基因,发现PIN3蛋白与拟南芥PIN3蛋白高度同源,暗示其具备生长素横向运输能力。
