空心莲子草性可塑性影响下的克隆整合
2018-09-10全慧敏李文琪姜楚欧阳学军邓汝芳刘卫
全慧敏 李文琪 姜楚 欧阳学军 邓汝芳 刘卫


摘要: 植物的生活史由其有性生殖構件和营养体构件相互作用共同完成,克隆整合作为克隆植物的重要特征,其与有性生殖特征的相互作用关系却所知很少。该研究通过同质园种植实验,分析了空心莲子草的分株表型、生理、性别等与克隆整合的关系。结果表明:(1)克隆整合以及分株间是否连接对空心莲子草的表型特征、气体交换等生理性状和性别特征均有显著影响。(2)克隆整合显著缩小了雌雄同花和雄蕊心皮化两种性别植株间表型特征的差距,后代的性别特征与营养体表型特征显著相关。(3)在贫瘠的沙土基质中克隆整合明显增加了空心莲子草的营养体生长特征和气体交换等光合生理指标,但这种增加在富含有机质的塘泥基质中不明显。(4)居于不同土壤基质分株间的联系会减少分株表型特征和气体交换对生长环境的响应,并保持母体性别特征不受环境的影响,但单独居于沙土或塘泥单一土壤基质的分株性别特征却因受到环境影响而改变。因此,克隆整合有利于空心莲子草性别特征的稳定。
关键词: 克隆整合, 性别特征, 可塑性, 异质环境
中图分类号: Q943.2文献标识码: A文章编号: 10003142(2018)03030610
广西植物38卷3期全慧敏等: 空心莲子草性可塑性影响下的克隆整合收稿日期: 2017-06-14
基金项目: 国家自然科学基金(31370437) [Supported by the National Natural Science Foundation of China(31370437) ]。
作者简介: 全慧敏(1992-),女,湖南永州市人,硕士,主要从事分子生态学研究,(Email)quan_huimin@163.com。
*通信作者: 刘卫,博士,副研究员,主要从事分子生态学研究,(Email)liuwei@scbg.ac.cn。Clonal integration of Alternanthera philoxeroides
underlying effects of gender plasticity
QUAN Huimin1, 2, Li Wenqi1, 2, JIANG Chu1, 2, OUYANG Xuejun1, 3,
DENG Rufang1, LIU Wei1*
( 1. Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences,
Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Dinghushan Arboretum,
South China Botanical Garden, Chinese Academy of Sciences, Zhaoqing 526070, Guangdong, China )
Abstract: Life history of plant is accomplished by sexual reproductive components and vegetative growth components together. Clonal integration is an important characteristic of clonal plant, while the interactive relationship between clonal integration and sexual reproductive characteristic is known very little. In this paper, the interactive relationship between clonal integration and phenotypes, physiological traits and gender of Alternanthera philoxeroides were studied by homogeneous garden cultivation experiment. The results were as follows : (1) Both clonal integration and connection or disconnection had significant effects on the phenotypic characteristics, gender phenotypic traits, gas exchange properties and other physiological characteristics of A. philoxeroides. (2) Clonal integration significantly reduced the phenotypic characteristics of hermaphroditic plant and carpelloid plant of two sexes, and the gender characteristics of offsprings were apparently correlated with vegetative phenotype. (3) Clonal integration signicantly increased growth and gas exchange in barren sandy substrates, but did not show significant effect on both of these characteristics in abundant organic bog substrates. (4) Connection between ramets reduced both gas exchange and phenotypic responses to local conditions. Connection between ramets reduced both gas exchange and phenotypic responses to local conditions in different soil matrixes. At the same time, ramet gender was consistent with mother plant regardless of environmental conditions during clonal integration, while ramet gender was affected by environment in unconnected condition that cultivated in sand or bog alone. Therefore, clonal integration facilitates the stability of gender trait of A. philoxeroides.
Key words: clonal integration, gender characteristic, plasticity, heterogeneous environment
克隆植物(clonal plant)具有克隆生长的习性,自然条件下克隆植物以营养生长的方式产生克隆分株,这些分株具有潜在的独立性且遗传一致。植物的克隆分株相互连接,整合成由根状茎或匍匐茎连接的植物克隆(或基株)。植物的生长受各种环境资源的制约,当地上资源(如阳光)和地下资源(如土壤养分)能平衡时才能使生长达到较好的状态。然而,不同空间和时间的环境资源分布往往出现相反的状态。例如,一个斑块内光照充足而土壤水分贫乏,与之相邻的斑块内光照不足而土壤水分丰富 (董鸣等,2011)。克隆植物由于固着生长的习性,发展出表型可塑性以及克隆整合的能力来更好地应对自然资源的斑块性分布,提高适应性。克隆整合的优势在于克隆分株通过连接整合一个植物克隆或基株可以实现环境资源的共享,从而最大程度地利用斑块性分布的养分资源(Alpert,1996; 陈劲松等,2004)。这种资源利用方式,能够使克隆植物在许多生态系统中成功定居并迅速成为优势物种(de Kroon & Groenendael,1997)。
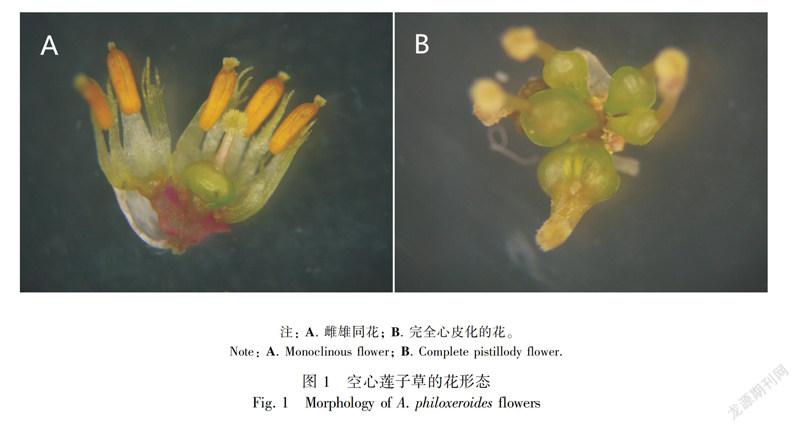
近年来,对克隆植物的研究多集中在克隆整合对营养体的影响和克隆植物对环境资源异质性的适应及其在生长发育、生理变化和繁殖方式等方面表现出的一些特殊性。克隆植物的特异性有表型可塑性(Hutchings & de Kroon,1994)、觅养特性(Oborny,1994)、风险分摊效应(Li & Takahashi,2003)、生理整合效应(Chen et al,2010)、劳动分工(Sampaio et al,2004)等。在资源等级分配的作用下,植物的完整生活周期在生殖构件和营养构件之间的相互影响下共同完成。Matsushita & Tomaru (2012)对雌雄异株植物Lindera triloba克隆分株的调查研究表明不同性别的克隆整合对无性分株的动力学起着不同作用,雄性系更为敏感。张玉芬和张大勇(2006)的研究表明克隆植物的有性繁殖特征在克隆整合的作用下明显发生变化,克隆分株的大小会影响有性繁殖和无性克隆间的资源分配策略。因此,克隆整合与植物性特征变化的关系应是整合效应研究中的重要方面,但目前这方面的研究明显不足。雄蕊心皮化指两性花中本应当发育为雄蕊的部位受环境条件的影响转变为雌蕊,从而表现为雄蕊消失的现象,这一雌化现象是植物以直接进化方式从雌雄同花向雌雄异花进化的途径之一(Liu et al,2011)。克隆植物空心莲子草的花器官受环境条件的影响表现为雌雄同花和心皮化两种可塑性状态(图1),并在自然条件下发育为只开雌性花和只开两性花的两种植株类型(Liu et al,2011)。空心莲子草花器官的这些特性,为我们研究克隆整合作用下有性的生殖构件和无性的营养构件之间的影响提供了理想的研究材料。本研究以空心莲子草为材料,尝试回答以下问题:异质环境下性别特征是否受克隆整合的影响以及克隆整合对营养体表型和部分生理特征的影响?
1材料与方法
1.1 研究地点与材料
在华南植物园的生态实验温室中进行,水泥池 1 m宽,铺有0.6 m深的基质,基质组成按实验设计来确定。沿水泥池长轴方向每间隔1 m插一木板,将水泥池分成数个相连的1 m2的单独小池用于不同实验处理。水生环境池铺有防水塑料,土壤相对持水度通过土壤水分测定仪[moisture probe meter (MPM160, ICT international)]测定调节。心皮化和雌雄同花的空心莲子草分株于自然环境中随机收集得来,洗净后每个分株选取两个外形一致的分枝茎段(顶芽下9个茎节约60 cm)作为繁殖体,材料以性别分为雌株(♀)和雌雄同花(♀)植株。将茎段两端的节放入1/2的Hongland培养液中培养10 d,待其生出须根后用于试验。注: A. 雌雄同花; B. 完全心皮化的花。
Note: A. Monoclinous flower; B. Complete pistillody flower.
1.2 试验设计
环境因素组合为陆生土培/水生沙培,水生沙培基质为1∶4的塘泥与沙混合,水漫过沙层5~10 cm,陆生土培基质为粉碎的塘泥。影响因素有性别和连接与否,交叉成4种处理,材料如图2所示在每两个不同生境水泥池的交界处放置一组4种处理,因两种生境间隔相连,每个池(1 m2)有8个繁殖体材料的各1个分枝平均分布在池子的两侧,每侧4个分枝中2个通过茎段与旁边另一生境中来自同一繁殖体材料的分枝相连,另2个在水泥池交界处与与旁边另一生境中来自同一繁殖体材料断开,每个处理10个重复,材料放置时未考虑茎段方向,为随机放置(图2)。栽培初期每天浇水,待3~4 d植株都恢复后,对陆生环境3 d浇一次水,用电导率仪测定控制土壤持水度为20%左右。
为保证所发现的处理间差异不是由于源株植物的遗传差异所致,来自同一分株(主茎长超过1 m)的两个茎段(分别长0.6 m)分别用于连接和截断处理。试验从2015 年11 月10 日至2016 年4 月10 日结束。于采收前一周测量气体交换数据。
1.3 数据测量
参照前面的研究,我们确定了两类需测量的性状。(1)表型性状,包括性别、平均叶面积(主茎上的第5到第9节上的叶片)、叶形指数(叶长与叶幅之比)、叶片数、分枝强度(分枝数/主茎长)、分枝长、分株产量、生物量、花序所含花数量、开花所需天数。(2)在收获的前两天进行气体交换测定,包括净光合速率(Pn,μmol·m2·s1)、蒸腾速率(Tr,mol·m2·s1)、氣孔导度(Gs,mol·m2·s1)、胞间二氧化碳浓度(Ci,μmol·mol1),气体交换参数测定采用Li6400 (LICOR Inc,Lincoln,NE,USA)测定。
测量各个指标,集中整理多变量。所有植物材料在70 ℃下烘48 h,并用电子天平称重,精确到0.1 mg。
1.4 数据分析
将各测定数据以株为重复单元,先以连接与否为控制变量对各类表型指标进行多元单因子方差分析(MANOVA);然后用ANOVA对性别、连接与否和环境条件以及它们的相互作用对各类表型指标是否有显著作用进行方差分析。所有的统计分析均用软件SPSS10.0完成,图用Sigmaplot 12.5软件进行制作。
2结果与分析
通过MANOVA进行方差分析,Pillai 检验得到F=41.37(P<0.01),表明不同处理中材料来源(包括重复材料的细微差异、茎段放置方向的差异等)所导致的结果差异不是试验结果的主要差异来源,因此对不同指标进行单独分析。
2.1 克隆整合对表型可塑性的影响
对心皮化的和雌雄同花的空心莲子草在不同生境下7个营养体特征和3个有性繁殖特征进行比较,结果表明无论不同生境分株之间是否连接,环境条件都显著影响两种性别植株的表型特征(图3,图4)。两种环境条件下,心皮化个体在有机质丰富的土培生境中空心莲子草的总分枝长、生物量、叶片数、分枝强度、分株数和平均叶面积分别是同样性别个体在沙土环境下的11.27、6.94、6.18、3.98、6.48和1.6倍。雌雄同花个体在有机质丰富的生境中的这些营养体特征分别是沙土环境下个体的9.61、5.85、5.52、4.02、5.1和1.87倍。由以上结果可知,在异质环境下不同斑块空心莲子草各组成器官变化显著,即空心莲子草分株之间表现出强的形态可塑性。环境对植株构型的影响建立在分株的基础上。
图3和图4结果表明,克隆整合明显地缩小了两个性别系统之间表型特征的差距。在含有丰富有机质的土培生境中,当克隆分株与其生长在贫瘠水生条件下的克隆分株相连时,心皮化个体的总分枝长、生物量、叶片数、分枝强度、叶形指数、分株数和平均叶面积分别是雌雄同花个体的1.05、1.09、1.09、0.96、1.06、1.04和1.0倍。相同生境条件下当克隆分株与其生长在贫瘠水生条件下的克隆分株断开时,心皮化个体与雌雄同花个体之间这些表型特征的比值为1.16、1.19、1.22、0.92、1.06、1.08和0.90。在贫瘠水生条件下,当克隆分株与其生长在含有丰富有机质生境中的克隆分株相连时,心皮化个体的总分枝长、生物量、叶片数、分枝强度、叶形指数、分株数和平均叶面积分别是雌雄同花个体的0.99、1.01、1.12、0.97、1.04、0.84和1.12倍,相同生境条件下当克隆分株与其生长在含有丰富有机质生境中的克隆分株断开时,心皮化个体与雌雄同花个体之间这些表型特征的比值为0.85、0.86、0.92、0.93、1.3、0.83和1.12。
由于克隆整合明显减少了两个性别系统之间表型特征的差距,因此,整体分析结果显示克隆分株是否与其他生境中的克隆分株相连对其表型无显著影响(图3)。但是,单独分析贫瘠生境或含有丰富有机质生境中克隆连接的影响表明,异质环境之间克隆分株的连接对空心莲子草的影响程度在这两种生境条件下有显著的不同。当贫瘠水生条件生长的空心莲子草分株与其生长在有机质丰富的环境下的克隆分株相连时,其平均叶面积、叶片数、总分枝长、分株数和生物量受到显著影响。在含有丰富有机质生境中生长的空心莲子草分株与其生长在贫瘠水生条件下的克隆分株相连时,只有叶片数受到显著的影响,但所受影响的显著程度低于在贫瘠水生条件生长的克隆分株(表1)。
图3结果表明,空心莲子草后代的性别特征与其营养体表型特征有显著的相关性。值得关注的是在不同的生长环境下这种相关性表现有明显的不同,在贫瘠水生条件下这种相关性只表现在后代性别与分株数(表1)。在含有丰富有机质生境中,这种后代性别与其他表型性状之间的相关性表现在叶片数、分枝强度、总分枝长、生物量上(表1)。
2.2 克隆整合对性系统特征的影响
克隆整合对空心莲子草的有性生殖特征有显著影响,在所测量的三个有性特征中受整合影响最显著的是性别转换频率(移栽后花性别发生可塑性变化的个体数/移栽个体总数),在两种环境中与异质环境相连的克隆株均未发生性别变化,但单独生长在贫瘠水生条件下的雌性个体和单独生长在有机质丰富的环境下的雌雄同花个体的性别转换频率分别为40%和20%。整合总体上对开花所需天数以及花序所含花的数量之间影响不显著,但在不同的生境下表现不同,在有机质丰富的环境下整合作用不明显(表1)。在贫瘠水生条件下异质环境之间克隆植株的连接显著影响这两个有性生殖特征(表1),连接的植株比不连接的植株平均早开花9 d并且花序中多4.8朵花,另外后代的性别在两种环境条件下都显著影响这两个有性特征。
2.3 克隆整合对光合特性的影响
异质生境的克隆片段连接与否显著影响空心莲子草的净光合速率和气体交换特征,无论是在有机质丰富的土培条件下还是在贫瘠水生条件下,与异质生境连接的克隆植株的各项光合参数都有所提高。在前一种生境条件下与异质生境连接的克隆植株的净光合速率、胞间CO2浓度、气孔导度、蒸腾速率、水分利用效率分别是同一生境下截断组的1.1、1.03、1.21、1.07、1.03倍。在后一种生境条件下,这些比值变为1.59、1.09、2.25、1.51、1.05。这表明除了水分利用效率外,不同生境之间克隆分株的连接对水生贫瘠条件下的分株的光合生理特征的提高更为显著。
不同生境条件下个体的性别对光合特性的影响也有所不同,在贫瘠水生条件下性别间的差异表现在蒸腾速率和水分利用效率上,另外连接状态和性別特征的交互作用对净光合速率、气孔导度以及水分利用效率有显著影响。在有机质丰富的条件下,性别间的差异表现在胞间CO2浓度和水分利用效率上,而连接状态和性别特征的交互作用对所测的光合生理指标没有影响(表2)。
3讨论与结论
本研究结果表明,空心莲子草除了可通过对克隆构型表型的改变(如在水,陆两种异质环境条件下分枝强度的不同)来适应环境外,生理的适应性也是其适应可塑性的重要组成部分。
影响克隆生长和克隆繁殖的因子很多,如种内竞争和环境异质性。自然环境中,资源分布在时间和空间上的异质性普遍存在。因此,克隆植物对环境的反应还可能表现在分株种群的特征变化上。克隆植物同一基株内各分株相互连接,除分株个体会对特定环境产生变化外,特定环境中的分株还可能通过匍匐茎或根状茎等连接物引起其它分株的变化,导致基株水平上克隆构型发生改变,并进一步引起种群特征变化(Hutchings & de Kroon,1994)。克隆分株之间间隔距离的长度决定了克隆植物不同分株间的资源共享主要发生在较小的尺度上。因此,小尺度上的分布格局更能反映克隆共享对克隆分株分布格局的影响。董鸣等(1999)、于飞海和董鸣(1999)、陈玉福和董鸣(2000)、廖明隽等(2002)分别对中国内陆沙化生境中的根茎禾草沙鞭、根茎草本披针叶黄华、根茎灌木羊柴以及内蒙古锡林河流域的羊草的分株种群特征进行了研究,他们的研究结果表明克隆植物的分株种群特征能用于反映和指示克隆植物的生态适应对策。因此,克隆构型整合和分株种群特征是植物克隆生长及其生态适应对策研究的基本内容(Hutchings & de Kroon,1994)。
本研究中,与资源的空间同质性条件相比,当空心莲子草生长在资源异质性条件下,其克隆片段的生长指标如生物量和分株数显著增加,这一结果暗示相互连接的分株之间可能发生了资源的交互传输,而且这种交互传输促进了分株和克隆片段的生长。这些相连的克隆分株和克隆片段间存在以源-汇关系为驱动力的养分、水分以及光合产物等的传递。通过物质之间的这种传递,分布在不同环境斑块中的克隆分株彼此间可以相互传输养分等资源,从而缓解了局部资源的匮乏,促进克隆植物的生长(Cook,1985; de Kroon & Groenendael,1997;李德志和王长爱,2004)。de Kroon et al(1996)对根茎苔草(Carex hirta)的研究发现,当生注: (A) Pn. 净光合速率; (B) Ci. 胞间二氧化碳浓度; (C) Days. 开花所需天数; (D) Gs. 气孔导度; (E) FT. 花序所含花数量;
(F) Tr. 蒸腾速率; (G) SRO. 性别转换频率; (H) WUE. 水分利用率。x轴1. 塘泥连接; 2. 塘泥截断; 3. 沙土连接; 4. 沙土截断。
Note: (A) Pn. Net photosynthetic rate; (B) Ci. Intercellular CO2 concentration; (C) Days. Days elapsed before flowering; (D) Gs. Stomatal
conductance; (E) FT. Total flowers per fullblown inflorescence; (F) Tr. Transpiration rate; (G) SRO. Sexual rate of offsprings;
(H) WUE. Water use efficiency. x axis1. Bog connection; 2. Bog disconnection; 3. Sand connection; 4. Sand disconnection.
图 4雌雄同花和心皮化空心莲子草在同质和异质环境下有性特征和气体交换参数的平均值
Fig. 4Mean values of reproductive traits and gas exchange parameters of monoclinous and pistillody
Alternanthera philoxeroides under homogeneous and heterogeneous environmental conditions
长于水分供应条件良好的分株与水分供应条件差的分株相连时,生长于水分供应条件良好的分株根的水分速率有显著提高。
本研究中,环境条件明显地决定了植株从整合中获得利益的程度。当克隆整合发生时,生长于水生贫瘠条件下植株增长的程度远高于生长于富营养条件下的植株,特别是在富营养条件下的植株对整合的反应并不十分显著,即使在异质环境中相互连接的两部分植株之间表型和生物量上的差距也非常明显,这表明空心莲子草的克隆分株应对环境变化时首先将营养等用于自身的生长。因此,生物量分配格局对资源水平的可塑性反应是实现分株功能特化的基本途径之一,分株获取地上和地下资源功能的特化可以通过调整对地上和地下资源吸收器官的生物量相对投资比例来实现。伴随着养分和光合产物等物质的相互传递,空心莲子草相连分株通过生物量分配的可塑性实现了对局部丰富资源吸收的功能特化。克隆整合虽能够缓解局部资源胁迫对克隆分株生长的影响,但它可能只能在一定程度上缓解资源异质性对空心莲子草生长的影响。性别因素以及性别与环境因素的交互综合作用对植株的部分表型特征有显著的影响,但这种影响在不同的环境条件下表现有所不同,这其中的意义尚需进一步的分析。
克隆整合显著增强了空心莲子草性别特征的稳定性,本研究中处于两种环境中与异质环境相连的克隆株均未发生性别变化,而相连接的各分株间在表型和气体交换等特征上还是表现出由环境造成的明显不同,这一性系统和营养体对克隆整合反应的不同至今未见其它报道,是否意味着控制性别信号的物质和水光和产物等在克隆分株间的传输效率不同尚需其它研究验证,如同位素標记物在分株间运输等实验(Saitoh et al,2006),但从繁殖生态学角度来说,这种性系统信号传输的高效性能够保证一个斑块内各个分株间生殖周期一致。另外,本研究中空心莲子草母体的性别特征对实验材料性别的影响也值得考虑,这种影响与其它心皮化现象研究中认为其母体性别特征具有传代性的结论相似(Murai et al,2002; Mitchell & Diggle,2005; Liu et al,2011)。现有对心皮化现象的分子生物学研究表明,心皮化与染色体上某些区域的染色质结构的异质化有关(Iltis,1983; Olimpieri & Mazzucato,2008)。例如,木瓜Y染色体上雄性区域的特异性造成了花结构的心皮化(Zhang et al,2008)。异质小麦杂交所导致的核质互作与花的心皮化可塑性有关(Murai et al,2002)。类似于异质小麦的杂交来源,分布于我国的空心莲子草被证明为异源六倍体,与其来源地的杂合基因型相近(Sosa et al,2008; 蔡华等,2009)。另外,虽然在野外空心莲子草不发生有性生殖,但在第二年春季地下肉质根萌生新分株时依然要经历一个单细胞的起始过程,此时低温可以起到类似有性生殖的作用——诱导细胞中核小体发生染色质释放和去甲基化(Steward et al,2002)。因此,环境因素作用于性别转换及光合特性等其它表型可塑性的一致性表明,环境决定空心莲子草的性别转换可塑性中双稳态的表现形式。
参考文献:
ALPERT P, 1996. Nutrient sharing in natural clonal fragments of Fragaria chiloensis [M]. J Ecol, 84: 395-406.
CAI H, WEI CL, CHEN N, 2009. Chromosome karyotype characters of biological invasion in Alternanthera philoxe roides [J]. Chin J Trop Crops, 30(4): 530-534. [蔡华, 韦朝领, 陈妮, 2009. 生物入侵种喜旱莲子草的染色体核型特征 [J]. 热带作物学报, 30(4): 530-534.]
CHEN JS, DONG M, YU D, et al, 2004. Intraclonal spatial division of labour in two stoloniferous plants with different branching type in response to reciprocal patchiness of resources [J]. Acta Ecol Sin, 24(5): 920-924. [陈劲松, 董 鸣, 于 丹, 等, 2004. 资源交互斑块性生境中两种不同分枝型匍匐茎植物的克隆内分工 [J]. 生态学报, 24(5): 920-924.]
CHEN JS, LEI NF, DONG M, 2010. Clonal integration improves the tolerance of Carex praeclara to sand burial by compensatory response [J]. Acta Oecol, 36: 23-28.
CHEN YF, DONG M, 2000. Genet characters of Hedysarum leave and the characters of its ramet population in different habitats in Mu Us sandland [J]. Chin J Plant Ecol, 24(1): 40-45. [陈玉福,董鸣, 2000. 毛乌素沙地根茎灌木羊柴的基株特征和不同生境中的分株种群特征 [J]. 植物生态学报, 24(1): 40-45.]
COOK RE, 1985. Growth and development in clonal population [M]// JACKSON JBC, BUSS LW, COOK RE. Population biology and evolution of clonal organisms. New Haven: Yale University Press: 259-296.
DE KROON H, FRANSEN B, VAN RHEENEN JWA, et al, 1996. High levels of interramet translocation in two rhizomatous Carex species, as quantified by deuterium labeling [J]. Oecologia, 1: 730-84.
DE KROON H, GROENENDAEL J, 1997. The ecology and evolution of clonal plants [M]. Leiden: Backhuys Publishers: 263-289.
DONG M, ALATENG B, XING XR, 1999. Genet features and ramet population features in the rhizomatous grass species Psammochloa villosa [J]. Chin J Plant Ecol, 23(4): 302-310. [董鸣, 阿拉腾宝, 邢学荣, 1999. 根茎禾草沙鞭的克隆基株及分株种群特征 [J]. 植物生态学报, 23 (4): 302-310.]
DONG M, YU FH, CHEN JS, 2011. Clonal plant ecology [M]. Beijing: Science Press: 36-39. [董鸣, 于飞海, 陈劲松, 2011. 克隆植物生态学 [M]. 北京: 科学出版社: 36-39.]
HUTCHINGS MJ, DE KROON H, 1994. Foraging in plants: the role of morphological plasticity in resource quisition [J]. Advan Ecol , 25: 159-238.
ILTIS H, 1983. From Teosinte to maize: The catastrophic sexual transmutation [J]. Science, 222: 886-895.
LIAO MX, WANG QB, SONG MH, 2002. Clonal architecture and ramet population characteristics of Leymus chinensis from different habitats in the Xilin River watershed [J]. Chin J Plant Ecol, 26 (1): 33-38. [廖明雋, 王其兵, 宋明华,2002. 内蒙古锡林河流域不同生境中羊草的克隆构型和分株种群特征 [J]. 植物生态学报, 26 (1): 33-38.]
LI DZ, WANG CA, 2004. Extraordinary the ecology of clonal plants——review on The Ecology and Evolution of Clonal Plants [J]. Chin J Ecol, 23(1): 139. [李德志, 王长爱, 2004. 别开洞天的克隆植物生态学——评《克隆植物的生态学与进化》 [J]. 生态学杂志, 23(1): 139.]
LI DZ, TAKAHASHI S, 2003. Particularities of clonal plant species induced by physiological integration [J]. Grassl Sci, 49(4): 395-402.
LIU W, DENG RF, LIU WP, et al, 2011. Phenotypic differentiation is associated with gender plasticity and its responsive delay to environmental changes in Alternanthera philoxeroidesphenotypic differentiation in alligator weed [J]. PLoS ONE, 6(11): e27238.
MATSUSHITA M, TOMARU N, 2012. Differences in clonal integration between the sexes: longterm demographic patterns in the dioecious, multistemmed shrub Lindera triloba [J]. Botany, 90(10): 1028-1035.
MITCHELL CH,DIGGLE PK, 2005. Evolution of unisexual flowers: morphological and functional convergence results from diverse developmental transitions [J]. Am J Bot, 92(7): 1068-1076.
MURAI K, TAKUMI S, KOGA H, et al, 2002. Pistillody, homeotic transformation of stamens into pistillike structures, caused by nuclearcytoplasm interaction in wheat [J]. Plant J, 29: 169-181.
OBORNY B, 1994. Growth rules in clonal plants and environmental predictability—a simulation study [J]. J Ecol, 82:341-351.
OLIMPIERI I, MAZZUCATO A, 2008. Phenotypic and genetic characterization of the pistillate mutation in tomato [J]. Theor Appl Genet, 118(1): 151-163.
SAITOH T, SEIWA K, NISHIWAKI, 2006. A. Effects of resource heterogeneity on nitrogen translocation within clonal fragments of Sasa palmata:an isotopic (15N) assessment [J]. Ann Bot, 98(3): 657-663.
SAMPAIO MC, ARAUJO TF, SCARANO FR, et al, 2004. Directional growth of a clonal bromeliad species in response to spatial habitat heterogeneity [J]. Evol Ecol, 18: 429-442.
SOSA AJ, TELESNICKI M, CORDO HA, 2008. Alligator weed. in: south American biological control laboratory [J]. Ann Rep: 15-18.
STEWARD N, ITO M, YAMAKUCHI Y, et al, 2002. Periodic DNA methylation in maize nucleosomes and demethylation by environmental stress [J]. J Biol Chem, 277: 37741-37746.
YU FH, DONG M, 1999. Multi_scale distribution pattern of natural ametpopulation in the rhizomatous herb, Thermopsis lanceolata [J]. J Integr Plant Biol, 41(12): 1332-1338. [于飛海, 董鸣, 1999. 根茎草本披针叶黄华自然分株种群多尺度分布格局 [J]. 植物学报, 41(12): 1332-1338.
ZHANG YF, ZHANG DY, 2006. Asexual and sexual reproductive strategies in clonal plants [J]. Chin J Plant Ecol, (1):174-183. [张玉芬, 张大勇, 2006. 克隆植物的无性与有性繁殖对策 [J]. 植物生态学报, (1):174-183.]
ZHANG WL, WANG X, YU QY, et al, 2008. DNA methylation and heterochromatinization in the malespecific region of the primitive Y Chromosome of papaya [J]. Geno Res, 18(12): 1938-1943.
