circRNA研究现状及展望
2018-09-10王宏浩韩文静张燕军苏蕊刘志红王瑞军王志英赵艳红王志新李金泉
王宏浩 韩文静 张燕军 苏蕊 刘志红 王瑞军 王志英 赵艳红 王志新 李金泉
摘要:circRNA(Circular RNAs)是一类不具有5末端帽子和3末端poly(A)尾巴并以共价键形成封闭环状结构的内源性非编码RNA(ncRNA)分子,根据circRNA的剪接来源不同,可分为外显子circRNA(Exonic cirRNAs)、内含子circRNA(cirRNAs)和外显子一内含子circRNA(Exon-in-troncircRNA,ElciRNA)。文章从分子特征、研究历程、形成过程、作用机制及其数据库及分析工具等方面对circRNA研究现状和生物学功能进行综述,结果显示随着高通量测序技术的进步和生物信息学的快速发展,越来越多的circRNA被挖掘,其具有稳定性、广泛性、保守性及组织特异性等性质,在基因表达调控中发挥重要作用,可作为一种疾病的生物标志物,在临床诊断和治疗方面具有广阔的应用前景。但circRNA还有许多尚未知悉的重要生物学功能,如其在绒毛生长发育中的作用机理,且在疾病诊断和治疗方面研究不够深入。在今后研究中,应深入研究circRNA及其在疾病诊断、治疗方面及绒毛发育的作用机制,充分了解其更多的生物学功能,使其广泛应用于各研究领域。
关键词:circRNA;特征;形成;生物学功能;作用机制
中图分类号:S818.9 文献标志码:A 文章编号:2095-1191(2018)03-0431-09
0引言
人类基因组中可编码蛋白质的DNA仅为20%,其余均称为非编码RNA(Non-coding RNAs,ncRNAs),不被翻译成蛋白质,被认为是基因组的“暗物质”(Pennisi,20 10),但并不表示其无遗传信息或生物学功能(Mattick and Makunin,2006)。研究表明,ncRNAs参与细胞增殖、分化、凋亡、代谢、衰老等重要的生物学过程(Blaxter,2010;李静秋等,2015),尤其是其参与基因转录后调控(Memczak et a1.,2013)。自1990年人类基因组计划被提出以来,ncRNAs逐渐被人们重视。根据ncRNAs结构的不同,可将其分为线形ncRNAs和环状ncRNAs(Circular RNAs,circ-RNA)(Capel et a1.,1993)。但人们把更多的注意力聚焦在线性ncRNAs,尤其微小RNA(microRNA)和长链非编码RNA(Long non-coding RNA,IncRNA)的相关研究(Wang and Chang,2011;Qi et a1.,2013;王建勋等,2016),而鲜见circRNA的相关研究,主要原因是相对于大量的经典线性ncRNAs,circRNA较罕见,曾被认为是错误剪接的副产物(Cocquerelle eta1.,1993;Benderdour et a1.,2002)。直到2015年An-dreeva和Cooper研究报道了circRNA广泛存在于动植物细胞组织中,且具有很多特殊的生物学特性之后,才引起国内外科学家的高度重视。目前circRNA还有许多尚未知悉的生物学功能,应加大研究力度,使其广泛应用于各领域。
1circRNA的分子特征
circRNA有别于线形ncRNAs,是一类不具备5末端帽子和3末端poly(A)尾巴并以共价键形成封闭环状结构的内源性ncRNA分子,具有以下特征:
(1)circRNA广泛存在于真核生物体的不同组织,如睾丸(Capel et a1.,1993)、大脑(Hansen et a1.,2011;Rybak-Wolf et a1.,2015;Hanan et a1.,2016)、胃(Chen et a1.,2016)、乳腺(Nair et a1.,2016)和前列腺等(Greene et a1.,2016),多数位于细胞质,少数位于细胞核(Zhang et a1.,2013)。此外,circRNA在人体某些组织中的表达水平超过其线性异构体的10倍(Julia et a1.,2012;Jeck et a1.,2013)。
(2)circRNA序列高度保守,但部分内含子构成的circRNA序列保守性不强(Jeck et a1.,2013)。
(3)circRNA的结构有别于线形ncRNAs,对核酸酶不敏感,因此较线形ncRNAs更稳定(Memczaket a1.,2013;肖时曦和王涛,2017)。
(4)大多数circRNA由蛋白质编码基因的外显子组成,少数由内含子或内含子片段直接环化形成(战昊等,2016)。
(5)circRNA具有microRNA应答元件(micro-RNA response element,MRE),可作为竞争性内源RNA(Competing endogenous RNA,ceRNA),与mi-croRNA结合,在细胞中发挥microRNA海绵效应(microRNA sponge),强烈抑制microRNA活性,导致靶基因表达水平增加(Hansen et a1.,2013a;玄丽佳和孙亚男,2016)。
(6)虽然circRNA不翻译成蛋白质,但可作为蛋白质的合成模板(Wang and Wang,2014),已有研究证实,大量circRNA可作为信使RNA(mRNA)来编码蛋白(Hsiao et a1.,2017;Yang et a1.,2017)。
(7)多数circRNA在转录后发挥调控作用,少数只能在转录水平发挥调控作用(Zhang et a1.,2013)。
2drcRNA的研究历程
1976年Sanger等在研究马铃薯纺锤块茎病中发现了可致病的单链环状类病毒。这是人类第一次发现circRNA。1979年Hsu和Cocaprados用电子显微镜观察到circRNA存在于真核细胞细胞质。1980年Amberg等在酵母线粒体中发现了circRNA。1993年Capel等在小鼠睾丸决定性别基因(Sex-determiningregion Y,Sry)中发现有circRNA转录。1993年Coc-querelle等在人体细胞中发现circRNA。201 1年Danan等研究发现,circRNA普遍存在于古生菌细胞中。Jeck等(2013)在人类成纤维细胞中检测出25000多种circRNA。2013年Memczak等结合人白細胞数据库,从RNA-Seq数据中鉴定出人类circRNA 1950种、小鼠circRNA 1903种(其中与人类circRNA相同的有81种)和线虫circRNA 724种。在今后研究中,科学家应借助高通量测序技术和生物信息学技术,对circ-RNA展开系统深入的研究。
3drl2RNA的形成过程
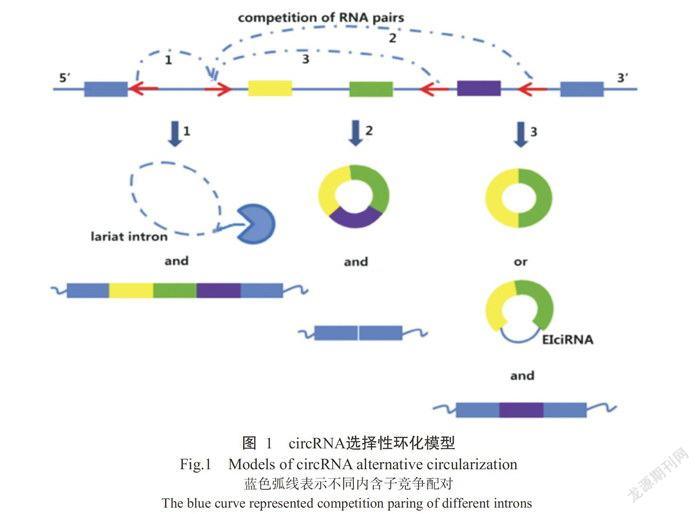
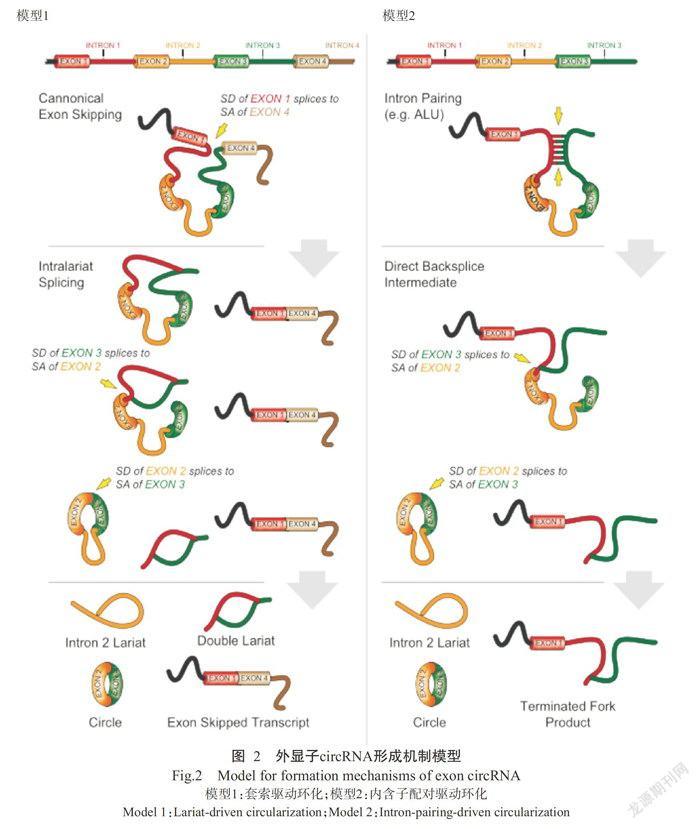
目前,人们普遍熟知从mRNA前体(pre-mRNA)到线形ncRNAs的转录过程,但事实上pre-mRNA还可被非线性地反向剪接成circRNA(Memczak et a1.,2013)。circRNA的形成取决于顺式作用元件(Cis-acting element)和反式作用因子(Trans-act factor)。circRNA选择性环化机制分为三类,其模型(Qu eta1.,2015)如图1所示。基因组同一位置可转录出mRNA和circRNA,通过内含子序列问竞争性互补配对形成外显子circRNA(Exonic cirRNAs)、内含子circRNA(cirRNAs)和外显子一内含子circRNA(EX-on-in-troncircRNA,EIciRNA)3类circRNA分子(Quet a1.,2015)。这些circRNA分子问存在竞争性平衡关系,可影响mRNA的表达。该选择性环化机制非常复杂,可能需要一些因子参与调控,如RBPs等(chen and Yang,2015)。其中外显子circRNA的形成机制又分为两种,其模型(Jeck et a1.,2013)如图2所示。模型1为套索驱动环化(Lariat-driven circula-rization),外显子跳跃形成套索结构促进环化,随后剪掉内含子形成circRNA(Jeck et a1.,2013)。模型2为内含子配对驱动环化(Intron-pairing-driven circu-larization),不需要外显子跳跃,而是通过两侧内含子上的反向互补序列碱基配对形成环状结构,随后剪掉内含子,最终形成circRNA(Jeck et a1.,2013;闫宁宁等,2015)。
3.1剪接体促进circRNA形成
circRNA是剪接体通过非经典剪接方式对pre-mRNA进行反向剪接而形成,其中剪接体主要是由蛋白质和小分子核RNA(snRNA)组成,是一个庞大、复杂的动态分子机器,在circRNA形成过程中发挥重要作用。外显子circRNA形成的两种模型不同之处在于第一步形成环状结构机制,即套索驱动的环化是由外显子组成的剪接供体和剪接受体共价结合形成环状结构,而内含子配对驱动的环化则是2个内含子互补配对结合形成环状结构(Jeck et a1.,2013)。但也有研究表明,circRNA表达并不总是与同源基因所产生的线形ncRNAs表达相关,说明剪接体能区分前向剪接(Forword splicing)(Jeck et a1.,2013)和后向剪接(Back splicing)(Starke et a1.,2015),但目前对剪接体区分机制尚不清楚。
3.2顺式作用元件促进circRNA形成
已有研究证实,不同顺式内含子互补配对序列的竞争性配对可导致circRNA可变反向,从而使一个基因位点产生多个circRNA,表明顺式元件一内含子互补配对序列对circRNA表达发挥重要作用(zhanget a1.,2014;Zhang et a1.,2016)。
3.3反式作用因子调控circRNA形成
3.3.1盲肌(Muscleblind,MBL)调控circRNA形成 盲肌是一种RNA结合蛋白(RNA-binding pro-tein,RBP)。pre-mRNA两侧的内含子上均有MBL结合位点,可特异性结合MBL(Ashwal-Fluss et a1.,2014)。这些结合位点的存在表明一些MBL亚型可能促进circRNA产生。因此,MBL是参与调控circ-RNA合成的生物因子(玄丽佳和孙亚男,2016)。
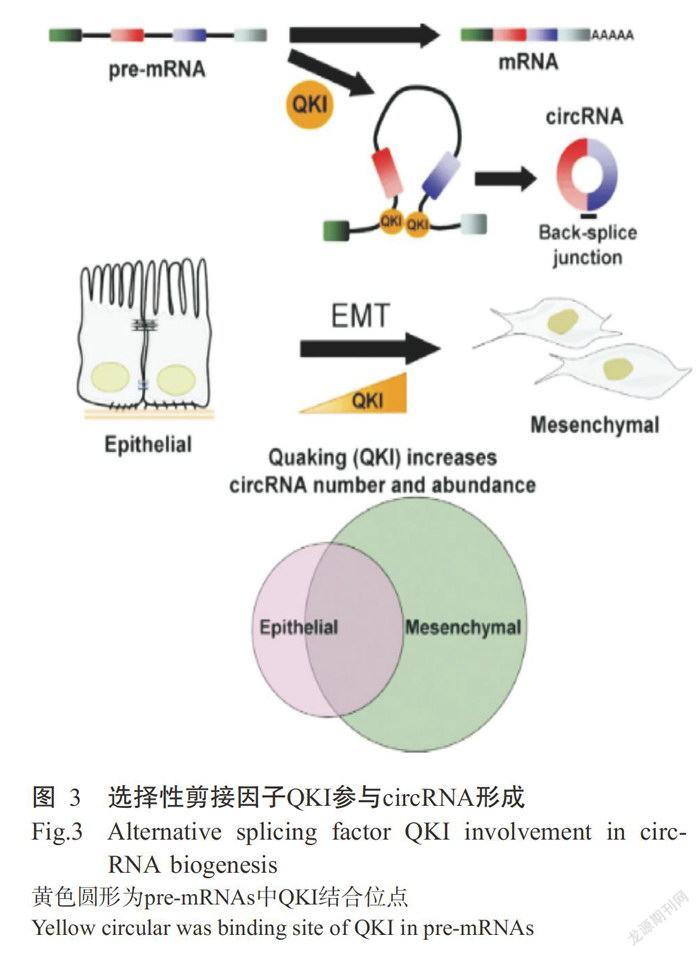
3.3.2选择性剪接因子QKI参与circRNA形成 前人研究表明,在人类上皮间质转化(Epithelial tomesenchymal transition,EMT)过程中超过33%的circ-RNA受选择性剪接因子Qrd的动态调节(Conn eta1.,2015),如图3所示。在EMT过程中,Qrd通过结合pre-mRNA上的特定结合位点,促进circRNA形成,Qrd表达量越高,circRNA数目越多。可见,上皮细胞中高效表达QKI会促进circRNA的形成。
3.3.3 RNA编辑修饰酶ADARl抑制circRNA形成 ADARl不仅能编辑修饰RNA,还可作为双链RNA结合蛋白。因此,ADARl可通过干扰RNA配对抑制circRNA形成,从而使circRNA数量减少(chen andYang,2015;chen,2016),表明cicrRNA形成与RNA编辑修饰酶ADARl表达呈负相关。
4circRNA作用机制
4.1circRNA发挥microRNA海绵效应
circRNA具有许多的生物学特性,但相关研究主要集中在其与microRNA调控机制上。前人研究表明,富含microRNA结合位点的circRNA是microRNA的海绵吸附体,可抑制microRNA与靶基因结合,从而发挥microRNA海绵效应,提高靶基因的表达水平,是一类高效率的ceRNA(李祯祺等,2015;Liu et a1.,2016)。目前,研究最多的是ciRS-7,其作为miR-7的海绵吸附体,通过募集miR-7间接调控多种癌相关信号转导通路中主要的致癌基因表达下调(李培飞等,2014;战昊等,2016),从而发挥抑癌作用。Jens(2013)研究发现,小脑退行性相关蛋白基因1(CDR,)的circRNA分子(cDRlas-ciRS-7)广泛存在于人和小鼠的大脑,包含73个保守的miR-7靶位點,从而抑制miR-7与靶基因CDRI的结合,发挥microRNA海绵效应。可见,当microRNA与对应的mRNA结合时可阻止蛋白质生成,而microRNA被circRNA吸附时,mRNA翻译合成蛋白质(肖时曦和王涛,2017)。
4.2 circRNA的生物合成过程影响基因剪接
已有研究证实,真核细胞circRNA是通过反向剪接pre-mRNA而形成(Chen,2016)。由此推测剪接pre-mRNA的方式有3种:第一种是通过经典剪接生成线性mRNA;第二种是反向剪接和经典剪接相结合生成circRNA;第三种是反向剪接、经典剪接和选择性剪接相结合对线性mRNA进行剪接(chen eta1.,2015)。Chen(2016)研究发现,虽然circRNA表达水平较低,但可通过不同途径影响基因表达,而有效扩展真核细胞转录组的多样性和复杂性。
4.3circRNA调节基因转录
circRNA可参与基因转录调控,如来源于锚蛋白重复结构域52(ANKRD52)基因二号内含子区域的circRNA(ciRNA derived from intron ofANKRD52,ci-ankrd52)能产生锚蛋白重复结构域,通过反义寡核苷酸(Antisense oligo deoxynucleotide,ASO)封闭ci-ankrd52的表达,致使ANKRD52 mRNA的表达量显著减少(Zhang et a1.,2013)。另外,含内含子序列的circRNA常位于细胞核内,发挥基因转录调控的功能,ci-ankrd52主要于细胞核中积累,可促进RNAPolII对ANKRD52基因的转录(chen,2016)。
4.4circRNA调控基因表达
ElciEIF3J是由真核翻译延长因子3J(EW3J)产生的一种含内含子的circRNA,与U1 snRNP和EIF3J启动子共同促进EIF3J转录(Li et a1.,2015b)。此外,ElciRNAs的发现暗示circRNA的种类及功能具有多样性,证实了circRNA在细胞核中调控基因表达的重要功能,并揭示了ElciRNA与U1 snRNA相互作用的调控机制(Li et a1.,2016)。
4.5circRNA抑制翻译
人抗原R(Human antigen R,HuR)是一种RNA结合蛋白,通过结合多种RNA调控蛋白表达模式(Brennan and Steitz,2001)。Abdelmohsen等(2017)从Hela细胞中分离出与HuR相互作用的circRNA,其中以hsa circ 0031288的变化最明显,与其对应是poly(A)结合蛋白核1[poly(A)-binding protein nu-clear 1,PABPN1]基因的mRNA前体(circPABPNl),其功能不受HuR影响,但大量的circPABPNl会竞争性结合HuR,阻止HuR结合至PABPNl的线性ncRNAs上,从而降低PABPN1 mRNA的翻译效率(图4)。
4.6circRNA为疾病的生物标志物
circRNA作为microRNA的海绵吸附体,可间接调控microRNA靶基因表达,在人类疾病的发生发展过程中发挥重要作用(Garzon et a1.,2009;Hansen eta1.,2013b;任显显等,2016)。因此,circRNA可作为疾病的生物标志物,广泛用于疾病诊断。Ahmed等(2016)对3例IIIc期病人的原发部位、腹膜和淋巴结转移灶进行了RNA双端测序(Paired-end RNA se-quencing),结果发现在高度异质性癌症转录组中,circRNA表达具有一致性,最适合作为卵巢癌的生物标志物,为该症治疗及预后提供新的候选标志。Floris等(2016)利用Arraystar Human circRNA探针(含有13617个人类circRNA)和qRT-PCR技术筛选差异表达的circRNA,并检测抗抑郁治疗4周和8周后的circRNA水平,结果发现仅1个circRNA(hsa circ103636)表达量在抑郁症患者治疗8周后显著下调,表明hsa circ 103636与抑郁症的疾病进程有关,可用作重度抑郁症患者诊断与治疗的潜在的新生物标志物(Cui et a1.,2016)。Shang等(2016)检测了肝癌变组织中3个circRNA(hsa circ 0000520、hsa circ0005075和hsa circ 0066444)的表达水平,结果发现hsa circ 0005075是一个潜在的肝癌标志物,但尚不清楚hsa circ 0005075及其他circRNA在肝癌发生发展中的作用机制。circRNA在心血管疾病和胃癌诊断中也发挥重要作用,如hsa circ 0124644可用于冠状动脉疾病的诊断(zhao et a1.,2017),hsa circ002059、hsa circ 0000096和hsa circ 0000190可作为胃癌诊断标志物(Li et a1.,2015a;Chen et a1.,2017;Li et a1.,2017)。此外,动脉粥样硬化(Burd eta1.,2010)、病毒性肝炎(Taylor,1991)等疾病均与circ-RNA有关。
5circRNA数据库及分析工具
随着高通量测序技术和生物信息学技术的不断发展,circRNA的研究越來越深入,不同种类的circRNA被发现,相关数据库和分析工具也相继推出。目前,广泛使用的数据库包括deepBase数据库(http://deepbase.sysu.edu.cn/)、microRNA-circRNA相互作用网络数据库circRNABase(http://starbase.sysu.edu.cn/mirCircRNA.php)(Li et a1.,2013;Li eta1.,2014)、circBase数据库(http://www,circbase.org/)、Circ2Traits功能数据库(http://gyanxet—beta.com/circdb/)、CircRNADb数据库(http://reprod.nimu.edu.cn/circmadb)等。其中,deepBase数据库包含大约人类、鼠等不同物种的15万个circRNA数据;circ-Base数据库包含大量已知或未知功能的circRNA数据,不但可准确地找到某个circRNA的成熟体序列,还能查询某基因能产生circRNA的个数及其表达丰度;CircRNADb数据库是首个汇总可编码蛋白的circ-RNA的数据库(Chen et a1.,2016)。HppRNA(https://sourceforge.net/projiects/hppma/)是一种新开发的RNA—Seq数据分析工具(Huang et a1.,2015;Wang,2017),包含pre—mapping、sequence variation detec-tion等模块,可满足大样本系统性分析的需求。可见,这些数据库和分析工具的推出为深入挖掘当前大数据提供了便利,但仍需通过试验对其预测分析结果进行验证。
6展望
circRNA在各领域的研究如雨后春笋。目前人们对circRNA的形成和特征已基本了解,但仍有许多生物学功能尚不清楚。有关circRNA的研究主要集中在其与疾病问的相关性方面,大量研究表明,circ-RNA可作为疾病的生物标志物,在癌症治疗及预后方面存在巨大的发展潜力,但对各种疾病的发生发展调控机制知之甚少,需进行深入探究。现已研究证实,在动物体内microRNA通过与靶mRNA的3端非翻译区结合抑制转录后的翻译,从而发挥调控基因表达的作用(Doench et a1.,2003)。付绍印等(2012)研究表明,microRNA靶基因指纹图谱(Mi—croRNA targets fingerprint,MTFP)经济适用、操作性强,可用于探索microRNA调节的靶基因或评估靶基因的表达谱特征。马继登(2014)研究发现,猪体内circRNA可能同样发挥着microRNA海绵吸附体的作用,间接调控猪的生理代谢。VenO等(2015)运用高通量测序技术从猪胚胎脑组织中鉴定出4634个circRNA,其在猪胚胎脑组织发育过程中发挥重要调节作用。随着高通量技术的发展,越来越多与绒毛生长发育相关的miRNA及其靶基因被发掘鉴定。刘洋(2016)对绒山羊胎儿期毛囊发育相关xnicroRNA进行筛选和鉴定,结果显示,oar-let-7和oar-miR-200家族的microRNA在妊娠45 d显著上调,表明其可能在毛囊发育中发挥关键作用。因此,基于circRNA、microRNA和mRNA问的关系,可推测有一些circ-RNA也参与调控绒毛生长发育。但其在绒毛生长发育中的作用机理未见报道。circRNA还有许多尚未知悉的重要生物学功能,其在各研究领域均具有巨大的应用潜力。
(责任编辑 陈燕)
