水稻短根突变体Osksr6的遗传分析和基因定位
2018-09-05诸周洁朱俊兆王晓飞丁沃娜
叶 金,诸周洁,姚 捷,朱俊兆,王晓飞,丁沃娜*
(1 宁波大学 海洋学院,浙江宁波 315211;2 宁波大学 科学技术学院,浙江宁波 315212;3 浙江农林大学 农业与食品科学学院,浙江杭州 311300)
水稻作为重要的谷类粮食作物,养育着地球半数以上人口。随着经济发展和人口数量增加,对水稻的质量与数量的要求与需求越来越高。然而水稻总产量增幅由于众多因素逐年下降,使得提高水稻产值仍是水稻研究者的优先任务[1-2]。水稻根系研究始于1919年,且在近半世纪得到迅速的发展,众多研究表明水稻根部特性与水稻地上部器官的生长发育及水稻单株产量息息相关[3-4]。
水稻根系主要分为主根、不定根、侧根和根毛四个部分。水稻主根由种子中的胚根发育形成(仅1条),主要在水稻幼苗期发挥作用,吸收土壤中水分及营养物质。因发根位置的不同,在水稻茎部发生的根称为不定根,数量较多[5],水稻不定根结构与主根基本相同[6]。在主根、不定根韧皮部的中柱鞘和内皮层中长出侧根[7]。根毛则是表皮细胞外伸形成的管状凸起[8]。侧根与根毛的存在增加了根的直径与根系的表面积,扩大了根系与根际土壤接触面积,使得根系可以吸收的水分、流动或不流动的营养物质范围更广[9]。
水稻根系结构形态受多种因素影响。首先根系发育受基因调控,随着水稻全基因组测序的完成,根系形态特征、生物学作用相关的基因被大量发现并克隆[10]。水稻根系形态也受环境因素的制约,根际含氧量可以影响水稻根长与其生物量[11-12];在含水量低的土壤中,根系总量增多且根系较长[13];过低温与过高温均抑制根系伸长,在一定温度范围内,根系伸长量与温度成正相关[14]。不同元素也影响根系生长,氮元素以NO3-形式存在时,有利于根系的伸长[15];铝元素对杂交水稻根系伸长具有负调节作用[16];硅元素对根系的长度与粗度具有促进作用[17]。水稻根系发育的调控机制很复杂,目前人们对其认知还很有限,更多水稻根系发育基因的克隆将有助于阐明作物根系发育调控的分子机理。
本研究选用从EMS(ethyl methane sulfonate)诱变的籼稻Kasalath突变体库筛选而来的短根突变体Osksr6作为研究材料。苗期,Osksr6的根系与野生型有着明显差异,主根显著变短、侧根发生与根毛伸长受不同程度的抑制。在此基础上,对Osksr6进行遗传分析与基因定位,为该基因的克隆和分子机理研究奠定基础。
1 材料和方法
1.1 水稻材料准备与培养
水稻短根突变体Osksr6,是EMS诱变的籼稻品种Kasalath突变体库中筛选获得的,其短根突变性状经几代种植,确认可稳定遗传。将可稳定遗传的突变体Osksr6与野生型籼稻Kasalath杂交产生的F2代群体用于遗传分析;将Osksr6与粳稻品种日本晴Nipponbare杂交产生的F2代用于突变基因定位。
水稻种子用0.6%稀硝酸进行破休眠处理,待露白后播种于水稻培养液进行溶液培养[18]。人工气候室培养条件:白天30 ℃ 12 h,夜晚22 ℃ 12 h,湿度为80%。
1.2 方 法
1.2.1突变体Osksr6的表型观察和统计各取20株野生型Kasalath与突变体Osksr6生长7 d的幼苗,分别统计株高、主根长度、不定根长度、不定根数量。用照相机(Nikon D70s)观察并拍摄具代表性的全株表型和根部性状,且统计侧根数量。用体式镜(Leica MZ95,Germany)对水稻根茎结合部,主根中段和根尖进行拍摄。
1.2.2突变体Osksr6的遗传分析突变体Osksr6与野生型Kasalath杂交,产生的F1代与野生型有相似的长根性状,取F1代自交产生的F2代种子400颗,播种于人工气候室。取生长7 d的F2代幼苗,统计长根、短根数量,用卡方检测分离比,确定显隐性及是否为单基因突变。
1.2.3突变基因定位(1) DNA的提取。将Osksr6与粳稻品种日本晴Nipponbare杂交产生的F2种子萌发后用水稻培养液培养7 d,挑取短根表型的幼苗,采用TPS提取法提取单株幼苗的DNA[19]。选取其中30株苗的DNA,把浓度稀释至100 ng/μL,每一管DNA吸取10 μL放入1.5 mL离心管中形成突变池,同时提取野生型Kasalath、日本晴Nipponbare、F1代幼苗的DNA。
(2) 分子标记的选择与设计。根据http:// www.gramene.org/网站中已公布的SSR简单重复序列,选取98对实验室已有的均匀覆盖水稻12条染色体且各具特异性的SSR引物,用于突变基因的初定位。在已定位到的水稻染色体SSR分子标记附近,根据已公布的籼稻品种9311、粳稻品种日本晴全基因组序列对比有差异两侧设计InDel标记,进行突变基因的精细定位。
(3) PCR扩增和电泳。将提取的野生型、日本晴、F1代和F2代群体的DNA,进行PCR扩增。扩增体系包括:模板DNA、10×PCR buffer、25 mmol/L MgCl2、2.5 mmol/L dNTP、上下游引物、5 U/mLTaqDNA聚合酶、超纯水。PCR反应条件:94 ℃预变性4 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,34个循环;72 ℃延伸5 min。PCR产物于6%非变性聚丙烯凝胶(PAGE)电压190 V条件下电泳,3 h后银染显色分析并拍照记录。
2 结果与分析
2.1 突变体Osksr6的表型鉴定
通过比较溶液培养生长7 d的幼苗发现,Osksr6的地上部分和野生型Kasalath无明显差异,但根系差异显著。Osksr6主根长度为野生型的38.01%,不定根长度为野生型的 53.58%(表1)。在体式镜下观察根部表型,可知突变体根毛变短,侧根发生和伸长都受抑制(图1)。
比较成熟期野生型和突变体Osksr6的农艺性状发现:Osksr6的分蘖数明显减少,株高与野生型相比无显著差异。此外,Osksr6的总穗长和结实率均不如野生型(图2、表2)。由此可得,Osksr6的基因突变不仅影响根系发育,而且影响水稻的分蘖及产量。

a.野生型(左)和突变体Osksr6(右)全株表型(标尺=1 cm);b.野生型(左)和短根突变体Osksr6(右)根部表型(标尺=1 cm);c~f.野生型(c, e)和Osksr6(d, f)的主根体视镜照(标尺=1 mm)图1 野生型和突变体Osksr6生长7 d表型a. Seedlings of the wild type (left) and Osksr6 mutant (right), bar=1 cm; b. The root of wild type (left) and Osksr6 mutant (right), bar=1 cm; c- f: The primary root of wild type (c, e) and Osksr6 (d, f) under stereocope, bar=1 mmFig.1 Phenotypic characterization of 7-day old seedlings of wild type and Osksr6 mutant

a.野生型(左)和突变体Osksr6(右)全株表型(标尺=10 cm);b.野生型(左)和突变体Osksr6(右)稻穗表型(标尺=2 cm)图2 野生型和突变体Osksr6成熟期表型a. Phenotypic characterization of the wild type (left) and Osksr6 (right), bar=10 cm; b. Panicle characterization of the wild type (left) and Osksr6 (right), bar=2 cmFig.2 Mature phenotypes of wild type and Osksr6 mutant

性状Trait 野生型 WTOsksr6较野生型增减Compared with WT/%株高Plant height/cm17.80±0.9517.77±0.59-0.17主根长Primary root length/cm13.47±0.635.12±0.66-61.98*不定根长aAdventitious root length/cm3.77±0.432.02±0.38-46.42不定根数Adventitous root number3.66±0.634.05±0.62+10.66侧根数Lateral root number195.02±9.3146.18±5.67-76.32*
注:a最长3根不定根长度的平均值;数值后*表示0.05水平显著性差异;下同
Note:aAverage of three longest adventitious roots on each plant; * means siginificonce difference at 0.05 level; the same as below

表2 野生型和突变体Osksr6的农艺性状参数
2.2 突变体Osksr6的遗传分析
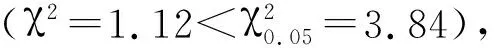
2.3 突变体Osksr6的基因定位
以混合30株短根突变体DNA形成的突变池、Kasalath、Nipponbare和F1的DNA为模板,用98对SSR引物逐一进行验证后发现,突变基因与第3染色体上SSR标记RM5813存在连锁关系(图3)。扩大定位群体,增加了142株短根单株,同时根据籼稻品种9311和粳稻日本晴基因组差异性,在RM5813物理位置附近设计了5对有多态性的InDel标记(表4),最终将突变基因定位在InDel标记28420k和28880k之间(图4),物理距离约460 kb(图5),分析表明该区间没有已报道的与根系发育相关的基因。
3 讨 论
根系是水稻尤为重要的器官,影响地上部的性状及产量。本研究的短根突变体Osksr6在幼苗期的主根、不定根、侧根和根毛的生长发育均受到抑制,分蘖数、稻穗长和结实率等成熟期农艺性状受到显著影响,说明OsKSR6基因呈一因多效,是一个与水稻生长发育和单产密切相关的重要功能基因。

表3 短根突变体Osksr6/ Kasalath F2的遗传分析
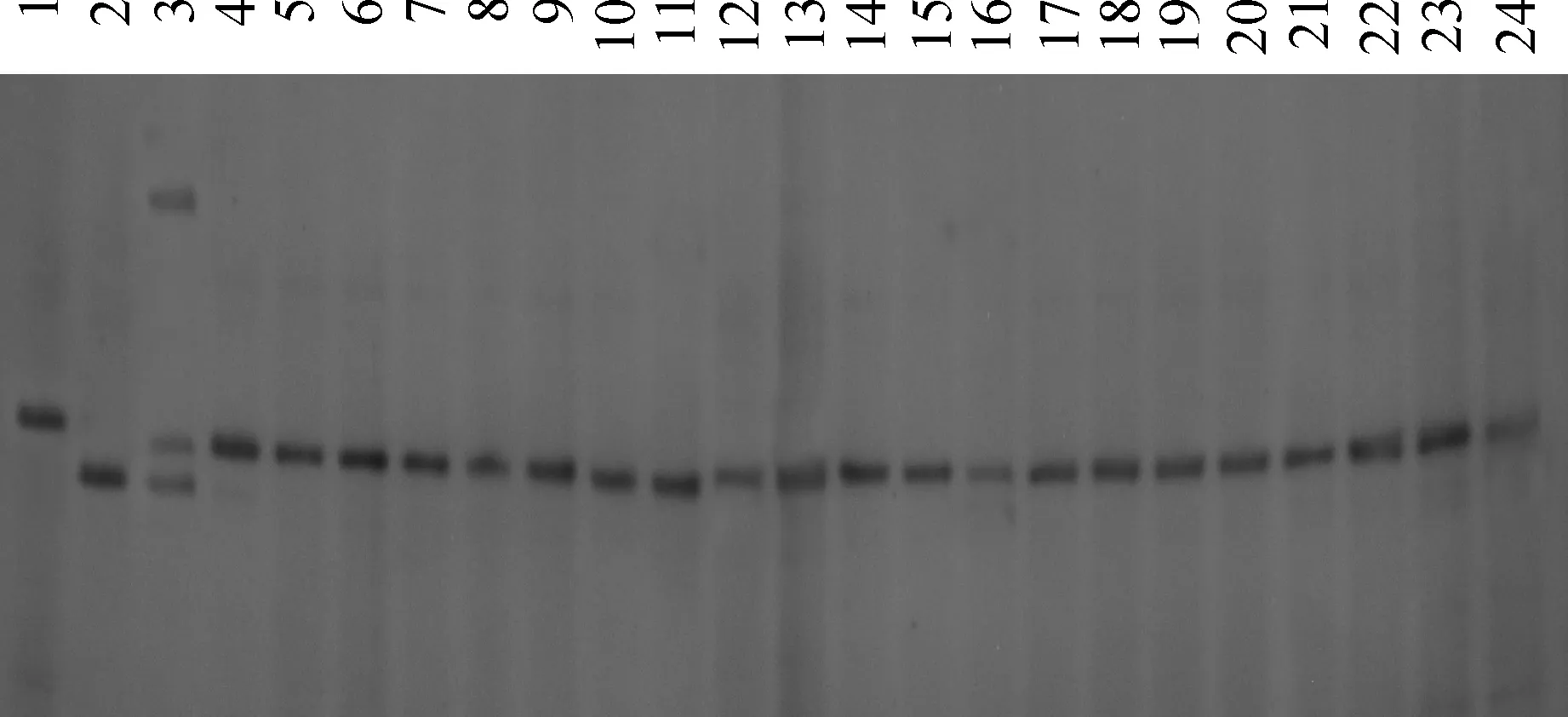
1. Kasalath;2. Nipponbare;3. F1(Osksr6×Nipponbare);4~24. F2短根单株图3 同OsKSR6基因连锁的SSR标记RM5813的电泳图1. Kasalath;2. Nipponbare;3. F1 (Osksr6 × Nipponbare);4-21. Short root individuals in F2 populationFig.3 Electrophoresis of the SSR marker RM5813 linked with the OsKSR6 locus

表4 具有多态性的SSR及InDel标记序列

图5 OsKSR6在水稻第3染色体上的分子定位Fig.5 The gene mapping of OsKSR6 on rice chromosome 3
根据已报道的水稻根系相关基因对应的突变体表型,与突变体Osksr6具有类似表型的基因有:OsSPR1[20]、OsKSR5[21]、OsCYT-INV1[22]、GLR3.1[23]、OsWOX11[24]和OsRAL1[25]等。OsSPR1基因位于第1染色体,突变体Osspr1第7天幼苗的主根与不定根变短,侧根发生受抑制,成熟期植株较野生型变矮,分蘖数显著减少,结实率也略有降低[20]。OsKSR5基因位于第1染色体,突变体Osksr5生长7 d时,主根长度为野生型的44.44%、不定根长度为野生型的53.44%、侧根明显变短;成熟期,突变体株高变矮(为野生型的59.90%),分蘖数大幅减少,结实率为88.82%[21]。OsCYT-INV1基因位于第2染色体,幼苗期突变体表现出主根、不定根、侧根均变短;花期延迟,花粉部分不育;成熟期分蘖数减少,结实率仅为11.5%[22]。GLR3.1基因位于第4染色体,发芽后的2周,与野生型相比突变体主根、侧根、不定根均受到不同程度的抑制,突变体主根根尖直径明显变小;苗期之后与繁殖阶段,野生型与突变体新生根系长度相当;成熟期分蘖数与结实率受影响[23]。OsWOX11基因位于第7染色体,该基因突变后幼苗表现出严重的生长缺陷,发芽后2周突变体几乎没有根冠产生;成熟阶段植株高度明显降低,分蘖数减少,结实率大幅下降[24]。OsRAL1基因突变,突变体表现出主根缺失,不定根、侧根减少;成熟期分蘖数明显减少[25]。
本研究通过图位克隆技术,最终将OsKSR6基因定位在第3染色体上的InDel标记28420k和28880k之间约460 kb范围内。目前在该区域还未发现有关根系发育基因的报道,所以OsKSR6基因是一个调控水稻根系发育的新基因。对OsKSR6基因的进一步研究,将有助于阐明水稻根系发育的分子机理,也为提高水稻单株产量提供基因资源。
