多不饱和脂肪酸日粮中添加酵母硒对湖羊脂肪酸组成和抗氧化的影响
2018-09-03樊懿萱邓凯平澹台文静黄欣爱
樊懿萱,邓凯平,澹台文静,黄欣爱,王 锋*,陈 明*
(1. 南京农业大学 羊业科学研究所,南京 210095;2. 江苏农牧科技职业学院,泰州 225300)
随着人们保健意识的增强,对饮食品质的要求越来越高,羊肉因富含蛋白质、钙、钾和维生素B1等,且脂肪、胆固醇含量低,其市场需求量日益增加,现已成为人们日常膳食肉类的主要来源之一。因此,不断丰富市场上高端羊肉产品的数量与种类,进一步满足消费者对绿色、健康、保健肉产品的需要,成为研究者们需要解决的主要问题之一。研究表明,多不饱和脂肪酸(polyunsaturated fatty acids, PUFAs) 在抑制肿瘤血管发生、冠状动脉硬化等过程中均发挥重要作用,是人体生长和健康所必需的物质,因人体不能合成而成为人体必需脂肪酸[1-2]。因此,通过日粮添加富含PUFAs的油脂、籽实或饼粕,提高反刍动物体组织中不饱和脂肪酸的沉积[3],获得有益于人类健康的奶或肉制品,成为近年来研究热点[4]。紫苏籽富含α-亚麻酸,出油率高达45%,具有抗氧化、抗炎和抗菌抗病毒等功效。然而,反刍动物瘤胃微生物对PUFAs的氢化作用,致使体组织沉积大量饱和脂肪酸(saturated fatty acids, SFAs),而且,肌肉中PUFAs 增加容易引发脂质氧化,造成组织损伤,引起酸败不易保存。因此,饲料添加 PUFAs 提高反刍动物体组织中 PUFAs沉积的同时,应添加抗氧化物质,防止不饱和脂肪酸在体内的过氧化损失。
硒是谷胱甘肽过氧化物酶的活性中心,具有很强的抗氧化功能,酵母硒以其吸收率高、污染小、易在动物体内储存等优点成为常用硒源。日粮中添加硒可以提高肉仔鸡、奶牛及羊等动物体内硒含量和抗氧化性能[5-7]。但是,酵母硒和PUFAs互作是否能增强机体抗氧化性能,降低脂质氧化腐败,提高羊肉中脂肪酸的沉积量,尚需进一步研究。本试验以湖羊为研究对象,探讨酵母硒和PUFAs 互作对湖羊生产性能、肌肉和肝脂肪酸组成及抗氧化性能的影响,为生产富含稳定PUFAs的高品质羊肉提供理论依据和实践指导。
1 材料与方法
1.1 试验设计与饲养管理
选取3月龄、体重为(23.02±1.36)kg、健康状况良好的湖羊公羔45只,随机分成3组,每组5个重复,每个重复3只羊,分别饲喂基础日粮(C组)、C+10% 紫苏籽(PFS组)和C+PFS+475 mg ·kg-1DM酵母硒(即0.75 mg ·kg-1硒)(PFS+Se组)。试验日粮参照国内肉羊饲养标准(NY/T816-2004)配制,以等能等氮形式搭配,具体配方见表1。试验在江苏省海伦羊业有限公司进行,整个试验期为70 d,其中预饲期10 d,正饲期60 d。试验前清洗、消毒羊舍,并对羊统一驱虫、健胃,试验羊采取自由采食和饮水。
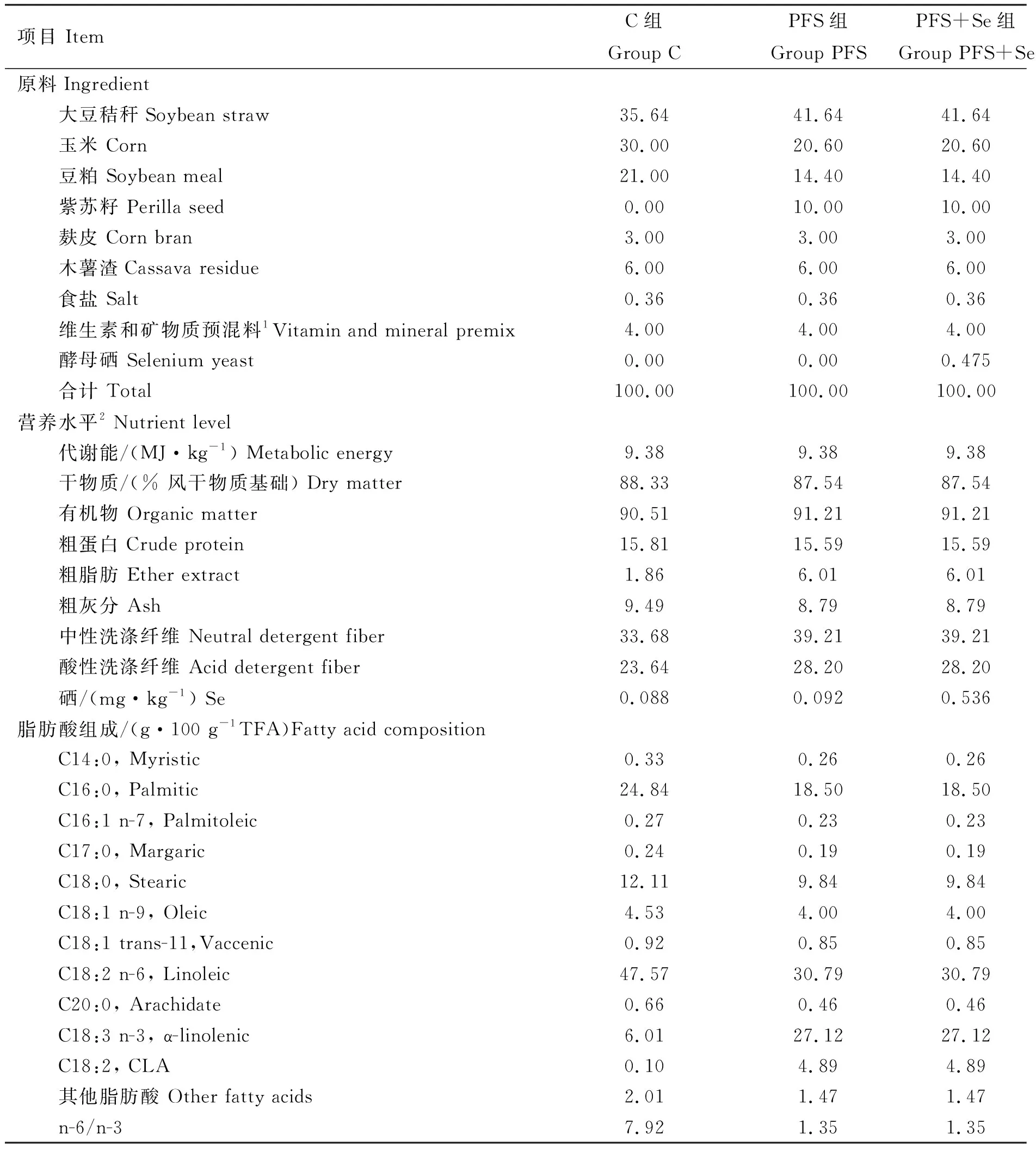
表1 日粮组成及营养水平(干物质基础)
1. 维生素和矿物质预混料为每千克饲料提供:维生素A 2 150 IU,维生素D 170 IU,维生素E 13 IU,铁56 mg,铜15 mg,锰30 mg ,锌40 mg,碘1.5 mg,硒0.2 mg,钴0.25 mg,硫3.2 g,超浓缩源康宝 (芽孢杆菌、乳酸菌、酵母菌等有益菌及其代谢产物蛋白酶、淀粉酶、纤维素酶、氨基酸、有机酸、维生素等,有效活菌含量≥2.0×1010CFU·g-1) 2.7 g,2%莫能霉素6 g,硫酸钠10.1 g。2. 代谢能是根据原料组成计算所得(干物质为基础),其他均为实测值
1. The vitamin and mineral premix provide the following per kg of diets: Vitamin A 2 150 IU, Vitamin D 170 IU, Vitamin E 13 IU, Fe 56 mg, Cu 15 mg, Mn 30 mg, Zn 40 mg, I 1.5 mg, Se 0.2 mg, Co 0.25 mg, S 3.2 g,super-concentrated Yuan Kangbao (bacillus, lactic acid bacteria and yeast bacteria and their metabolic products such as protease, amylase, cellulase enzyme, amino acids, organic acids, vitamins, etc. The content of effective living bacterium≥2.0×1010CFU·g-1)2.7 g, 2% monensin 6 g, sodium sulfate 10.1 g.2. Metabolic energy is calculated values by feed composition (DM basis), while the others are measured values
1.2 试验方法
1.2.1 样品采集与生产性能指标测定 以正试期第1 天的体重作为初始体重,之后每隔30 d分别对各组羊进行空腹称重,以试验结束时的体重为末重,并记录每组羊的给料量和剩料量。分别计算3组羊日干物质采食量(dry mater intake,DMI)、平均日增重(average daily gain,ADG)和料重比(F/G)。
在正试期最后1 d,各组羊禁食24 h,禁水2 h。屠宰前对所有羊进行颈静脉采血,采集的血样放入促凝管,静置于室温30 min,3000 r·min-1离心10 min 后转移血清,将血清置于-20 ℃冰箱冻存备用。羊只屠宰过程中,记录其宰前重、胴体重、背膘厚和胴体脂肪含量,并计算其屠宰率。屠宰后所有羊取同一部位肝、背最长肌,置于-80 ℃超低温冰箱冻存备用。
1.2.2 背最长肌常规营养成分和矿物元素指标测定 取背最长肌样品称重,通过烘箱、马福炉、全自动凯氏定氮仪、索氏脂肪提取器分别测定其中水分、粗灰分、粗蛋白和粗脂肪的含量;按照Reykdal等[8]的方法,使用电感耦合等离子体质谱仪(ICP-MS)(Agilent 7500ce; Agilent Technologies, Germany)测定肌肉中矿物元素的含量。
1.2.3 肌肉和肝脂肪酸组成指标测定 采用气相色谱法测定肌肉和肝脂肪酸组成。参考Folch等[9]的方法提取样品总脂肪酸。现做简单的描述,取1 g左右样品放入装有40 mL氯仿/甲醇溶液(v∶v为2∶1, 含有丁羟甲苯)的50 mL玻璃萃取管中,匀浆后过滤。然后加入10 mL 0.9% NaCl溶液便于液相分离,将下层澄清相收集到圆底烧瓶中,70 ℃ 水浴中旋转蒸发10~20 min,再添加内标二十一烷酸(C21:0; Sigma Chemicals, St. Louis, MO, USA)。将提取的脂肪酸按照AOAC[10]进行皂化和酯化以制备脂肪酸甲酯(fatty acid methyl ester, FAME)。制备好的FAME用气相色谱分析(GC-MS;Shimadzu QP 2010 Ultra)。采用100 m的FAME专用毛细管柱(Supelco SP 2560,100 m×0.25 mm×0.2 μm)分离甲酯,用FID检测,进口温度为250 ℃。管柱程序升温条件:在150 ℃下运行2 min,以1 ℃·min-1升温至158 ℃,维持28 min,然后以1 ℃·min-1升温至220 ℃,维持20 min使甲酯充分分离。载气(N2)流速为1.2 mL,分流比为1∶20。脂肪酸的种类通过保留时间与标准对比确定,按峰面积计算含量,脂肪酸结果用占总脂肪酸百分含量表示,g·100 g-1TFA。
1.2.4 肌肉和肝抗氧化性能指标测定
1.2.4.1 抗氧化酶指标测定:血清、肌肉和肝中的超氧化物歧化酶(superoxide dismutase, SOD)、过氧化氢酶(catalase, CAT)、谷胱甘肽过氧化物酶(glutathione peroxidase, GSH-Px/GPX)、总抗氧化能力(total antioxidant capacity, T-AOC)及氧化产物丙二醛(malondialdehyde, MDA)使用试剂盒(R&D Systems, USA)按照说明方法用分光光度计测定。
1.2.4.2 抗氧化基因表达量检测:使用TIANGEN 组织总RNA提取试剂盒提取肌肉和肝组织的总RNA,然后按照TaKaRa反转录试剂盒说明书进行反转录。采用SYBR Green法,以甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)为内参,对肌肉和肝进行SOD、CAT和GPX基因的荧光定量PCR分析。利用2-△△Ct法对基因表达进行相对定量。引物序列见表2。
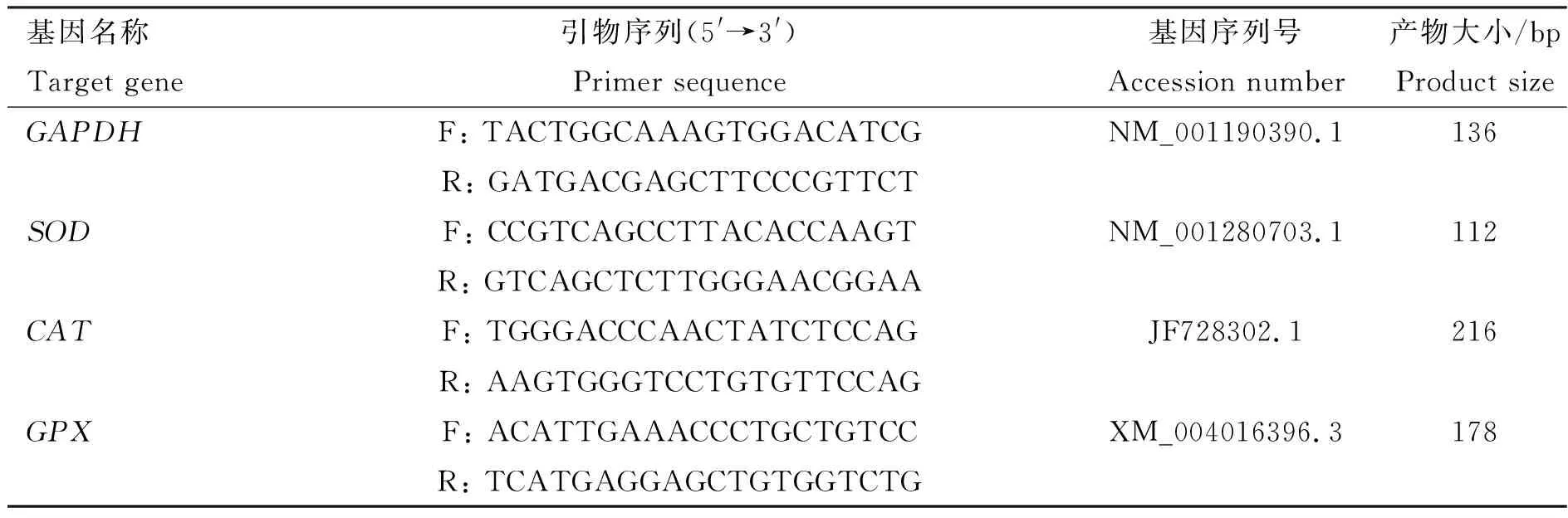
表2 Real-time PCR所用引物信息
1.3 数据处理与分析
试验数据用Excel 2013初步整理后,采用SPSS 19.0中ANOVA过程进行单因素方差分析,使用Duncan’s法进行多重比较,结果均用“平均值(Means)±标准差(SD)”表示,以P<0.05表示组间差异显著水平。
2 结 果
2.1 PUFAs日粮中添加酵母硒对湖羊生产性能的影响
根据表3和表4可知,C组、PFS组和PFS+Se组的湖羊在生长性能和屠宰性能方面的指标均无显著性差异(P>0.05)。
2.2 PUFAs日粮中添加酵母硒对湖羊肌肉常规营养成分和元素组成的影响
由表5可知,3组间试验羊背最长肌中水分、灰分和粗蛋白含量均无显著差异(P>0.05),但与C组相比,PFS组和PFS+Se组的背最长肌中粗脂肪含量分别显著提高了42.3%和46.8%(P<0.05);由表6可知,PFS+Se组湖羊背最长肌中硒含量显著高于PFS组(P<0.05),而PFS组背最长肌中锌含量显著高于C组(P<0.05),但C组与PFS+Se组背最长肌中锌含量无显著差异(P>0.05)。此外,3组之间背最长肌中Fe、Cu、Na、K和Mg等矿物质元素含量无显著差异(P>0.05)。
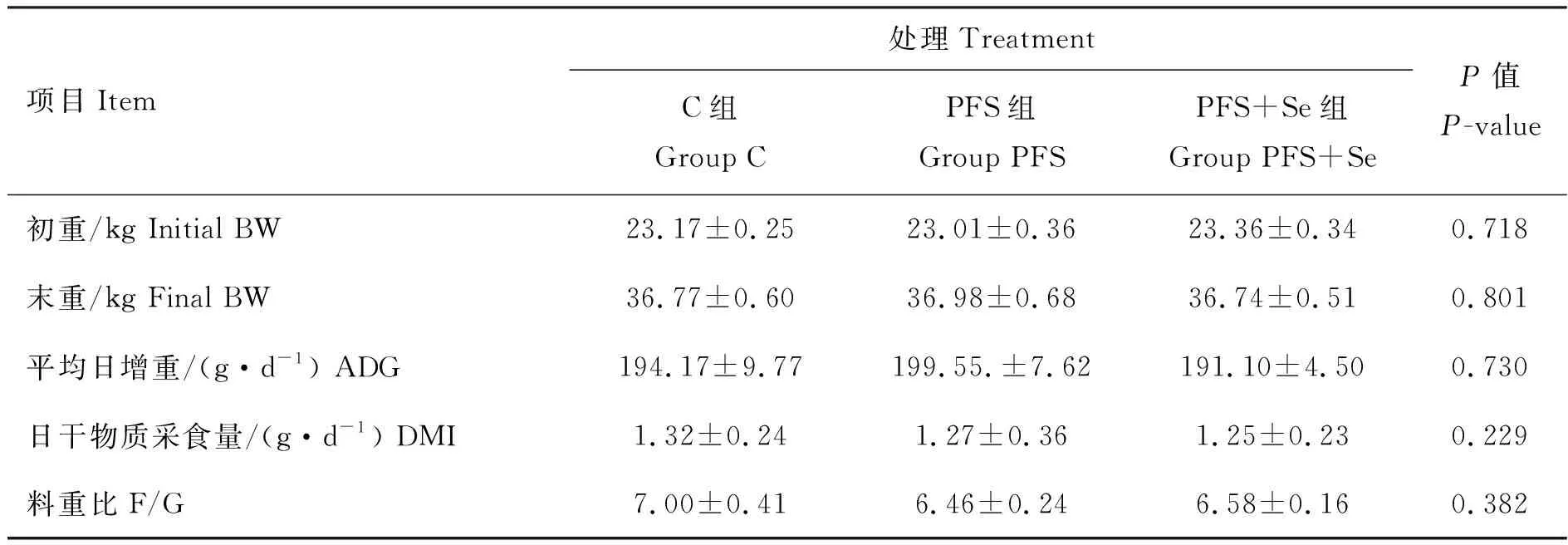
表3 PUFAs日粮中添加酵母硒对湖羊生长性能的影响
同行数据后所标字母相异表示差异显著(P<0.05),所标字母相同表示差异不显著(P>0.05)。下表同
Different letters in the same row means significant difference among the treatments(P<0.05), same letter in the same row means no significant difference among the treatments(P>0.05). The same as below
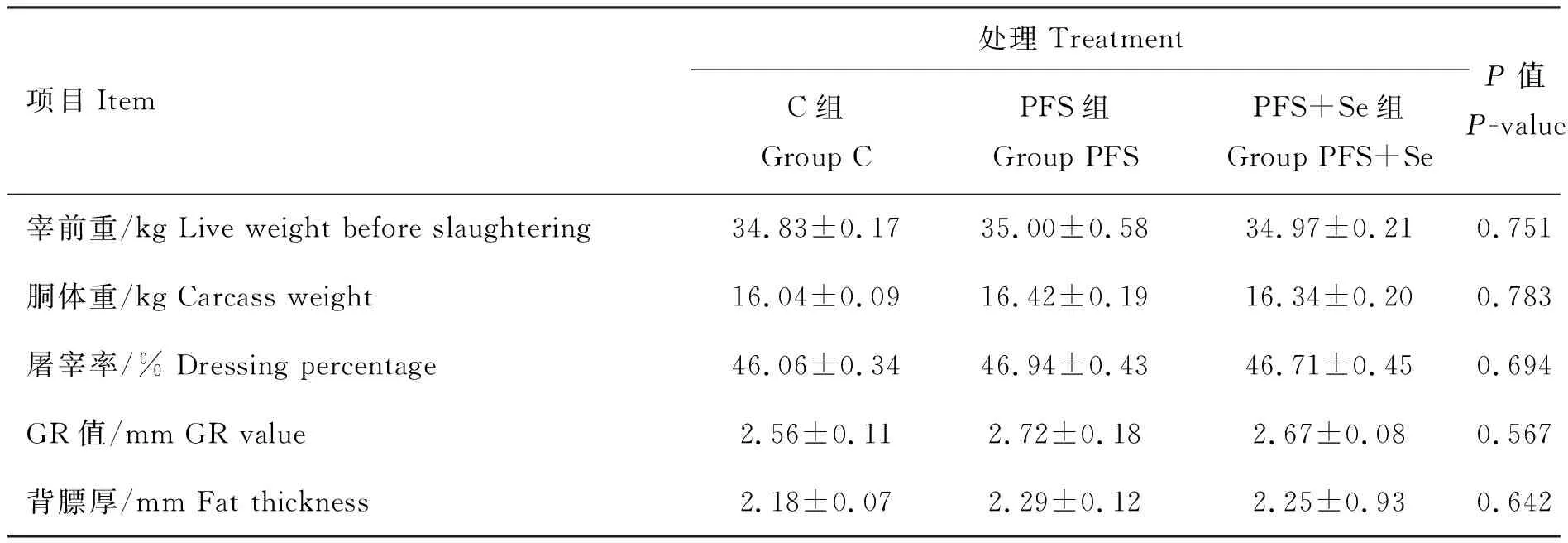
表4 PUFAs日粮中添加酵母硒对湖羊屠宰性能的影响
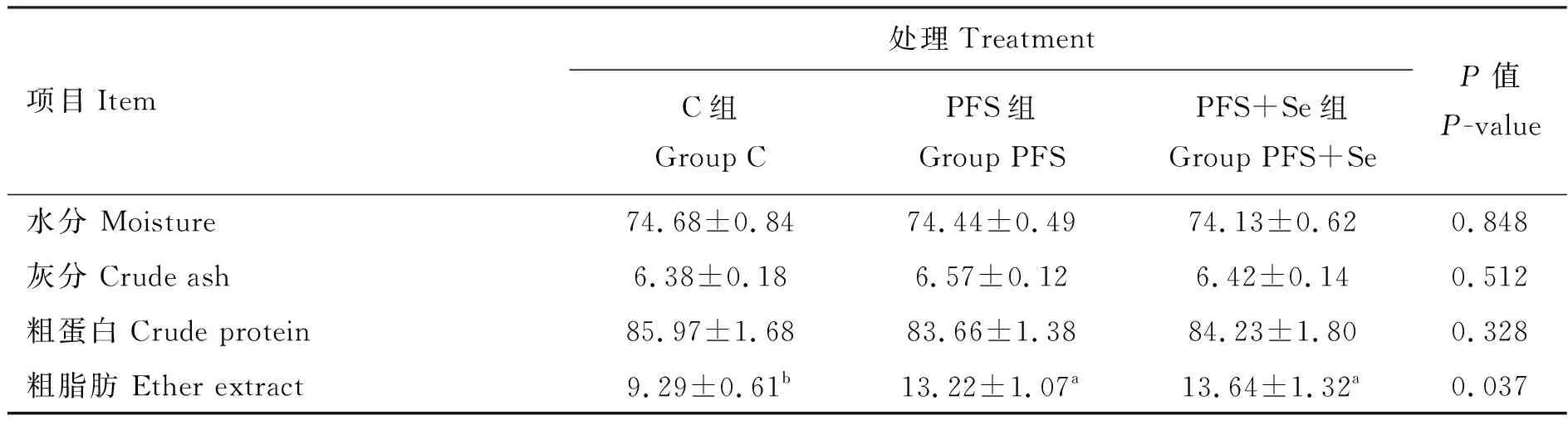
表5 PUFAs日粮中添加酵母硒对湖羊背最长肌常规营养成分的影响
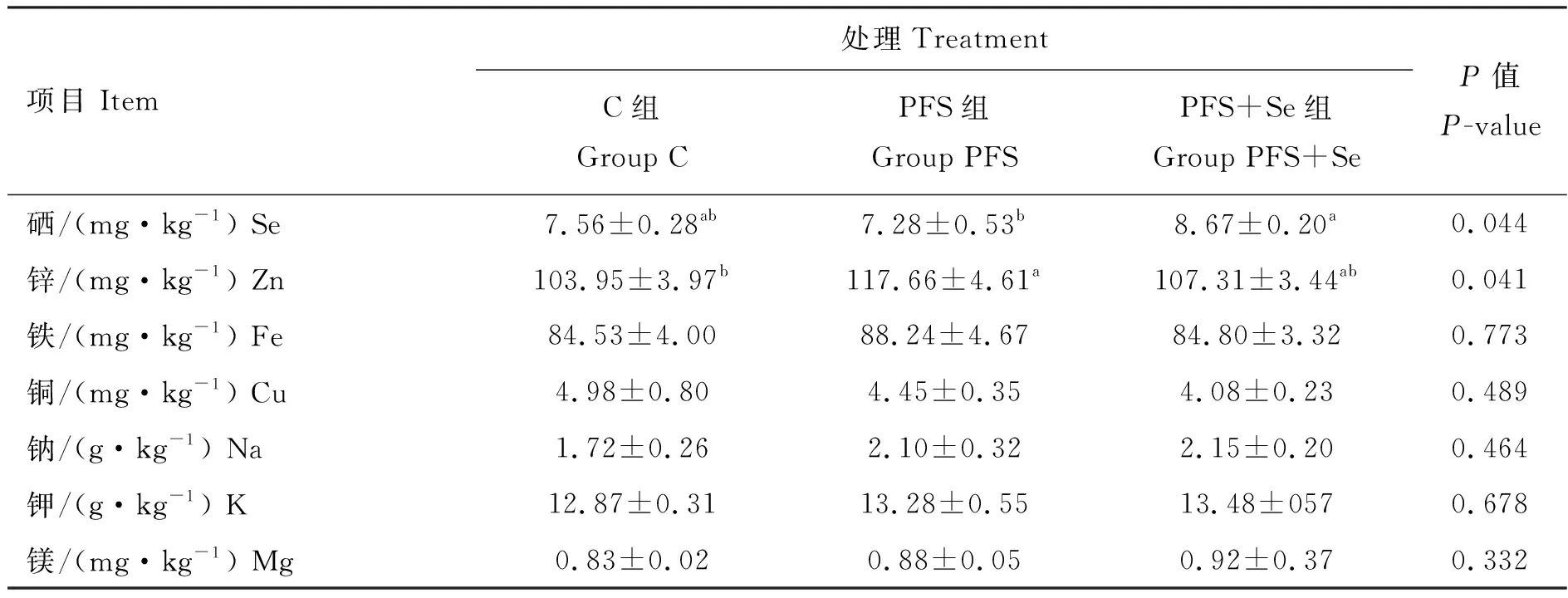
表6 PUFAs日粮中添加酵母硒对湖羊背最长肌矿物元素含量的影响
2.3 PUFAs日粮中添加酵母硒对湖羊肌肉和肝脂肪酸组成的影响
由表7可知,各试验组间湖羊背最长肌中总SFAs含量无显著差异(P>0.05),PFS组和PFS+Se组中背最长肌C17:0含量显著低于C组(P<0.05)。与C组相比,PFS组和PFS+Se组总MUFAs含量显著升高,其中C18:1cis9含量显著升高,C18:1trans-11含量显著降低(P<0.05)。3个试验组间总PUFAs含量无显著差异(P>0.05),但PFS组和PFS+Se组总n-6 PUFAs含量显著降低,总n-3 PUFAs含量显著升高(P<0.05),因此n-6/n-3 显著降低(P<0.05);n-6 PUFAs中的C18:2n-6和C20:4n-6的含量均显著降低,但C18:2n-6cis含量显著升高(P<0.05);n-3 PUFAs中的C18:3n-3含量显著升高(P<0.05),PFS组C20:5n-3含量显著高于C组(P<0.05),但PFS+Se组与其他两组均无显著差异(P>0.05)。
表8结果显示,PFS组和PFS+Se组湖羊肝中总SFAs含量显著下降(P<0.05),其中C12:0、C14:0、C15:0、C16:0、C17:0、C18:0的含量显著下降,但C23:0含量显著升高(P<0.05),与C组相比,PFS组的C24:0显著升高(P<0.05),但PFS+Se组C24:0的含量与C组无显著差异(P>0.05)。与C组相比,PFS组和PFS+Se组总MUFAs含量显著升高,其中C18:1cis9的含量显著升高,但C18:1trans-11含量显著下降(P<0.05)。与C组相比,PFS组和PFS+Se组湖羊肝中总PUFAs的含量显著升高(P<0.05),其中,PFS组和PFS+Se组总n-6 PUFAs含量显著降低,总n-3 PUFAs含量显著升高(P<0.05),因此n-6/n-3显著降低(P<0.05);n-6 PUFAs中的C18:2n-6和C20:4n-6的含量均显著降低(P<0.05); C18:3n-3、C20:5n-3、C22:6n-3和C22:5n-3等n-3PUFAs含量显著升高(P<0.05)。
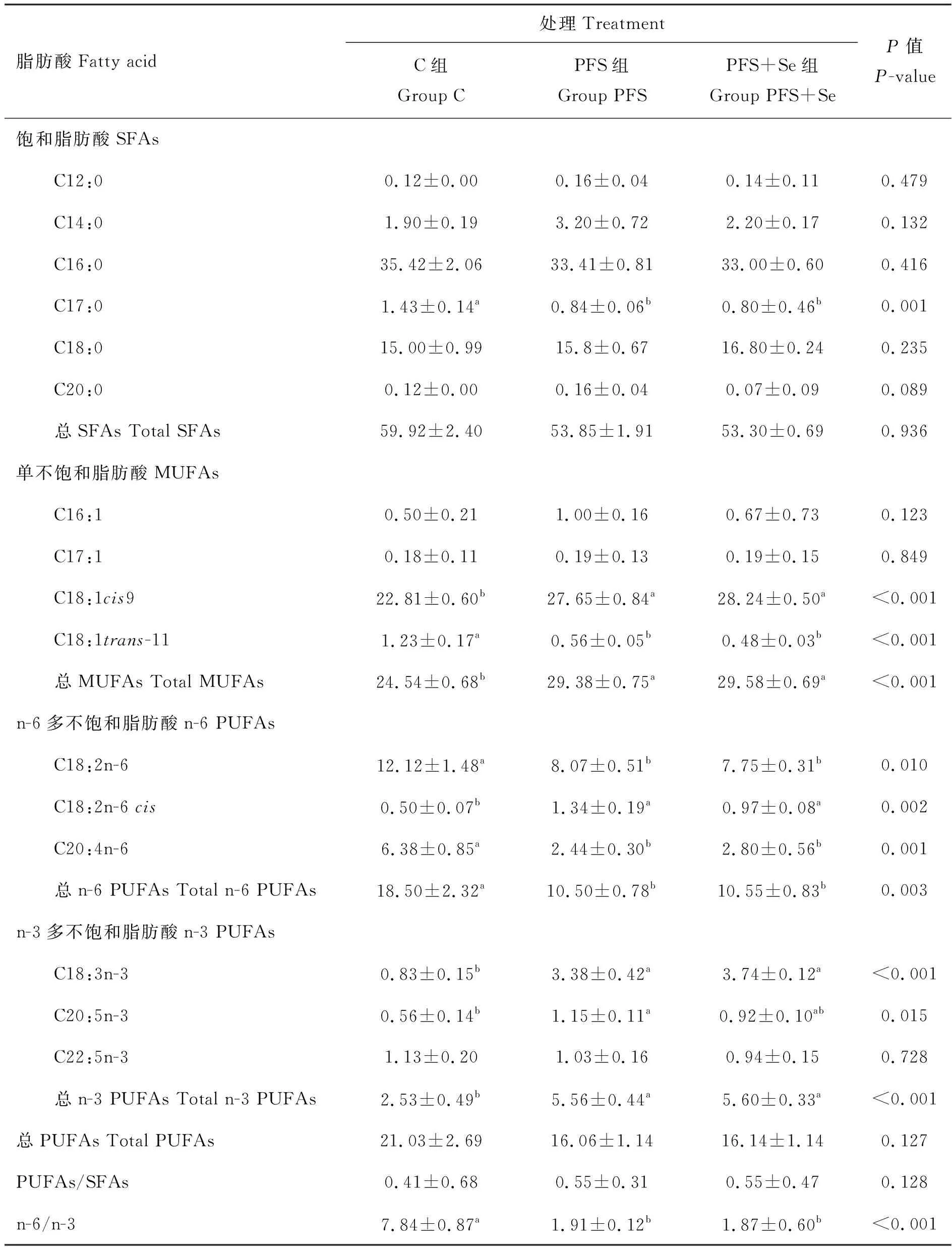
表7 PUFAs日粮中添加酵母硒对湖羊背最长肌脂肪酸组成的影响
2.4 PUFAs日粮中添加酵母硒对湖羊抗氧化性能的影响
从表9可见,血清、肝和肌肉中SOD和GPX的活力在3组之间均无显著差异(P>0.05),PFS+Se组湖羊血清中CAT活性在3组中最高(P<0.05);并且PFS+Se组湖羊肝中MDA含量显著低于C和PFS组(P<0.05)。
由图1可见,与C组比较,PFS组和PFS+Se组的湖羊肌肉中SOD和GPX的表达量均有所提高,而CAT表达有所下降,但均无显著性差异(0.1 >P>0.05) 。从图2可以看出,PFS+Se组湖羊肝中GPX和CAT表达量显著高于其他两组(P<0.05),而SOD在3组间的表达量无显著差异(P>0.05)。
3 讨 论
3.1 PUFAs日粮中添加酵母硒对湖羊生产性能的影响
目前关于在不饱和脂肪酸日粮中添加酵母硒对反刍动物生产性能影响的研究结果不一致。Benchaar等[11]在奶牛日粮中添加不同水平的亚麻籽油,对奶牛DMI无显著影响。Lunsin等[12]在水牛日粮中添加6%的米糠油降低了水牛的DMI。Kamada[13]在泌乳中期的荷斯坦奶牛日粮中添加有机硒,对奶牛DMI、产奶量和乳脂率等均没有显著影响。卓春燕[14]在含葵花籽油(350(g·头-1)·d-1)的奶牛日粮中分别添加8、15、22(g·头-1)·d-1硒对牛奶中乳脂、乳蛋白、乳糖等生产性能没有显著影响。本试验结果表明,在紫苏籽日粮中添加酵母硒对湖羊生长性能和屠宰性能方面的指标均无显著影响,这与Benchaar等[11]、Kamada[13]的研究结果相符。上述结果差异可能与反刍动物对硒的利用率有关,也可能与日粮中原有的硒水平有一定的相关性,若日粮中硒含量未能达到动物生理需求,则添加酵母硒可能促进其生长。
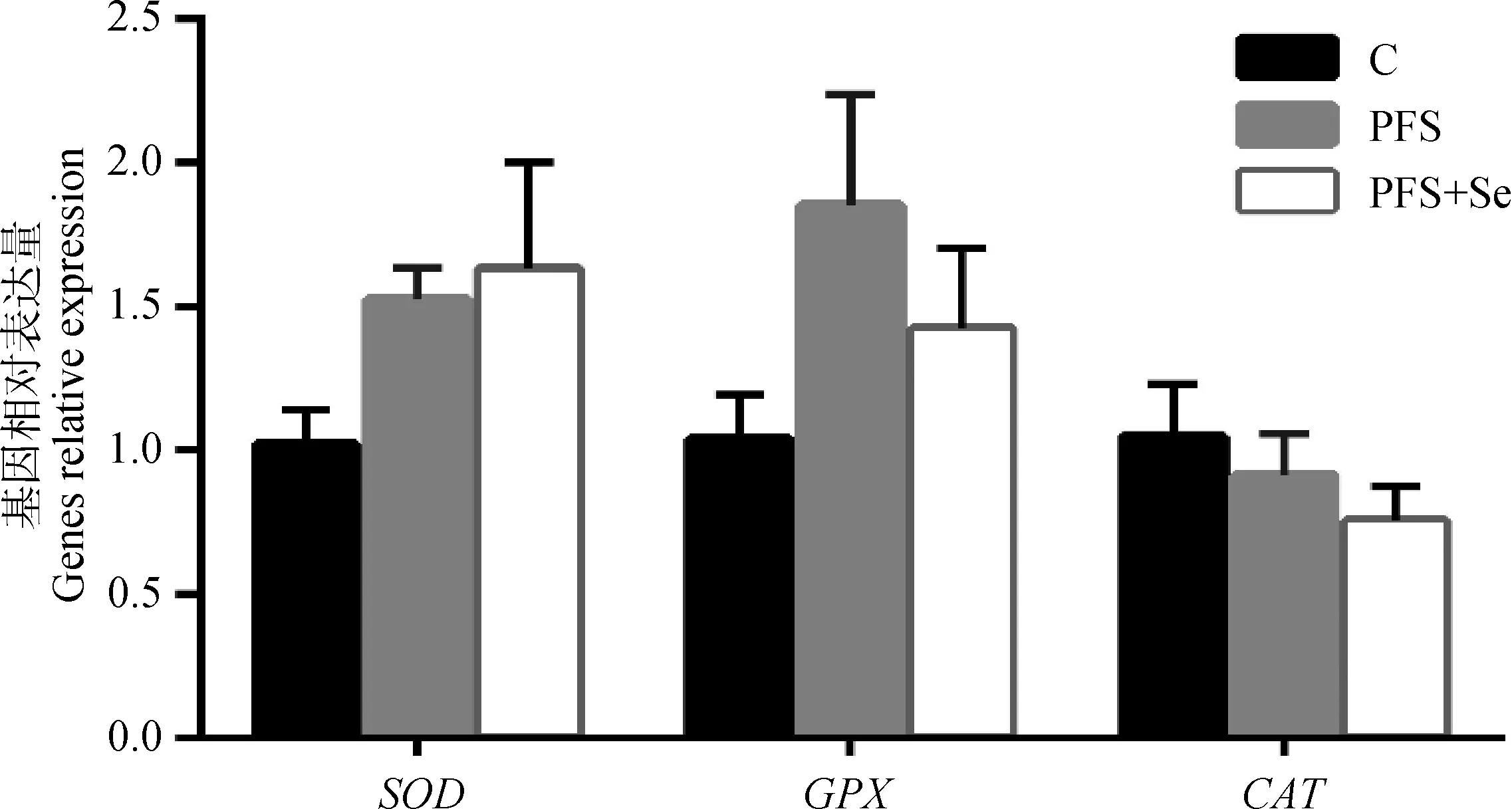
图1 PUFAs日粮中添加酵母硒对湖羊背最长肌抗氧化基因表达量的影响Fig.1 Effect of selenium yeast supplementation in containing PUFAs dietary on the expression of antioxidant genes in LD muscle of Hu lambs
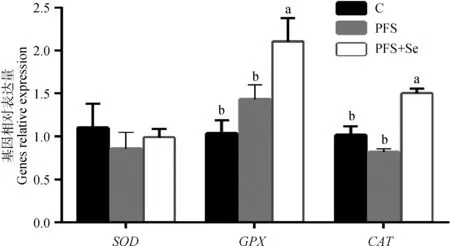
图2 PUFAs日粮中添加酵母硒对湖羊肝抗氧化基因表达量的影响Fig.2 Effect of selenium yeast supplementation in containing PUFAs dietary on the expression of antioxidant genes in liver of Hu lambs
3.2 PUFAs日粮中添加酵母硒对湖羊背最长肌常规营养成分和元素组成的影响
本试验结果表明,在紫苏籽日粮中添加酵母硒使湖羊背最长肌中粗脂肪含量提高了46.8%,但3组试验羊在水分、灰分和粗蛋白含量上无显著差异,说明日粮中添加紫苏籽可促进湖羊肌间脂肪沉积,但在紫苏籽日粮中添加酵母硒对改善肌肉营养成分并无显著影响。武雅楠等[15]在羔羊日粮中添加5%和10%亚麻籽也显著提高了羔羊肌肉中粗脂肪含量,与本研究结果一致。
酵母硒相对于无机硒来说,吸收率高且易在动物组织中储存,对环境造成的污染小,是一种理想的硒源添加物,在鸡、猪日粮中的添加均提高了肌肉硒含量[16-17]。Qin等[18]研究发现,添加酵母硒、富硒益生菌和亚硒酸钠均能显著提高肥育羔羊肾、肝和肌肉中硒含量。本试验结果表明,在紫苏籽日粮中添加酵母硒可提高硒在湖羊背最长肌内的沉积量,与上述结果一致。
3.3 PUFAs日粮中添加酵母硒对湖羊肌肉和肝脂肪酸组成的影响
在日粮中添加PUFAs的油脂、籽实或饼粕是提高反刍动物体组织中不饱和脂肪酸沉积,获得有益于人类健康的奶或肉制品的有效手段。Gómez等[19]在荷斯坦公牛日粮中添加亚麻籽,其肌肉和皮下脂肪中n-3PUFAs显著增加,n-6PUFAs/n-3PUFAs显著降低。Andrés等[20]对美利奴育肥羔羊的研究也表明,亚麻籽可以增加其背最长肌胸段的n-3 PUFAs含量。肌肉脂肪酸组成反映了日粮脂肪酸来源,能较好的反映羊肉脂肪酸营养状况。肝是动物体新陈代谢的一个中心站,所有维持生命力的各种养分都是在肝里产生,能较好地反映机体的脂肪酸营养状况,并能反映其它组织脂肪酸成分的变化情况。本试验结果表明,日粮中添加10%紫苏籽可促进湖羊肌肉和肝中n-3PUFAs含量提高,n-6PUFAs及n-6PUFAs/n-3PUFAs显著降低,与以上结果一致。紫苏籽日粮中亚麻酸浓度较高,被吸收后在体内可经过一系列的延长及脱氢作用生成 C18: 3n-3 及C22: 6n-3,因此底物浓度增加,相应提高了产物浓度,使饲喂紫苏籽日粮湖羊肌肉和肝中含有较高浓度的C18: 3n-3、C20: 5n-3及C22: 6n-3。此外,本研究发现,饲喂紫苏籽日粮湖羊的肌肉和肝中C18:1cis9含量显著提高。当日粮中亚麻酸含量较高时一部分亚麻酸就会在瘤胃微生物的氢化作用下产生较高含量的油酸,而另外一部分则直接以亚麻酸的形式进入机体,最终沉积在脂肪中[21-22]。油酸在生物氢化过程中存在异构化作用,可产生双键位置从C6到C16的多种反式C18:1异构体,但双键位置易受到瘤胃环境的影响,当瘤胃pH或流通速率降低时,双键位置被限制的低于C10。Kellens等[23]在绵羊日粮中添加油菜籽后发现,肌肉中C18:1的含量得到显著提高,与本试验结果一致。
反刍动物日粮中多不饱和脂肪酸进入瘤胃后,在微生物的作用下发生氢化反应,导致PUFA在反刍动物副产品(肉、奶等) 中的沉积减少。酵母硒具有很强的抗氧化功能,通过调控日粮中多不饱和脂肪酸在瘤胃中的氢化过程,以提高动物副产品中有益脂肪酸的含量。Papadomichelakis等[24]研究发现,日粮中添加硒可增加兔肌肉中PUFAs的含量,Jiang等[25]证明,杂交猪日粮中同时添加亚麻籽油和有机硒,猪肉中n-6/n-3脂肪酸比例显著下降。于玲玲[26]试验表明,葵花籽油日粮添加2 mg·kg-1DM的有机硒可提高皮下脂肪和网膜中C18:1cis9和总PUFA的浓度。本研究发现,在紫苏籽日粮中添加酵母硒对湖羊肌肉和肝脂肪酸的沉积量无显著影响。这可能是因为紫苏籽日粮中含有具有抗氧化性能的迷迭香酸、咖啡酸等酚酸类物质[27],在一定程度上可防止不饱和脂肪酸氧化。其次,可能是因为饲喂紫苏籽日粮的湖羊体组织中含有较多的n-3PUFAs,其三键结构比饲喂葵花籽油日粮羊体组织中含量较多n-6PUFAs的双键结构更易发生氧化反应,导致了 PUFAs 的流失。因此,酵母硒未能有效保护饲喂紫苏籽日粮湖羊体组织中n-3PUFAs的过氧化降解,不能增加其在体组织中的沉积。由此说明,硒对不同来源PUFAs日粮的抗氧化能力有差异,酵母硒可能不是紫苏籽日粮有效的抗氧化剂。于玲玲等[28]研究发现,日粮中添加硒提高了饲喂葵花籽油日粮绵羊血浆中C20: 4n-6 和红细胞膜中 C20: 5n-3 浓度,而对饲喂亚麻油日粮的绵羊无影响,与本研究结果相似。
3.4 PUFAs日粮中添加酵母硒对湖羊抗氧化性能的影响
PUFAs因其结构上的多双键或三键,易发生脂质过氧化反应,生成过氧化自由基,产生一系列的有害物质。硒最主要的功能是作为 GPX 的活性中心发挥抗氧化作用,降解体内脂质过氧化产物。因此,本试验在饲喂富含亚麻酸的紫苏籽的同时,通过添加酵母硒来达到降低体内脂质过氧化程度的目的。
GPX 是体内一种重要的抗氧化酶,对自由基的清除具有重要作用。CAT是过氧化物酶体的标志酶,催化细胞内过氧化氢分解,防止过氧化。T-AOC是指由GPX、CAT等酶促体系和由维生素C、维生素 E、谷胱甘肽等非酶促体系组成的总的抗氧化能力。MDA 是体内自由基攻击生物膜中PUFAs,引发脂质过氧化产生的一类较稳定的脂质过氧化物。秦顺义等[29]证实,日粮中富硒益生菌的添加提高了羔羊血浆中T-AOC和SOD的活性,降低了MDA的水平;石磊等[7]在妊娠牧羊血清中添加酵母硒,其血清中T-AOC、GPX和SOD活性提高,MDA含量降低。本试验中,血清、肝和肌肉CAT和 T-AOC活性随日粮中硒添加量的增加而升高,抑制或者清除了脂质过氧化产生的MDA,降低了肝中 MDA 的含量,从而抑制脂质过氧化反应。
此外,为了进一步探讨酵母硒与抗氧化酶基因表达的关系,本试验测定了湖羊肌肉和肝中抗氧化基因SOD、GPX和CAT的表达量,研究发现,紫苏籽日粮中添加酵母硒显著提高湖羊肝中GPX和CAT基因表达量。湖羊肝中CAT基因表达量提高,肝中过氧化物产物MDA的含量显著减少,两者结果一致,这表明,酵母硒可以通过上调湖羊肝中CAT和GPX基因的表达来增强机体抗氧化酶活性,提高抗氧化能力,抑制或者清除了脂质过氧化产生的MDA,增强对外界环境氧化应激的抵抗能力。但是,酵母硒添加组湖羊肌肉中SOD和GPX基因表达量升高,在肌肉抗氧化酶的活性上并无表现,其中机理还需进一步研究。
4 结 论
育肥湖羊饲喂富含亚麻酸的日粮(紫苏籽)可提高肌间脂肪和n-3PUFA的沉积,降低肌肉和肝中n-6/n-3的比值。在饲喂富含亚麻酸日粮的同时,添加酵母硒可提高血液中抗氧化酶活性和肝抗氧化基因的表达,但不能促进PUFAs在组织中沉积,因此,酵母硒可能不是促进饲喂紫苏籽日粮湖羊PUFA沉积最有效的方式,需进一步探究有效的抗氧化剂。
