Effect of Different Dietary Protein and Lipid Levels on the Growth, Body Composition, and Intestinal Digestive Enzyme Activities of Juvenile Yellow Drum Nibea albiflora (Richardson)
2018-08-28WANGLigaiHUShenyuLOUBaoCHENDongxingZHANWeiCHENRuiyiLIUFengandXUDongdong
WANG Ligai , HU Shenyu, LOU Bao , CHEN Dongxing, ZHAN WeiCHEN Ruiyi LIU Feng , and XU Dongdong
1) Zhejiang Marine Fisheries Research Institute, Zhejiang Provincial Key Laboratory of Mariculture & Enhancement,Zhoushan 316000, China
2) Marine and Fishery Research Institute, Zhejiang Ocean University, Zhoushan 316000, China
(Received September 20, 2017; revised November 21, 2017; accepted June 1, 2018)
© Ocean University of China, Science Press and Springer-Verlag GmbH Germany 2018
Abstract An 8-week feeding trial was conducted to determine the optimal dietary protein-to-lipid ratio for juvenile Nibea albiflora with an initial weight of (11.76 ± 0.20) g. Nine experimental diets containing different concentrations of protein (40%, 47%, or 54%) and lipids (5%, 9%, or 13%) in a 3 × 3 factorial experimental design were tested in triplicate groups of fish, while the protein-to-energy (P/E) ratios of the diets varied in the range of 19.74-28.32 mg kJ-1. Results showed that fish fed diets containing 9% or 13% lipids with 54% protein exhibited significantly higher weight gains and specific growth rates than those fed other diets. The feed conversion rate of fish fed the diet with 40% protein and 5% lipids was significantly poorer than that of fish fed other diets. The protein efficiency rate of fish fed diets with 5% lipids was significantly lower than that of fish fed 9% or 13% lipid diets. Carcass lipid and energy contents were positively correlated with dietary lipid level regardless of protein level. Fish fed a 54% protein diet showed the highest trypsin activity. The intestinal lipase activity of fish fed the diet containing 13% lipids was significantly higher than that of fish fed 5% or 9% lipid diets. These results demonstrate the high protein dietary requirements of N. albiflora. A diet containing 54% protein and 9%-13% lipids with a P/E ratio of 26.2-27.81 mg protein kJ-1 can be considered optimal for juvenile N. albiflora.
Key words Nibea albiflora; protein to lipid ratio; growth performance; feed utilization; digestive enzyme activities
1 Introduction
Protein is the most crucial and expensive nutrient affecting fish growth (Kim and Lee, 2005). Unfortunately,though increasing dietary protein intake up to a certain level can generally enhance fish production (Zhang et al.,2017), high dietary protein contents also proportionally increase feed costs (Lee et al., 2002), and the ammonia excreted after amino acid metabolism deteriorates water quality (Kim and Lee, 2005). Therefore, dietary protein levels must be optimized to meet the nutrition requirements on the one hand and the feed costs need to be reduced on the other hand (Rahimnejad et al., 2015).
Utilization of dietary protein by fish is related to protein level and non-protein energy sources (Yuan et al.,2010). Diets containing insufficient energy from nonprotein sources relative to the protein level may lead to protein catabolism to meet energy requirements at the cost of somatic growth (Sedgwick, 1979; Ward et al.,2003). Therefore, optimizing the protein-to-energy ratio(P/E) in feeds improves utilization of proteins for synthesizing body protein rather than allocating them for energy purposes (Lee et al., 2002). A number of studies have determined the optimal P/E ratios for cultured fish species,such as Japanese seabass Lateolabrax japonica, which is 25.9 mg protein kJ-1(Ai et al., 2004); Chinese sucker Myxocyprinus asiaticus, which is 29.22-31.43 mg protein kJ-1(Yuan et al., 2010); and hybrid grouper Epinephelus fuscoguttatus × E. lanceolatus, which is 23.9 mg protein kJ-1(Rahimnejad et al., 2015).
Protein utilization by fish can be improved by partially replacing the protein with lipids (NRC, 1993). Dietary lipids provide the essential fatty acids, phospholipids, and fat-soluble vitamins that are necessary for the proper functioning of physiological processes (Ghanawi et al.,2011). It also serves as an energy source for protein sparing (Mishra and Samantaray, 2004). The protein- sparing effect of lipids has been reported in common dentex Dentex dentex L. (Skalli et al., 2004), bagrid catfish Pseudobagrus fulvidraco (Kim and Lee, 2005), and Pacific blue-fin tuna Thunnus orientalis (Biawas et al., 2009). However, excess dietary lipid supplementation may cause growth retardation (Gonzalez-Felix et al., 2015) and even confer extreme metabolic burden to the liver (Jin et al.,2013). Therefore, dietary protein and lipid levels should be balanced when formulating diets to promote the maximum growth of fish (Zhang et al., 2017).
Yellow drum Nibea albiflora, Sciaenidae is an emerging commercially important marine species distributed in the coastal area of China, the Korean peninsula, and Japan’s southern coastal waters (Han et al., 2008). As culturing this fish mainly depends on chopped or minced trash fish as food, which is difficult to store and results in poor water quality, using formulated feeds to grow N.albiflora to market size would be more practical and efficient in terms of minimizing labor cost and mitigating adverse environmental impacts. Therefore, studies on N.albiflora nutrition should be carried out to develop a cost-effective and nutritionally balanced diet for this species. The protein (Lu et al., 2015), lipid (Wang et al.,2016), and vitamin C (Wang et al., 2017) requirements of N. albiflora have been well documented. However, the interactions between dietary protein and lipids and the optimal P/E ratio for this species remain unknown. The present study was conducted to determine the optimal dietary protein to lipid ratio for juvenile N. albiflora.
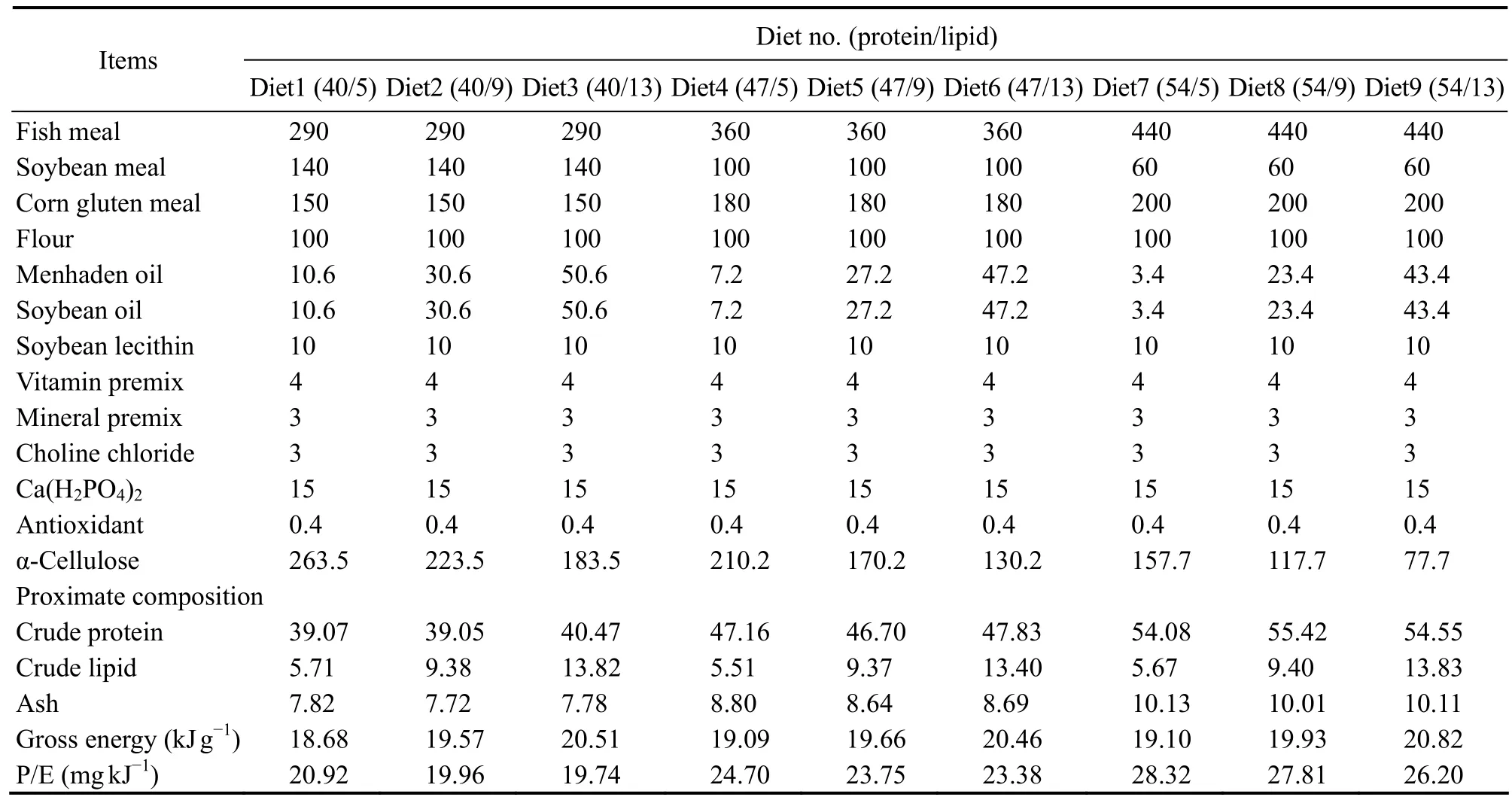
Table 1 Composition and nutrient contents of experimental diets (g kg-1 dry weight)
2 Materials and Methods
2.1 Experimental Diets
Nine experimental diets were formulated in a 3 × 3 factorial design to include three dietary protein levels (40%,47%, or 54%) and three dietary lipid levels (5%, 9%, or 13%) to produce P/E ratios in the range of 19.74-28.32 mg protein kJ-1(Table 1). Fish, soybean, and corn gluten meals were used as protein sources, and menhaden oil was used as the main lipid source (Table 1). All ingredients were purchased from Ningbo Tech-Bank Feed Co. Ltd.(Ningbo, China) and ground into a fine powder with a < 60 mesh particle size. The micro-components were mixed by progressive enlargement. Lipids and distilled water were added to the precombined dry ingredients and mixed to homogeneity in a Hobart-type mixer. Cold-extruded pellets with 3.0 and 5.0 mm in diameter were then produced.The pellet strands were broken into particles with uniform size with a granulating machine (G-250, machine factory of South China University of Technology, Guangzhou, China), steamed for 30 min at 90℃, and then air-dried to approximately 10% moisture. Finally the pellets were sealed in vacuum-packed bags and stored at -20℃ until use.
2.2 Animals and Feeding
Juvenile N. albiflora from XiShan Technology Island(Zhoushan, Zhejiang Province, China) were acclimated using a commercial diet for 2 weeks prior to the feeding trial. At the beginning of the experiment, 20 juvenile N.albiflora with the initial weight of 11.76 ± 0.20 g (mean ±standard deviation) and similar sizes were transferred to each of 27 cylindrical fiberglass tanks (500 L). The diets were assigned randomly to triplicate groups of fish. Each tank was provided with continuous water flow (2 L min-1)and uninterrupted aeration using air stones to maintain dissolved oxygen levels at or near saturation. All fish were fed to satiation twice daily at 08:00 and 17:00. During the experimental period, the water was maintained at 26-28℃ with pH 7.6-7.8 salinity 27.0 ± 1.00 g L-1, unionized ammonia nitrogen < 0.05 mg L-1, and dissolved oxygen > 6.0 mg L-1. All tanks were cleaned biweekly when the fish were removed to be weighed.
2.3 Sample Collection and Chemical Analysis
2.3.1 Sample collection
At the end of the 8-week feeding trial, the fish were fasted for 24 h. The total number and weights of fish in each tank were determined for evaluation of the final weight gain (WG), specific growth rate (SGR), feed conversion rate (FCR), protein efficiency rate (PER), and survival. Three fish were sampled randomly from each tank and anesthetized by MS-222, and then the body length,viscera, and liver weight were measured to calculate the viscerosomatic index (VSI), hepatosomatic index (HSI),and condition factor (CF). The mid-intestine was dissected from four fish in each tank and stored at -80℃ for further determination of intestinal digestive enzyme activity. An additional five fish were captured randomly from each tank for whole-body analysis.
2.3.2 Chemical analysis
Crude protein, lipid, moisture, and ash contents were determined from the fish diets and whole fish body according to the methods of the Association of Official Analytical Chemists (AOAC, 1995). Moisture content was determined after oven drying at 105℃ to a constant weight. Crude protein content (N × 6.25) was determined via the Kjeldahl method with a KjelFlex K-360 setup(Buchi, Flawil, Switzerland). Crude lipids were determined using a Soxtec System HT machine (Soxtec 2055;FOSS Tecator, Hillerød, Denmark). Ash content was determined by placing samples in a muffle furnace at 550℃for 8 h. Energy was determined by an adiabatic bomb calorimeter (OR2014, Shanghai, China).
2.4 Determining Digestive Enzyme Activities
2.4.1 Enzyme extraction
The mid-intestine was cleaned twice in pre-chilled(0-4℃) saline solution. The surface water was removed with filter paper, and the mid-intestine was weighed. The mid-intestine was homogenized in pre-cooled saline at a 1:9 mass/volume ratio, and the supernatant was collected after a centrifugation at 3000 × g for 20 min at 4℃ using a high-speed refrigerated centrifuge (5810R; Eppendorf,Hamburg, Germany).
2.4.2 Digestive enzyme activity
Intestinal trypsin activity was determined by the Folin-phenol reagent method (Huang et al., 2013) with some modifications. Briefly, 1 mL of the extract was preheated in a 37℃ water bath for 5 min, after which 2 mL of 0.5%casein solution was added to it as the substrate for 10 min at 37℃. Thereafter, 3 mL of 10% trichloroacetic acid was added to the solution. Then the solution was centrifuged at 4000 r min-1for 10 min to obtain the supernatant for the following analysis. A total of 5 mL of 0.55 mol L-1sodium carbonate and 1 mL of Folin-phenol reagent were added to 1 mL of the supernatant. The color of the solution was then developed for 10 min in a 37℃ water bath, and colorimetric determination was performed at a wavelength of 680 nm. In this experiment, 1 ml of deionized water was used as the blank. One unit of trypsin activity was defined as the amount of enzyme needed to produce 1 μg of tyrosine from the hydrolyzed casein per minute at 37℃.
Intestinal lipase and amylase activities were determined using commercial reagent kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) according to the instruction manuals of the manufacture. For intestinal lipase determination, triglycerides were incubated in Tris buffer at 37℃. Control reactions were run without the enzyme and substrate. Optical density was determined at a wavelength of 420 nm for 10 min. One unit of lipase was defined as 1 μmol substrate liberated by 1 g protein min-1at 37℃. For amylase activity determination, reaction systems were incubated at 37℃ for 7.5 min and stopped by iodine solution. The control was run without the enzyme. The free starch was determined at a wavelength of 660 nm,and one unit of amylase activity was defined as 10 mg carbohydrate released by 1 mg protein for 30 min at 37℃.
2.5 Calculations and Statistical Analyses
The following parameters were used to evaluate growth performance:
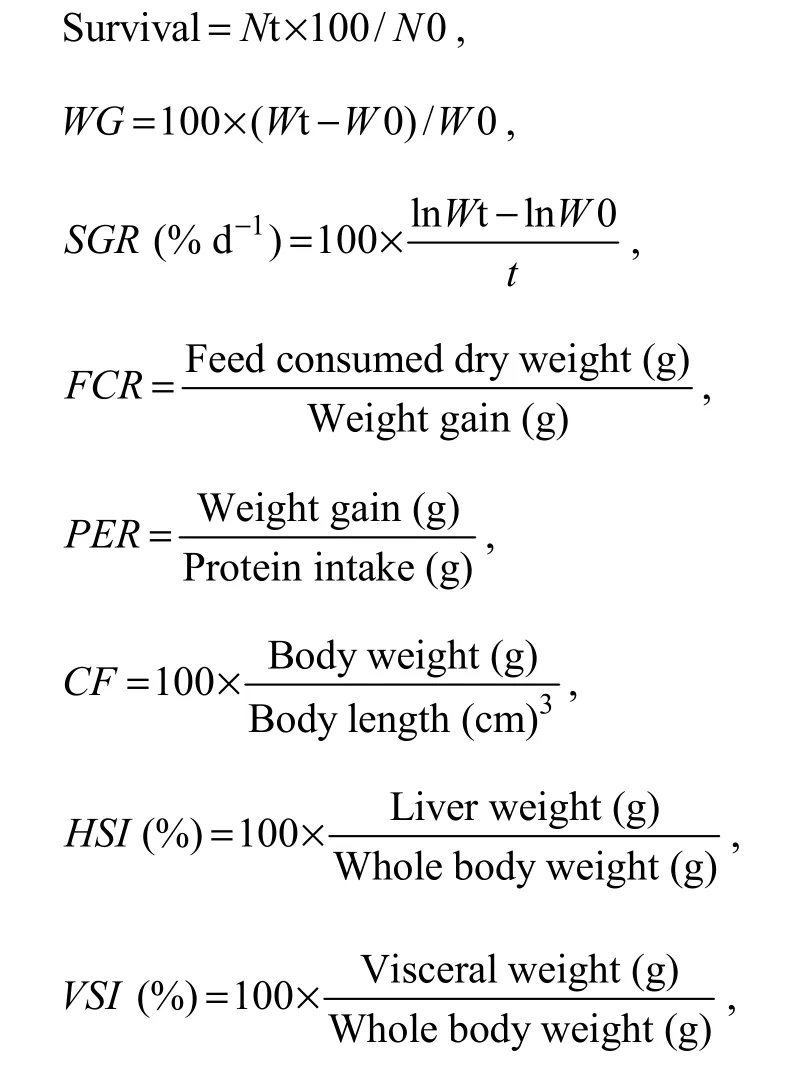
where Wt and W0 are the final and initial fish weights,respectively; WG, weight gain; SGR, specific growth rate;FCR, feed conversion rate; PER, Protein efficiency rate;Nt and N0 are the final and initial fish numbers in each tank, respectively; and t is the experimental duration in days.
To compare the effects of dietary treatments on the parameters tested, data were analyzed by two-way analysis of variance with three protein levels and three lipid levels as main factors. When a significant difference was detected in the main effect, Turkey’s multiple-range test was used to analyze the difference among treatments. The level of significance was set to P < 0.05. All statistical analyses were performed using SPSS 19.0 software (SPSS Inc., Chicago, IL, USA).
3 Results
3.1 Growth Performance and Feed Utilization
The WG, SGR, and FCR of yellow drum were significantly affected by dietary protein and lipid levels, and dietary lipid levels also significantly affected PER (Table 2). Fish fed a diet with 54% protein showed significantly higher WG and SGR values than those fed diets containing 40% or 47% protein, regardless of dietary lipid level.The FCR of fish fed a 40% protein diet was significantly higher than that of fish fed diets with 47% or 54% protein,regardless of dietary lipid level. Fish fed a diet with 5%lipids exhibited significantly lower WG, SGR, and PER values than those fed diets containing 9% or 13% lipids,regardless of dietary protein level. The FCR values of fish on diets with 9% or 13% lipids were significantly lower than those on diets with 5% lipids. However, no interactions between dietary protein and lipid levels were detected for growth performance or feed utilization.
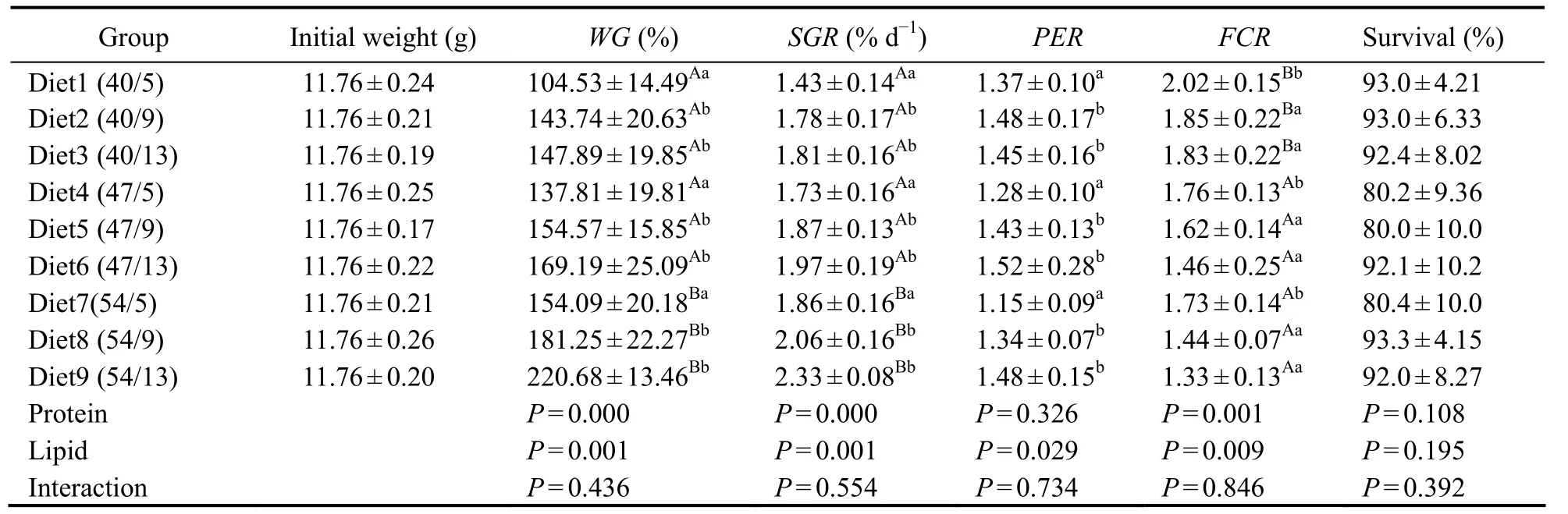
Table 2 Effects of dietary protein-to-lipid ratio on weight gain (WG), specific growth rate (SGR), protein efficiency rate (PER), feed conversion rate (FCR) and survival of juvenile Nibea albiflora
3.2 Morphological Indices
Fish fed diets with 9% or 13% lipids demonstrated a significantly higher VSI than those fed a diet with 5%lipids (Table 3). No interaction between protein and lipid levels was observed for VSI, and no significant differences in HSI and CF were detected among the treatments.
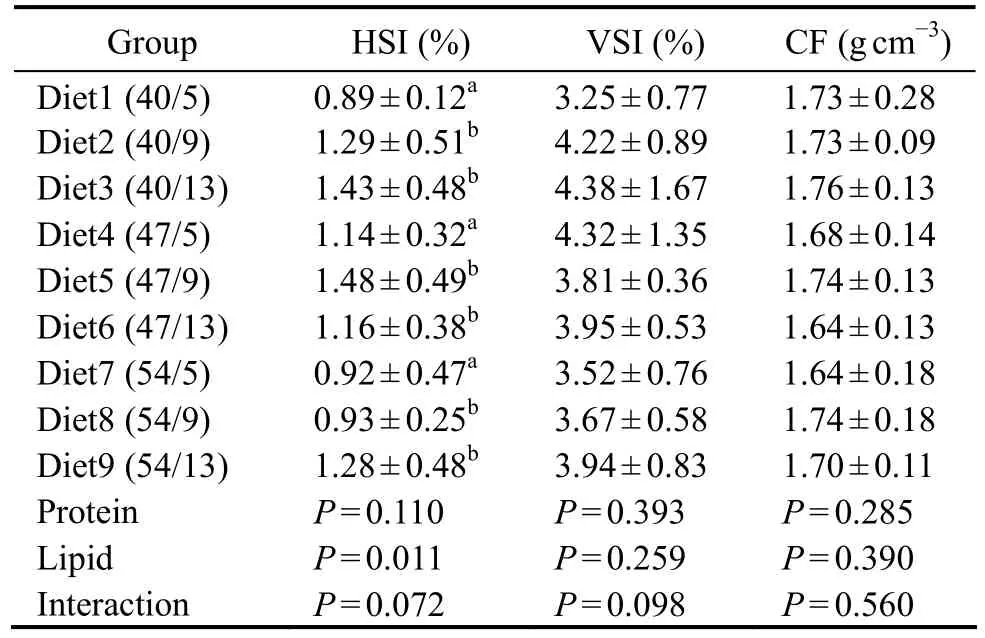
Table 3 Effects of dietary protein-to-lipid ratio on the body indices of juvenile N. albiflora
3.3 Body Composition
Dietary lipid levels significantly affected the whole body moisture, crude lipid, and energy contents (as shown in Table 4). Fish fed a diet containing 5% lipids revealed significantly higher moisture contents than those fed diets with 9% or 13% lipids. The crude lipid and energy contents of fish fed a 13% lipid diet were the highest among the groups examined. While no significant effects of dietary treatment on whole-body crude protein or ash contents were observed, an interaction between dietary protein and lipid levels was detected for energy content.This result suggests that lipids present a protein-sparing effect.
3.4 Digestive Enzyme Activities
The intestinal trypsin activities of yellow drum were significantly affected by dietary protein level (Table 5).Fish fed a diet with 54% protein showed significantly higher trypsin activities than those fed diets with 40% or 47% protein. The intestinal lipase activities of fish fed a diet with 13% lipids were also significantly higher than those of fish fed diets with 5% or 9% lipids. No significant difference in intestinal amylase activity was observed among the treatments.
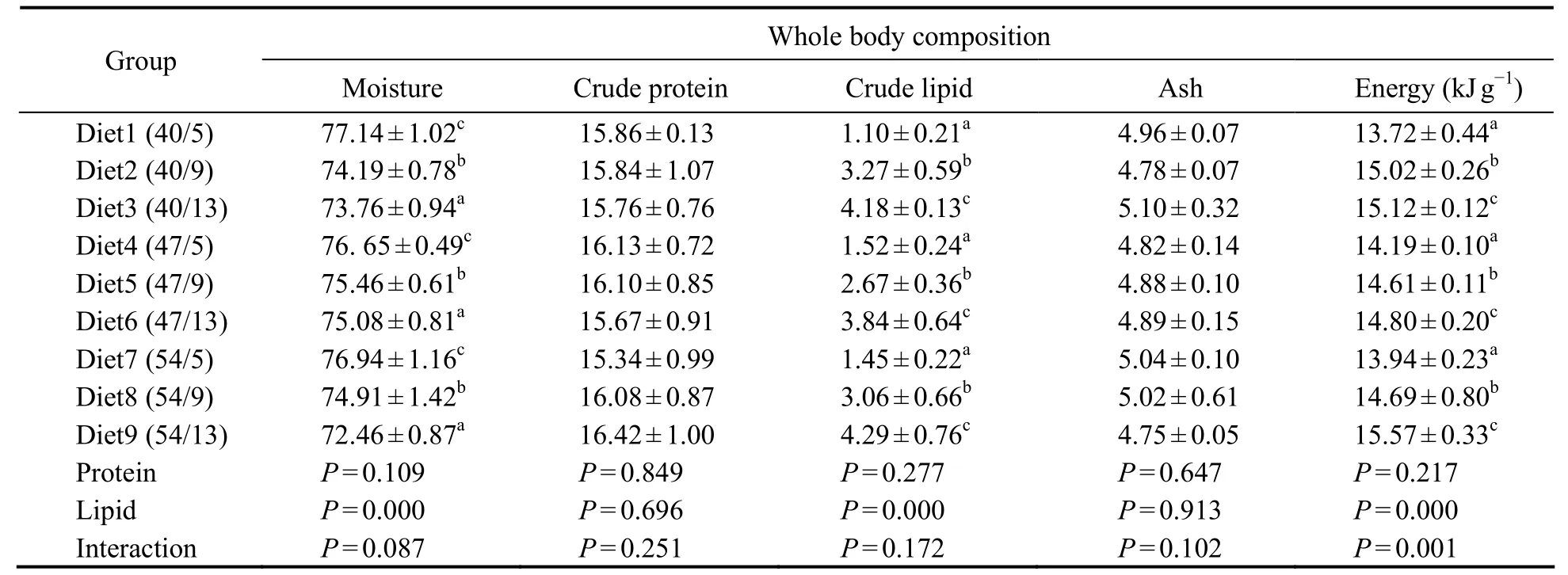
Table 4 Effects of dietary protein-to-lipid ratio on the whole-body composition of juvenile N. albiflora (% wet weight basis)
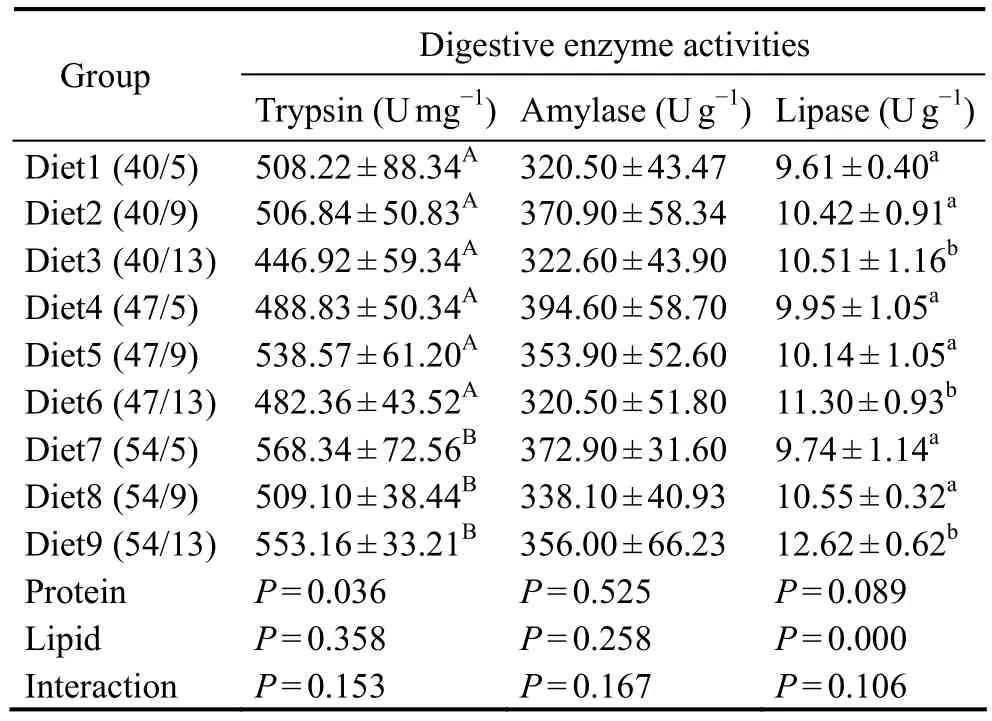
Table 5 Effects of dietary protein-to-lipid ratio on the intestinal digestive enzyme activities of juvenile N. albiflora
4 Discussion
The aim of this study is to determine the appropriate combination of dietary protein and lipids for optimal growth, efficient feed utilization, and improved survival of yellow drum (N. albiflora). In this study, the best WG and SGR were observed in fish fed diets containing 9% or 13% lipids with 54% protein, corresponding to a P/E ratio of 27.81 or 26.2 mg protein kJ-1, respectively. However, in view of efforts to conserve the cost of feeding, improve growth, and produce minimal pollution, a P/E ratio of 27.81 mg protein kJ-1can be considered optimal. This ratio is consistent with those reported previously for mutton snapper Lutjanus analis, which is 27.5-29.5 mg protein kJ-1(Watanabe and Eills, 2001), and Japanese seabass L. japonicas, which is 25.9 mg protein kJ-1(Ai et al.,2004). It is lower than that reported for seabass Lates calcarifer, which is 30.6 mg protein kJ-1(Catacutan and Coloso, 1995), and higher than that found for hybrid grouper E. fuscoguttatus × E. lanceolatus, which is 23.9 mg protein kJ-1(Rahimnejad et al., 2015). Maintaining a balanced P/E ratio results in efficient use of feed energy,enhances the rapid growth of aquaculture species, and promotes feed utilization (Winfree and Stiekley, 1981;Huo et al., 2014).
In this study, the optimal dietary protein and lipid levels for yellow drum based on WG and SGR were determined to be 54% protein and 9%-13% lipids. This result is similar to the findings in previous studies on humpback grouper Cromileptes altivelis, while the protein content is 53%-54% and lipids content is 10%-12%, respectively (Rachmansyah et al., 2005). In the present research, significant improvements in WG and SGR were observed as levels of dietary lipids increased at the same level of dietary protein. This result suggests that protein is utilized for growth rather than energy and confirms a protein-sparing effect, in agreement with previous studies (Skalli et al.,2004; Kim and Lee, 2005). Fish fed diets with 9% or 13%lipids demonstrated a significantly higher PER than those fed a 5% lipid diet, regardless of dietary protein level.This finding is a further evidence of a protein-sparing effect (Biawas et al., 2009). In contrast to this study, a protein-sparing effect was not found in Malaysian mahseer Tor tambroides (Ng et al., 2008) and white seabream Diplodus sargus (Ozorio et al., 2006).
VSI is an important index that directly affects fish yield(Wang et al., 2016; Wang et al., 2005). In the present study, fish fed diets containing 9% or 13% lipids revealed a higher VSI, which may be due to increased lipid retention. The result is similar to the findings in Pacific bluefin tuna T. orientalis (Biswas et al., 2009) and catfish Silurus asotus L. (Liu et al., 2013). While a very strong correlation between HSI and P/E ratio has been reported (Grisdale-Helland et al., 2008), no significant difference in HSI due to changes in dietary P/E ratio was observed in the present study.
Increasing dietary lipid levels may affect carcass qual-ity, mainly due to an increase in lipid deposition (Cowey,1993). Whole-body lipid and energy contents increased as dietary lipid levels increased, in accordance with previous studies on Japanese seabass L. japonicas (Ai et al., 2004),blunt snout bream Megalobrama amblycephala (Li et al.,2010), and brown-marbled grouper E. fuscoguttatus (Shapawi et al., 2014). Despite an increase in whole-body lipid contents of fish fed a diet with 13% lipids, the results of this study indicate that increasing dietary lipid levels from 5% to 13% enhanced growth performance and feed and protein utilization of yellow drum juveniles,suggesting that juvenile yellow drum can effectively utilize dietary lipids up to a certain level (Wang et al., 2016).Shearer (1994) reported that body lipid content is inversely correlated with moisture content, as was the case in the present study.
Digestive enzyme responsiveness to feeding and associated metabolic adjustments can be used in estimation of nutritionally effective diet formulations (Lundstedt et al.,2004). Trypsin is a major digestive enzyme that is positively correlated with dietary protein level (Lee et al., 1984)and growth rate (Lemieux et al., 1999). In the present study, enhancement of dietary protein level significantly enhanced intestinal trypsin activity, thereby suggesting that dietary protein levels can affect trypsin secretion levels and improve digestive tract proteolytic activities (Huang et al.,2013). This suggestion is supported by previous studies on yellow drum N. albiflora (Lu et al., 2015), rice field eel Monopterus albus (Ma et al., 2014), silver barb Puntius gonionotus (Mohanata et al., 2008), and spotted sorubim Pseudoplatystoma corruscans (Lundstedt et al., 2004).Aliyu-Paiko (2010) reported that many lipases are inducible. Carnivorous fish commonly consume fat-rich food,which can stimulate lipase activity in their digestive tract(Chakrabarti et al., 1995; Li et al., 2012; Ma et al., 2014).A positive correlation between intestinal lipase activity and dietary lipid level was observed in the present study.Improvements in lipase activity due to a high dietary lipid level reveal the importance of lipids as an energy source(Li et al., 2012; Divakaran et al., 1999).
In the present study, amylase was not significantly affected by dietary composition, possibly because of the stable dietary carbohydrate level, as shown in previous studies on rice field eel M. albus (Ma et al., 2014). However, Giri (2003) reported an increase in α-amylase activity in fish fed a diet with low protein levels.
5 Conclusions
The present study is the first one to report the effects of varying dietary protein and lipid levels on the growth,feed utilization, and intestinal digestive enzyme activities of yellow drum. A protein-sparing effect was observed,and intestinal trypsin and lipase activities were significantly enhanced by the supplementation of dietary protein and lipids. Based on the WG of yellow drum, a diet containing 54% protein and 9%-13% lipids with a P/E ratio of 26.2-27.81 mg protein kJ-1is recommended to achieve the maximum growth of juvenile yellow drum under the present experimental conditions. Our results indicate that N. albiflora is a species with high protein requirements.
Acknowledgements
This research was supported by grants from the National Natural Science Foundation of China (No. 4147 6127), the Science and Technology Planning Project of Zhejiang Province (No. 2016F50038), the Science and Technology Planning Project of ZhouShan City (No.2015C31010), and the Scientific Research Foundation of Zhejiang Ocean Universtiy (No. 2014Q1434).
杂志排行

Journal of Ocean University of China的其它文章
- Establishing Gene Delivery Systems Based on Small-Sized Chitosan Nanoparticles
- Methylation Status of the Follistatin Gene at Different Development Stages of Japanese Flounder(Paralichthys olivaceus)
- Taxonomic Clarification of A Well-Known Pathogenic Scuticociliate, Miamiensis avidus Thompson &Moewus, 1964 (Ciliophora, Scuticociliatia)
- Structural Variation Analysis of Mutated Nannochloropsis oceanica Caused by Zeocin Through Genome Re-Sequencing
- A Comparative Study on Hydrodynamic Performance of Double Deflector Rectangular Cambered Otter Board
- Characterization of Polysaccharides Extracted from a Cultivated Brown Alga Costaria costata During the Harvest Period