杂草对草甘膦抗性机制及治理对策
2018-08-24周欣欣黄兆峰魏守辉黄红娟张朝贤
周欣欣,黄兆峰,魏守辉,黄红娟,张朝贤
(1.农业部农药检定所,北京 100125;2.中国农业科学院植物保护研究所,北京 100193)
1 引言
草甘膦是由美国孟山都公司开发的广谱灭生性、内吸传导型除草剂,主要用来防除一年生及多年生恶性杂草。自1974年商品化以来,一直是最主要的除草剂品种(张朝贤等,2011)。草甘膦的作用机制是抑制植物体内5-烯醇式丙酮酸-3-磷酸合成酶(5-enolpyruvylshikimate-3-phosphate synthase,EPSPS)的活性,阻碍芳香族氨基酸的生物合成,导致莽草酸大量积累,进而扰乱正常的氮代谢致使植物死亡。草甘膦具有低毒、低残留等优点,简便高效的除草方式给人类带来了巨大经济效益。但是,由于草甘膦的长期大量使用,也使得抗草甘膦杂草不断产生与发展。截止到2018年3月,全球共发现41种抗草甘膦杂草(表1)(Heap,2018)。抗草甘膦杂草的出现与发展严重威胁到草甘膦的使用和农业生产。生产上,农民通常加大草甘膦的使用剂量来防除抗性杂草,不仅增加了成本,影响农药减量使用,还造成了环境污染。因此,杂草对草甘膦的抗药性机制研究及其防除已经成为全球关注的重要问题。

表1 抗草甘膦杂草分布
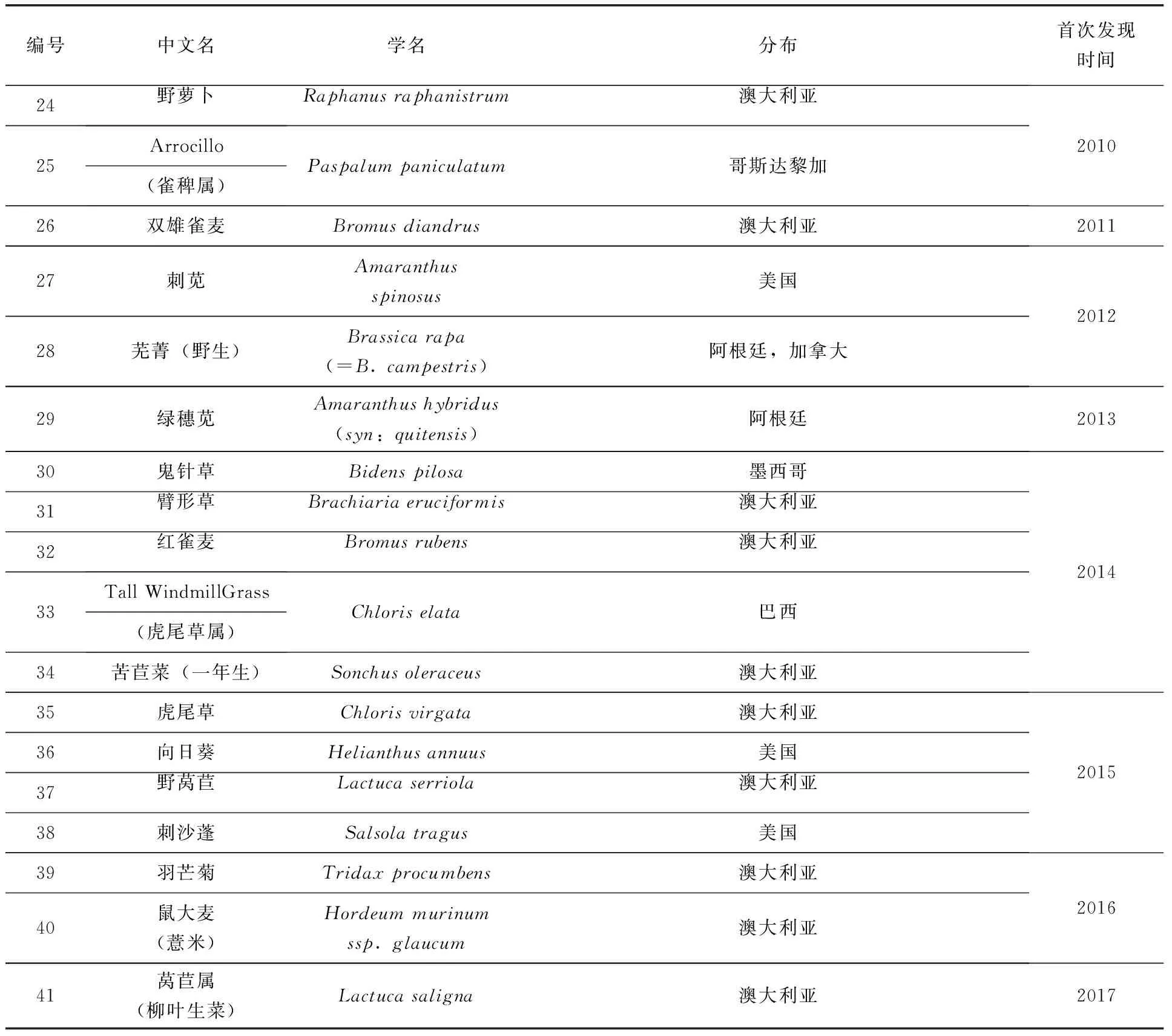
续表
2 抗草甘膦杂草研究进展
草甘膦具有独特的作用方式及代谢机制,使人们一度认为在田间不可能出现抗草甘膦杂草。然而,1996年在澳大利亚发现的瑞士黑麦草(Loliumrigidum)对草甘膦表现出了高水平的抗药性,标志着抗草甘膦杂草的出现(Pratleyetal.,1999)。在马来西亚,草甘膦连续10年使用后,牛筋草(Eleusineindica)对草甘膦的抗性达8~12倍。2000年在美国抗草甘膦转基因大豆田中发现,小蓬草(Conyzacanadensis)对草甘膦的抗性提高了8~13倍(Vangessel,2001)。2005年,Culpepper等对美国乔治亚州传统的棉花、花生和大豆田怀疑对草甘膦具有抗性的长芒苋(Amaranthuspalmeri)进行了抗性鉴定,结果显示其抗性生物型的I50 值是敏感生物型I50值的12倍(Culpcpper et a1.,2006)。同年,在美国还发现了抗草甘膦的具瘤苋(Amaranthusrudis)和三裂叶豚草 (Ambrosiatrifida),在巴西和阿根廷分别发现了抗草甘膦的猩猩草(Euphorbiaheterophyll)和假高梁(Sorghumhalepense)(Heap,2018)。2007年在澳大利亚发现了抗草甘膦的光头稗(Echinochloacolona)。耐草甘膦转基因作物种植面积的逐年提高加速了草甘膦的需求。此外,少耕免耕农业系统的需求增长,大大拓宽了草甘膦的施用范围。草甘膦的大面积推广使用将加速抗草甘膦杂草的发生和发展。
在我国,2005年抗草甘膦的小蓬草在浙江被发现和报道(Conyzacanadensis)(Songetal.,2005),2012年抗草甘膦牛筋草在我国南方作物田中被发现(杨彩宏等,2012)。最近,在湖南省部分施用草甘膦的棉田也发现抗性牛筋草种群(刘慧雄等,2015)。这表明草甘膦在我国果园等农作物种植区的长期应用已经使抗性问题逐渐产生。未来随着草甘膦应用范围的扩大,抗草甘膦杂草产生的几率将进一步升高。
3 杂草对草甘膦的抗药性机制
大多数除草剂通过抑制植株生长发育中的关键酶而能杀死杂草,杂草体内这些受除草剂抑制的酶被定义为除草剂的靶标酶。杂草对除草剂的抗药性通常情况可分为靶标抗药性和非靶标抗药性。靶标抗药性是指靶标酶基因发生突变或多拷贝,使除草剂不能有效抑制靶标酶(Leeetal.,2015;Hanetal.,2016;Duhouxetal.,2017)。非靶标抗药性主要是杂草通过代谢酶(如细胞色素酶P450、谷胱甘肽-S-转移酶和葡萄糖醛酸转移酶)代谢解毒作用使除草剂失活(Yu & Powles,2014;Iwakamietal.,2014)。就杂草对草甘膦的抗药性机制而言,主要包括以下几点:
3.1 靶标抗药性机制
3.1.1 靶标基因发生改变 草甘膦的靶标EPSPS基因关键的核苷酸被取代,导致表达的氨基酸类型改变,从而引起EPSPS酶活性发生变化,致使草甘膦与靶标酶结合能力降低,最终导致草甘膦的药效降低。到目前为止,已报道的抗草甘膦杂草EPSPS突变主要位于106位,如:马来西亚的牛筋草、智利的多花黑麦草在EPSPS 106位氨基酸由脯氨酸变为丝氨酸或苏氨酸(Baersonetal.,2002;Fideletal.,2012)。部分抗草甘膦杂草也伴随其他位点的突变,如在澳大利亚发现的抗草甘膦瑞士黑麦草EPSPS同时存在2个突变位点,一是106位脯氨酸突变为苏氨酸,二是301位脯氨酸突变为丝氨酸(Simarmataetal.,2008)。对草甘膦具有高抗性的牛筋草种群EPSPS同时存在102位和106位(T102I+P106S)2个氨基酸突变(Yuetal.,2015;陈景超等,2016)。
3.1.2 靶标酶的过量表达 杂草EPSPS基因表达量升高,植株体内产生过量的EPSPS酶能缓解草甘膦对其限制,从而对草甘膦产生抗药性。Yuan等研究发现,抗草甘膦的狗甘草经草甘膦处理后,EPSPS在转录水平上迅速增加,过量的EPSPS解除了草甘膦对其限制并提供足够的酶活性来满足代谢的需要,从而增加了对草甘膦的抗药性(Yuan et al.,2002)。但是有些抗草甘膦杂草是由于EPSPS基因在染色体上成倍复增导致的。Gaines等对抗草甘膦长芒苋(Amaranthuspalmeri)进行了抗性机理的研究,发现由于EPSPS基因在多个染色体上成倍扩增,在转录时增加了EPSPS的表达,从而导致长芒苋对草甘膦的抗药性(Gainesetal.,2010)。这一新的抗性机制在《PNAS》杂志刊登后,研究者在意大利黑麦草(Loliumperennessp. multiflorum)、地肤(Kochiascoparia)也发现了这些杂草对草甘膦抗药性是由于EPSPS基因大量复增所导致(Salasetal.,2012; Wiersmaetal.,2015)。
3.2 非靶标抗药性机制
3.2.1 草甘膦的输导与分布 在对小蓬草的草甘膦抗性机制研究中发现,减少草甘膦向分生组织输导是其抗草甘膦的主要原因(Fengetal.,2004)。随后在黑麦草中也发现了同样的抗性机制(Wakelinetal.,2004)。Culpepper等用高浓度的草甘膦对抗草甘膦长芒苋茎叶喷雾处理后发现,叶腋处的生长点可以继续生长(Culpcpperetal.,2006)。Fidel在研究抗草甘膦多花黑麦草(LoliummultiflorumLam.)时发现,抗药性生物型不仅在EPSPS106位发生突变,并且抗性生物型比敏感型在植株根和叶片积累的草甘膦更少(Fideletal.,2012)。
3.2.2 草甘膦的隔离 部分抗草甘膦杂草可以通过将吸收到的草甘膦迅速在液泡中封存,从而明显降低草甘膦的毒副作用。Ge等通过31P核磁共振在小飞蓬上观察到了草甘膦的快速封存化,这说明杂草可以通过将除草剂隔离的办法来实现一种在生物化学层面的抗性(Geetal.,2010)。
4 抗草甘膦杂草的治理
资料显示,草甘膦抗性杂草发生,主要是重复而单一地使用草甘膦,尤其是那些以种植抗草甘膦转基因作物为主的地区。因此,有针对性地进行除草技术的改进,是有效防除杂草,降低杂草抗性的必经之路。
4.1 除草剂混用 不同作用方式的除草剂轮换或混合使用是防除抗药性杂草的有效方法之一。对于草甘膦防除效果不佳或产生抗性的杂草如苣荬菜、田旋花、小飞蓬等,可以与苯氧羧酸类除草剂2,4-D丁酯等混用来提高防除效果。也可以与其他类型的灭生性除草剂如草铵膦等轮换使用这样既可以提高防除效果。草甘膦也可以与一些土壤处理活性较高的除草剂混用,如酰胺类,磺酰脲类等,这样既能延长除草持效期又能降低重复施药成本。
另外,草甘膦与茎叶处理除草剂的混用。如在生产上将草甘膦与苯氧羧酸类除草剂2,4-滴丁酯、2甲4氯等混用,既可提高防效,又可解决难防杂草问题。其次是与土壤处理除草剂的配合使用。由于草甘膦只有在杂草出土后进行茎叶处理,不能防除未出土杂草,因此,在喷施草甘膦的同时,将一些土壤处理除草剂(如酰胺类、磺酰胺类、咪唑啉酮类、三嗪类等)进行封杀,既能延长除草剂的持效期,减少施药量,延缓抗性的产生。
4.2 提高施药技术 现在生产上仍然存在一些不科学的施药行为如:错施、误施、漏施、滥施等。如何能够真正做到因草施药,见草施药;如何落实因草情而确定施药剂量,考虑生态和经济阚值而不盲目追求高防效是目前生产上亟待解决的重要课题。开发推广高性能喷雾器械,提高喷雾质量等提高施药相关技术对草甘膦的施用年限具有重要意义。
4.3 加强管理,发挥优势 由于草甘膦在土壤中活性低,见效慢,人们很容易忽视其污染问题而加大施用剂量,尤其是在水田边、铁路、果园等非耕地施用,结果造成水体的严重污染。欧洲一些国家已经检测发现,草甘膦对地表水、地下水和雨水都造成一定程度的污染。如何加强草甘膦的管理和正确指导科学地使用,充分发挥其优势,避免抗性杂草的发展和环境污染,以延长其使用年限,确保草甘膦能更好的为农业发展服务,这是当前的一个重要任务。
4.4 利用新技术-RNA干扰 RNA干扰(RNA interference,RNAi)是指由双链RNA分子引起的序列特异性靶标基因mRNA降解,从而导致内源或外源靶基因沉默的机制。2013年2月在澳大利亚举行的“全球除草剂抗性挑战”国际会议上,孟山都公司利用RNA干扰技术进行了草甘膦杂草治理的探索和研究,结果表明,通过导入特异的双链RNA,可以使抗性长芒苋失去对草甘膦的抗性。但该技术只能解决靶标酶改变的抗性机制杂草,且在田间是否可应用,暂无相关研究。
5 小结
尽管目前研究者将杂草抗草甘膦机制分为靶标抗性和非靶标抗性,但不少研究者坚持认为EPSPS活性增高是产生抗性的主要原因。同时EPSPS其他位点的突变以及这些突变位点与抗性的关系有待于进一步挖掘和验证。相信随着研究的深入,人们将揭示出更加深入合理的抗性机制。
草甘膦在我国已使用40余年,长期使用草甘膦,抗药性杂草产生与发展的风险在迅速增加。抗草甘膦的小飞蓬和牛筋草已在我国浙江和广东发现(Songetal.,2005)。为防止抗草甘膦杂草的产生与发展,在抗药性杂草的治理中,应重视除草剂使用的多样性,倡导不同作用方式除草剂的轮用和交替使用,以延缓抗药性杂草的发生;引入绿色发展理念——重视耕作方式和作物品种的多样性,充分发挥农艺措施、生态调控等手段对杂草的控制作用;同时应加强对抗药性杂草的监测,建立高效、系统的抗性杂草治理策略。
