套袋对翠冠果实外果皮褐斑形成结构及生理影响
2018-08-23施春晖王晓庆张学英
施春晖,王晓庆,张学英,骆 军*
(1.上海市农业科学院 林木果树研究所,上海 201403;2.上海市设施园艺重点实验室,上海 201403)
砂梨果皮的颜色可分为3种类型:全锈色、半锈色和绿色[1-2]。翠冠是典型半锈色中国砂梨品种,具有产量高、质量好的特点。但外果皮颜色不均一,覆盖了一层不均匀的木栓层,而消费者并不喜欢不美观的果实外观。翠冠的栽培面积已占全国梨面积的8%左右[3]。南方砂梨品种优,但由于外果皮锈斑而严重影响其外观品质,从而阻碍了南方半锈砂梨向全国大面积的推广。因此,提升半锈砂梨外观品质是当务之急。
具有锈色的外果皮是木栓层覆盖在绿色或黄色底色上导致的。在梨果的实发育过程中,果皮角质层和表皮细胞破损后积累的木栓层形成梨果皮绣斑(褐色),木质素(Lignin)是木栓层的重要成分[4-5]。因此木质素的积累与梨果皮绣斑(褐色)的形成有关[6]。木质素是一类苯丙烷类聚合物,是植物细胞壁次生加厚过程中形成的重要物质,木质素参与木栓层的形成。木栓质被发现是一种普遍存在的不溶性高分子聚酯由两聚一聚酚类和脂肪族结构域(SPPD为木栓质聚酚域;SPAD为木栓质聚脂肪族结构域)[7-9]。对锈色果皮和绿色果皮突变体之间的生化物质的比较分析也表明:木质素、纤维素、半纤维素的含量可高于绿色果皮的[10-12]。PAL酶是催化其代谢反应第一步的酶,PAL的表达直接影响整个合成过程,当PAL含量增加后,木质素含量明显增多;当PAL酶活性下降时,植物中的木质素含量下降[13-15]。CCoAOMT在植物木质素合成中对G木质素特意调控,研究均表明:CCoAOMT的表达活性与木质素含量密切相关。不同的下游分支代途径代谢增强都会引起苯丙氨酸代谢的加剧,木质素合成途径就是苯丙氨酸下游的代谢途径之一。张云华等[16]研究木质素合成相关的PPO和POD酶活性与果皮褐色明显相关。
目前,生产上利用套袋技术可消除或减轻锈斑。张勃等通过对黄冠梨套袋研究果实锈斑随套袋时期推迟而减少,另外,不同的果袋对果实表面锈斑也有影响,对黄冠梨的研究表明,套双层袋果实的锈斑指数与不套袋相近,套单层袋和单条纹纸袋袋内温度相对较低,相对湿度也低于双层袋和对照,抑制了果皮木栓形成层的活动,减轻了果实锈斑的形成和发育[17]。玻璃纸和牛皮纸促进了果点和果锈的形成,而蜡纸显著地抑制果点的木栓化,报纸袋提早果面木栓化,促进果锈的形成[5,18]。目前,关于套袋对梨果发育期到成熟期木质素积累及木质素合成关键酶活性变化影响的研究较少。
本研究以半锈砂梨为翠冠为试材,不套袋处理为对照,通过对套袋梨果实的褐斑指数、木质素含量、木质素代谢关键酶活性的测定,比较分析了套袋后外果皮褐斑的发生机理,深入探讨了果皮褐色的形成机理,为今后提高砂梨果实的外观品质提供理论依据。
1 材料与方法
1.1 供试材料
2016年在上海金冠蜜梨种植园(30.92° N,121.47° E)中开展套袋试验。土壤为粘壤土,年平均气温15.7 ℃,年平均降水量1178.2 mm。半锈色中国砂梨12年生翠冠果实为试材,盛花后5 d套袋,观测取盛花后10 d到成熟期(盛花后120 d)分为10个阶段,从果实发育早期到中期,果实样品每10 d取样1次,近成熟期果实样品每20 d取样1次。
1.2 方法
选择12年树龄生长健壮、树势一致,负载量相近的翠冠树8株,标记挂牌。分别在每棵树上随机套白色小林袋60个,不套袋的果实作为对照,重复3次。果实发育早期(花后10~80 d),木栓层变化显著,每10 d取样1次;花后80~120 d,每20 d取样1次,当天带回实验室并削下果实中部外果皮液氮冷冻后,磨成细粉后待测定。
1.2.1 果锈指数统计 参照Jones等[19]的方法略有改进,由于砂梨锈斑的覆盖率高,即根据果锈程度将果实分为8级:无锈果为0级,果锈面积1%~5%为1级,6%~10%为2级,11%~15%为3级,16%~20%为4级,21%~40%为5级,41%~60%为6级,61%~80%为7级,81%~100%为8级。
果锈指数=[∑(各级代表数值×该级果数)]/(总果数×最重级代表数值)×100。
1.2.2 外果皮褐斑的显微结构观察 盛花后10~120 d利用体视显微镜(放大10倍)观察翠冠梨的外果皮木栓层形成过程并拍照。
1.2.3 木质素含量 采用Morrison[20]的木质素测定方法测定木质素含量。
1.2.4 木质素合成代谢关键酶活性测定 苯丙氨酸解氨酶(Phenylalanine ammonia-lyase,PAL)的提取和活性测定参照Dyckmans等[21]的方法测定,稍做修改。果肉4.0 g,加10 mL含5 mmol/L的巯基乙醇的硼酸缓冲液、0.5 g PVP,加少量石英砂研磨成匀浆,在4 ℃下,10000 r/min离心15 min,上清液为粗酶液。活性测定的反应体系为4 mL,含1 mL酶液、1 mL 0.02 mol/L苯丙氨酸、2 mL蒸馏水,以不加底物为对照,40 ℃水浴1 h后,在波长290 nm处测定溶液吸光度值,以每小时吸光度变化0.001所需酶量为1个酶活性单位。
4-香豆酸辅酶A连接酶活性的测定(4-hydroxycinnamate co-A ligase,4CL)参照Knobloch等[22]的方法进行测定。
肉桂酸羟化酶(cinnamate-4-hydroxylase,C4H)酶活性测定参考陈雷等[23]的方法进行测定。
肉桂醇脱氢酶(Cinnamyl alcohol dehydrogenase,CAD)活性的肉桂醇脱氢酶(CAD)活性测定参照Goffner等[24]方法进行测定。
过氧化物酶(Peroxidase,POD)活性的测定参照Cai等[25]的方法进行测定。样品取样1 g,加8 mL(0.05 mol/L、pH 5.5)PBS,冰浴研磨,在4 ℃下,10000 r/min离心10 min,取上清液定容于25 mL容量瓶中。反应体系:2.9 mL(0.05 mol/L、pH 5.5)PBS,1.0 mL 2% H2O2,1.0 mL 0.05 mol/L愈创木酚和0.1 mL酶液,对照管加0.1 mL死酶液。在波长470 nm下比色。以OD470变化1.0为1个酶活性单位,酶活性表示为ΔOD470/(g·min)。
1.3 数据处理
数据整理和处理采用Excel 2003软件,数据相关性分析用SPSS 17.0数据处理系统。
2 结果与分析
2.1 套袋对翠冠梨外果皮结构影响的动态变化
外果皮的果锈实质是木栓层的沉着,在盛花后10 d对果实套袋,整个果实发育进程中木栓层的形成速度明显减缓。由图1可知,从外观上看,套袋和不套袋果实发育到成熟期的动态观测中,盛花后50 d即果实快速膨大期之前,外果皮颜色均为绿色,木栓层的发生时间存在明显差异。盛花后50 d套袋果实外果皮除气孔处褐色木栓化外果皮较为洁净,直到盛花后60 d套袋果实外果皮才有明显的褐色木栓层,而不套袋的果实在盛花后30 d外果皮上已近出现点状锈斑。盛花后70 d,果实进入快速膨大期,外果皮木栓层面积随着果实迅速发育而迅速扩大,但此时套袋的果实仍以绿色为主,而不套袋的果实外果皮绝大多数被木栓层覆盖,表现为浅褐色。随后木栓层随着发育进程而迅速扩大,套袋果实外果皮木栓层覆盖始终较慢,且木栓层颜色变深较慢,直到成熟期外果皮虽有锈斑但颜色浅且仍可见大面积的绿色底色,相比较不套袋的果实盛花后80 d,其外果皮基本被褐色木栓层覆盖且颜色逐渐加深,成熟时深褐色锈斑几乎完全覆盖外果皮。
利用显微镜观察套袋和不套袋果实外果皮结构的变化,由图1可知,木栓层微观结构的变化早于外观的变化。果点发生在幼果发育初期且早于锈斑的发生,套袋后果点的形成进程也会显著减缓,不套袋的果实外果皮在盛花后10 d就进入了皮孔期,即气孔明显下限与表皮细胞内形成圆锥结构,圆锥体内部处于亚表皮细胞恢复分裂能力形成木栓层。套袋后20 d,果实外果皮才逐渐进入皮孔期,而此时不套袋的外果皮已进入了果点形成期,即木栓化了的填充细胞呈黄褐色,且撑破周围的表皮,逐渐填满空腔,形成果点。盛花30 d后,套袋与不套袋的外果皮均进入果点膨大期,不套袋果实果点不仅迅速膨大,而且外果皮角质膜出现龟裂少量表皮细胞发生木栓化。套袋后果实盛花后50 d,外果皮才出现角质膜龟裂表皮细胞木栓化的现象,此后外果皮表皮细胞继续木栓化,但木栓化程度及速度始终慢于不套袋的果实。盛花后70 d,果实进入快速膨大期,外果皮木栓化面积随着果实发育角质膜龟裂迅速扩大,并且木栓化细胞逐渐加深,不同的是套袋果实外果皮果点木栓化颜色较不套袋的颜色深,但表皮细胞的木栓化程度轻于不套袋,而且发展速度缓慢。盛花后120 d,成熟期的套袋果实外果皮的木栓化程度与不套袋果实盛花后50~60 d的相近。
2.2 套袋对翠冠梨外果皮褐斑指数的影响
果实锈斑指数是衡量外果皮锈斑覆盖程度的标志性参数。由图2可知,翠冠果实外果皮随着果实发育,褐斑指数呈上升的趋势,幼果期上升较缓慢,盛花后70 d果实迅速膨大期,果实锈斑指数显著上升。套袋果实与不套袋(对照)果实相比,果实全发育期果实锈斑指数趋势一致,但上升幅度显著低于不套袋(对照)果实。与不套袋对照相比,套袋果实在幼果期即盛花后10~20 d,果锈未形成期果锈指数差异不显著,盛花后30 d不套袋果实外果皮角质膜的龟裂表皮细胞开始发生木栓化,果锈指数高于套袋11.67%。盛花后60 d果实迅速膨大期前,套袋果实果锈指数低于不套袋(对照)果实的果锈指数16.9%。盛花后70 d果实迅速膨大期,套袋果实果锈指数上升显著低于不套袋(对照),这一时期也是它们之间果锈差异最大的时期,套袋果实果锈指数较不套袋(对照)低41.67%,盛花后100 d果实逐渐进入成熟期,两者仍差异显著,但幅度小于盛花后70 d。

图1 翠冠梨果实发育期套袋对外果皮褐斑影响的表观及显微观测
2.3 套袋对翠冠梨外果皮木质素含量的影响
木质素是木栓层木栓质聚酚域SPPD的主成分,如图3所示,盛花后10~120 d,翠冠外果皮木质素的含量随着果实发育呈上升的趋势,盛花后各个时期,套袋后果实外果皮的木质素含量均低于不套袋的果实,盛花后60 d果实成熟之前,套袋果实外果皮木质素略低于不套袋(对照)木质素含量,且差异不显著;盛花后70~80 d果实迅速膨大期,套袋与不套袋(对照)外果皮木质素含量差异显著,不套袋(对照)外果皮在这2个时期木质素含量高于套袋外果皮的15.56%和35.01%,盛花后100 d,两者差距逐渐缩小,但套袋后外果皮木质素含量仍低于不套袋(对照)外果皮的。

图2 翠冠梨果实发育期套袋对外果皮褐斑指数的影响

图3 翠冠梨果实发育期套袋对外果皮木质素含量的影响
2.4 套袋对翠冠梨外果皮木质素生物合成关键酶活性的影响
2.4.1 套袋对翠冠梨外果皮苯丙氨酸解氨酶(PAL)活性的影响 由图4可知,翠冠梨果实外果皮PAL,酶活性在整个生育期呈先上升后下降的趋势。盛花后10~120 d套袋后果实外果皮PAL活性低于不套袋果实的PAL活性,盛花后30~40 d套袋与不套袋(对照)果实外果皮PAL酶活性均达到最大,此后开始下降,但两者的差异在盛花后50 d左右最为显著,不套袋(对照)果实外果皮PAL活性是套袋外果皮PAL酶活性的2.13倍。幼果发育期和果实成熟期两者的酶活性差异最小。
2.4.2 套袋对翠冠梨外果皮4-香豆酸辅酶A连接酶(4CL)活性的影响 翠冠梨外果皮的4CL酶活性在整个果实生育期呈双S曲线,在盛花后30~40 d显著上升后显著下降,盛花后70 d后小幅上升后下降。由图5可知,在整个生育期,套袋后的果实外果皮4CL活性低于不套袋(对照)。盛花后10~20 d,套袋梨果实外果皮4CL酶活性和不套袋(对照)差异不显著;盛花后30~60 d,幼果发育期即果实迅速膨大前套袋梨果实外果皮4CL酶活性比不套袋(对照)低71.43%~75.00%;仅在盛花后70 d和120 d,套袋后果实外果皮4CL活性高于不套袋果实,但盛花后80 d套袋比不套袋(对照)外果皮4CL酶活性低120%,且差异极显著。

图4 翠冠梨果实发育期套袋对外果皮PAL酶活性的影响

图5 翠冠梨果实发育期套袋对外果皮4CL酶活性的影响
2.4.3 套袋对翠冠梨外果皮肉桂酸-4-羟基化酶(C4H)活性的影响 C4H酶活性在翠冠梨果实外果皮内的活性变化见图6,随着果实的发育呈先上升后下降的趋势,下降后的酶活性缓慢降低,套袋与不套袋(对照)在盛花后30~40 d显著上升后急剧下降,在这段时期套袋果实的外果皮C4H酶活性显著低于不套袋(对照),分别比对照低43.19%和57.45%。其他各时期套袋和不套袋(对照)外果皮C4H酶活性相比时而略高时而略低,但差异不显著。
2.4.4 套袋对翠冠梨外果皮肉桂醇脱氢酶(CAD)活性的影响 由图7可知,随着幼果发育翠冠梨果实外果皮CAD酶活性变化趋势平缓,仅在盛花后50 d有小幅的上升,直到盛花后70 d果实进入迅速膨大期后CAD活性出现显著上升,盛花后80 d的CAD活性达到最大,盛花后100 d趋于成熟期,CAD酶活性出现下降趋势。在整个果实发育期,套袋梨果实外果皮CAD活性始终低于不套袋果实外果皮的CAD活性,其中在两者在盛花后30、70、100 d差值大于10 U/g FW。

图6 翠冠梨果实发育期套袋对外果皮C4H酶活性的影响

图7 翠冠梨果实发育期套袋对外果皮CAD酶活性的影响
2.4.5 套袋对翠冠梨外果皮过氧化物酶(POD)活性的影响 翠冠梨果实外果皮中的POD酶活性在整个果实生育期呈双S曲线。由图8可知,在盛花后10~60 d显著上升后显著下降,盛花后40 d达到第1个小高峰;盛花后60~120 d先显著上升后显著下降。在刚套袋后果实POD酶活性显著低于不套袋(对照)果实外果皮,此后幼果发育期两者差异不显著,盛花后70~120 d即果实膨大期至果实成熟期套袋的翠冠梨果实外果皮的POD活性极显著低于不套袋的果实。
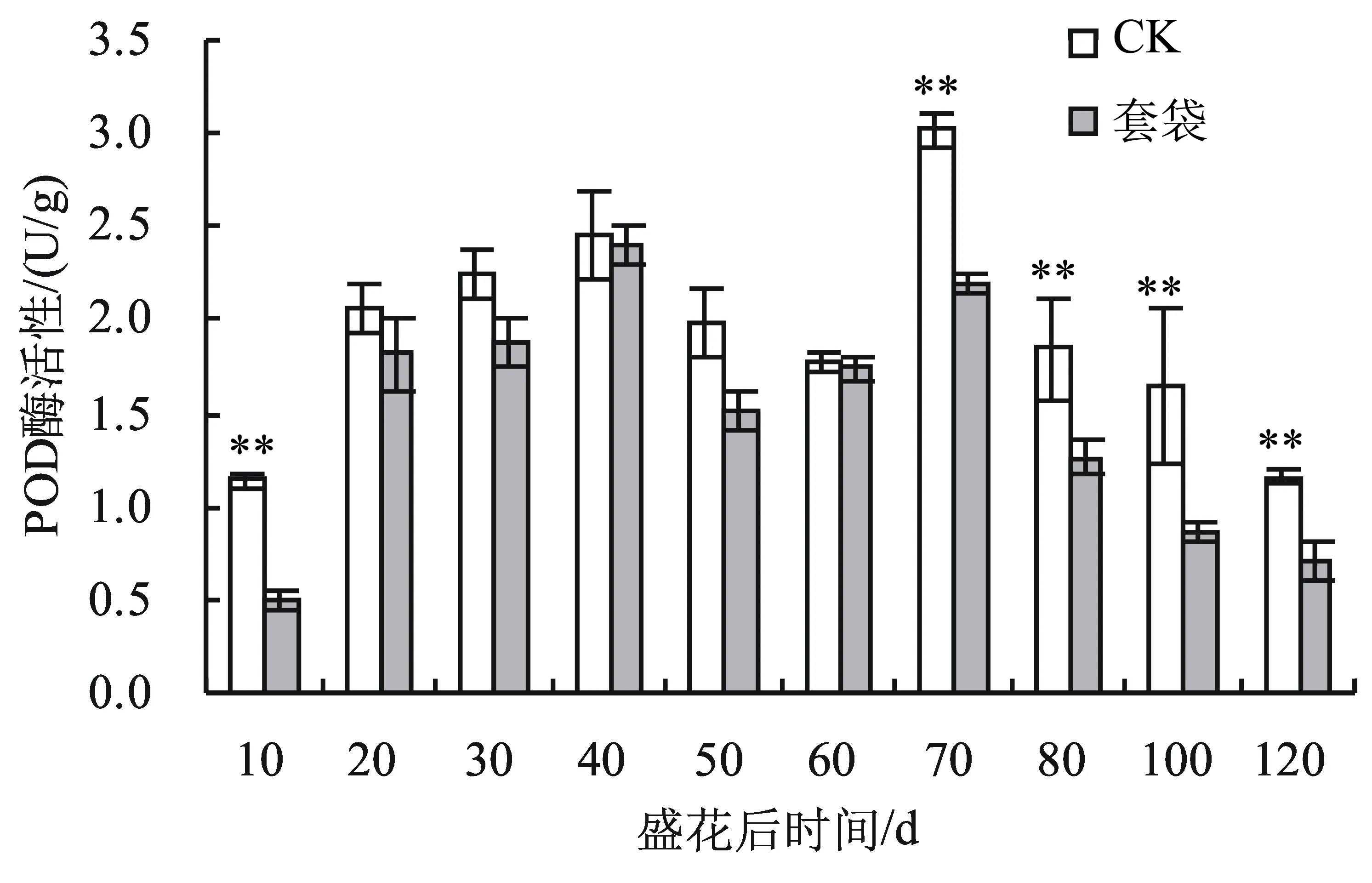
图8 翠冠梨果实发育期套袋对外果皮POD酶活性的影响
2.4.6 翠冠梨外果皮木质素含量与木质素合成代谢关键酶的相关性分析 由表1可知,翠冠梨外果皮木质素含量与PAL酶、4CL酶、C4H酶、CAD酶、POD酶均呈正相关性。不套袋的翠冠外果皮木质素含量与CAD呈显著正相关;PAL酶与4CL酶,C4H酶呈显著正相关;4CL酶与C4H酶呈极显著正相关。套袋后翠冠梨外果皮中PAL酶与4CL酶呈极显著正相关;4CL酶与C4H酶呈显著正相关。

表1 翠冠外果皮木质素含量与木质素代谢关键酶活性相关性分析
注:**表示极显著相关;*表示显著相关。对角线右上角数据为翠冠不套袋(对照);左下角数据为翠冠套袋。
3 讨论
翠冠梨属于典型的半锈色砂梨,它的果锈实质是果实外果皮覆盖的一层木栓层,其形成原因主要是由于外界雨水等不利果实发育的环境影响,导致外果皮最外层角质膜失去保护能力,下层表皮细胞木栓化逐渐覆盖于外果皮形成褐斑[26-29]。本试验结果表明,不套袋翠冠果实外果皮在盛花后30 d就形成一定面积的锈斑,这一结果比王慧等[30]认为盛花后49 d后出现不规则锈斑斑点要早很多,这可能是试验环境差异导致其形成时间不一致。在盛花后10 d对翠冠幼果套白色油蜡纸制作的小林袋可有效延缓外果皮木栓层的发生及发展进程,套袋后显微观测盛花后50 d果实外果皮才出现表皮细胞木栓化,盛花后60 d肉眼才能观测到褐斑小斑点。
果点是早期幼果皮孔处表皮细胞木栓化的结果,是否与果皮锈斑有关目前争议较大。本试验结果表明,翠冠梨果点与外果皮锈斑不存在因果关系,这一结果与施泽彬[31]的研究结果一致。套袋后翠冠果实锈斑明显减轻但盛花后50 d果点的发展程度比不套袋对照强烈。
木栓质是由木栓聚紛结构域(SPPD)和木栓聚脂结构域(SPAD)组成的大分子构成,其中聚酚结构域主要由羟基阿魏酸及其衍生物以类似木质素单体形成木质素的方式共价连接而成。因此,木质素生物合成在锈斑形成中起至关重要的作用。李晓峰等在砀山梨褐皮芽变中发现,锈皮芽变外果皮木质素合成代谢关键酶PAL、4CL、CAD以及POD酶活性均高于正常砀山梨外果皮。本试验研究结果表明:不套袋翠冠褐斑严重,它的外果皮中木质素单体含量以及木质素合成代谢关键酶活性,包括PAL酶、4CL酶、CAD酶、C4H酶的活性普遍高于套袋翠冠果实外果皮。然而有研究报道CAD酶基因得到抑制可能有助于褐色锈斑在外果皮上沉着[11-12],而一些研究表明:CAD活性减低会导致木质素前体物质减少[32-33]。Moura等[34]认为SPPD中木质素前体的含量降低可能进一步促进细胞壁松动和膨胀利于锈斑的积累,最近笔者对半锈砂梨结构研究中发现,即使外果皮角质膜破裂痕较大,但如果表皮细胞木质化程度低,外果皮锈斑较少(文章待发表)。推测果实锈斑不仅仅与机械膨胀挤压力有关,可能与自身防御调控能力有关。
在相关分析中,套袋与不套袋(对照)结果显示,两者的木质素与合成代谢酶的正相关性不一致,未完全达到显著水平,木质素生物合成PAL酶、C4H酶、4CL酶相关性较为显著。毕竟翠冠果实锈斑形成机制是一个复杂的进程,木质素合成仅是锈斑形成的其中一个环节,整个褐斑的形成过程尚需进一步研究。
