温室番茄对增施不同浓度CO2的光合响应
2018-08-16张振花袁宏霞郑金英邢国明
张振花,袁宏霞,刘 洋,李 靖,郑金英,孙 胜,邢国明
(山西农业大学园艺学院/山西省设施蔬菜提质增效协同创新中心,山西太谷 030801)
CO2作为植物进行光合作用的重要原料之一,也是植物气孔反应、光合作用及光合同化物分配的调节者,参与植物体内一系列的生理代谢反应,其浓度过高或过低都会影响植物的光合作用、呼吸作用以及蒸腾作用三大生理过程[1–2]。其中,光合作用为作物的生长发育提供必要的营养物质,是其获得产量的重要途径,尤其作物花期后的光合性能对其产量的形成具有重要作用,光合作用的强弱直接影响作物产量的高低[3]。因此,CO2浓度是植株生长发育的重要限制因素[4]。
随着工业化和城市化进程的加快,大气中CO2浓度虽将达到400 μmol/mol[5],仍远低于园艺作物光合作用的最适浓度。而温室栽培中,作物栽培密度较大,群体光合作用旺盛,但在其日光温室冬春季生产中,尤其是每年12月至翌年2月,温室往往由于保温压力而很少通风,甚至全天封闭,CO2得不
到及时的补充,导致温室内CO2常处于匮乏状态[6],严重影响作物的生长发育和光合作用[7]。因此在温室作物栽培过程中增施CO2是促进光合生理特性和提
高产量的重要途径之一。目前,不少学者研究表明,增施CO2处理的黄瓜株高、茎粗、净光合速率均显著高于未增施CO2的处理[8–9];CO2浓度倍增可显著提高辣椒和黄瓜幼苗净光合速率、显著降低蒸腾速率和气孔导度[10–11]。其中,CO2浓度升高能够增强作物的光合作用[12],促进作物叶片的生长和发育,一个重要的原因是CO2浓度升高会对作物叶片气孔的密度、导度和运动等方面带来较明显的影响[13–14]。而且,于作物不同生长时期增施CO2对其生长发育的影响也有所不同,小麦拔节期、孕穗期、抽穗期及灌浆期对增施CO2的光合生理响应表现出很大差异[15],CO2浓度升高对辣椒开花期和结果期光合作用的影响也有所差异[16]。
番茄 (Solanum lycopersicum Miller) 作为典型的喜温性蔬菜,是我国北方日光温室广泛栽培的蔬菜作物之一。虽然有研究发现,设施内增施CO2能促进番茄的生长发育、提高产量及改善番茄品质[8,17]。但是,目前关于温室番茄不同生育期对不同浓度CO2的光合生理响应的研究尚未见报道。本研究设置了4 个 CO2梯度,即 (400 ± 15)、(600 ± 20)、(800 ±25)、(1000 ± 30) μmol/mol,其中以大气 CO2浓度(400 ± 25) μmol/mol为对照,研究不同 CO2浓度对温室番茄不同生育期光合生理特性的影响,以明确温室番茄各生育期适宜生长的CO2浓度,为增施CO2有效促进番茄光合作用并提高产量的生产实践提供理论依据和技术支持。
1 材料与方法
1.1 供试材料
试验材料购买于山西省介休市兴海种苗公司,为培育40 d的番茄种苗,品种为‘兴海12号’。
1.2 试验方法
试验于2016年11月至2017年4月在山西农业大学园艺学院园艺站日光温室中进行。温室用塑料膜隔出四个独立的隔间,每个隔间为一个处理,每个隔间面积为 52 m2(6.5 m × 8 m)。待幼苗长至四叶一心时 (2016年11月3日),选取生长势一致的幼苗,分别定植于不同的隔间内。每个隔间栽培6畦,每畦为一个小区,每个小区为一个重复。每个小区定植30株,分为2行,宽行距90 cm,窄行距50 cm,株距40 cm。缓苗一周后,开始进行CO2施肥处理,于2017年4月底拉秧。以液态CO2钢瓶为碳源,施入的 CO2浓度分别为 (600 ± 20)、(800 ± 25)及 (1000 ± 30) μmol/mol ,分别记作 C1、C2、C3,以大气 CO2浓度 (400 ± 15) μmol/mol (C0) 为对照。CO2释放控制系统采用GMM220传感器 (芬兰VAISALA公司) 和邯郸冀南新区盛炎电子科技有限公司的自动控制系统,通过调整CO2钢瓶上的流量计控制气体流速和循流风机均匀施入。施肥时间为晴天 9:00~11:00,14:00~16:00,即每天增施 4 h,开膜放风与阴雨雪天不施用。
1.3 生育期确定
2016年11月3日番茄幼苗定植,2017年4月底拉秧,定植后田间生育期将近6个月。指标测定生育期 (田间生育期) 标准如下:
苗期:从定植到第一穗花序第一朵花现大蕾(2016.11.03~2016.11.24)。
开花期:从第一朵花现大蕾到第一朵花坐果(2016.11.25~2016.12.09)。
幼果期:从第一花序坐果到第一花序果实膨大前期 (2016.12.10~2017.01.07)。
成熟期:从第一花序果实成熟到采收末期(2017.02.05~2017.04.30)。
1.4 指标测定与方法
1.4.1 光合色素含量的测定 于番茄各生育期,每个处理有6个重复,每个重复分别随机选取1株生长势一致的植株,取生长点以下第3或4片完全展开的叶片后立即装入取样袋内,用冰壶带回实验室测定番茄叶片叶绿素a、叶绿素b和类胡萝卜素。测定方法采用80%丙酮浸提法[18]。
1.4.2 光合特性的测定 同上,每个处理有 6个重复,每个重复随机选择1株番茄植株,用美国LICOR公司的LI-6400便携式光合仪,选取植株生长点以下第3或4片完全展开叶测定其净光合速率(Pn)、气孔导度 (Gs)、胞间 CO2浓度 (Ci)、蒸腾速率(Tr) 等光合参数及光响应曲线,并计算叶片水分利用率WUE = Pn/Tr。测定时采用红蓝光LED叶室,叶室温度设定为25℃,气流速度设为500 μmol/s;光合参数测定时光合有效辐射强度 (PAR) 统一设置为1000 μmol/(m2·s),光响应曲线测定时 PAR 设为1800、1600、1400、1200、1000、800、600、400、200、150、100、50、0 μmol/(m2·s),测定时间为晴天9:00~11:00、14:00~16:00,其中,根据幼果期生长阶段的实际日出情况,测定时间调整为晴天9:30~11:30、14:00~16:00。
1.4.3 产量的测定 自收获期 (2017 年2月5日) 开始,每隔4~5 d采收一次成熟度一致的番茄,直至拉秧期 (2017年4月30日) 采收结束。
1.5 数据分析
采用 Microsoft Excel 2010 和 DPS 9.50 软件对试验数据进行统计分析,用LSD法进行差异显著性检验 (α = 0.05)。
利用经典的Farquhar模型进行光响应拟合。
Pn-PAR 曲线拟合:Pn =[PAR × AQY + Pmax −SQRT((AQY × PAR + Pmax)2− 4 × AQY × Pmax ×PAR × K)] /(2 × K) − Rd
式中,Pn:净光合速率;PAR:光合有效辐射;AQY:表观量子效率;Pmax:最大净光合速率;K:曲角;Rd:暗呼吸速率。
2 结果与分析
2.1 不同浓度CO2施肥对番茄各时期光合色素含量的影响
由图1可知,随着CO2浓度的升高,C3处理叶片叶绿素a含量在苗期、开花期和幼果期与对照C0处理相比差异达到显著水平 (P<0.05),成熟期的叶绿素a含量差异虽未达到显著,但C1、C2和C3处理与对照C0相比分别增加了8.0%、8.4%、28.9%。与对照相比,C3处理的叶绿素b含量在苗期、开花期、幼果期及成熟期差异显著,分别增加了17.5%、24.9%、43.9%、18.0%;随着CO2浓度的升高,类胡萝卜素含量在幼果期显著提高。相同的CO2浓度处理下,叶绿素a和叶绿素b含量在成熟期较其他生育期有所降低,类胡萝卜素含量在成熟期较开花期和幼果期也有所降低,但高于苗期。
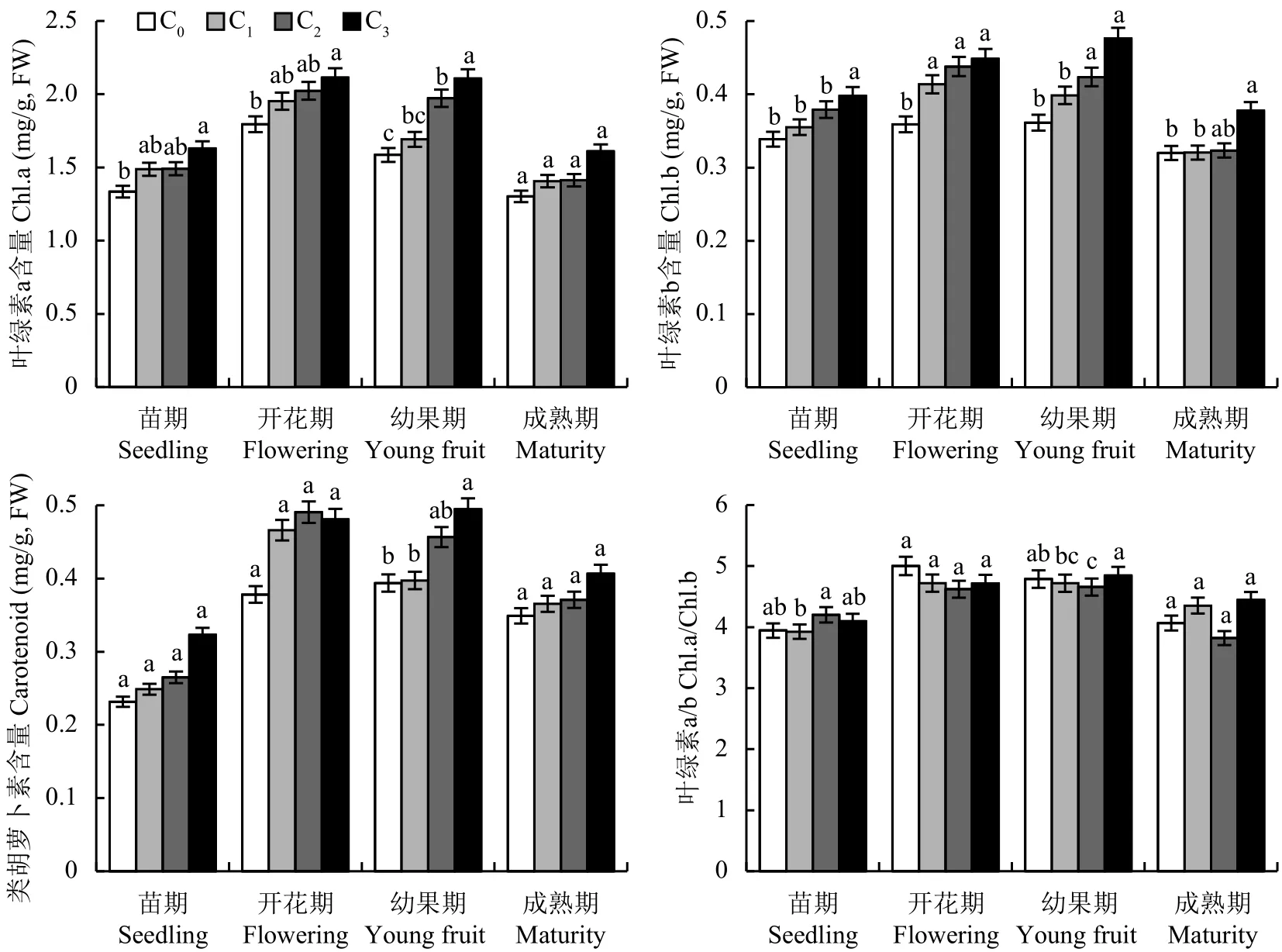
图1 不同CO2浓度下番茄不同生育期叶片的光合色素含量Fig.1 Content of photosynthetic pigments in tomato leaves of different growth stage under different CO2 concentrations[注(Note):C0—(400 ± 15) μmol/mol; C1—(600 ± 20) μmol/mol; C2—(800 ± 25) μmol/mol; C3—(1000 ± 30) μmol/mol.柱上不同字母表示处理间差异达 5% 显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
在开花期和幼果期,不同CO2浓度处理之间,叶绿素b的增幅大于叶绿素a的增幅,即随着CO2浓度的升高,虽然叶绿素a与叶绿素b含量有所提高,但叶绿素a与叶绿素b的比值 (chl.a/chl.b) 却随之减小,且差异性不显著。
2.2 不同CO2浓度施肥对番茄光合特性的影响
2.2.1 不同 CO2浓度对番茄叶片净光合速率 (Pn) 的影响 随着CO2浓度的升高,番茄叶片净光合速率 (Pn)在苗期、开花期、幼果期及成熟期均随之显著增加(图2)。与对照处理C0相比,C1、C2及C3处理的叶片Pn在苗期分别增加15.35%、23.40%、29.35%,开花期分别增加9.60%、23.69%、25.28%,幼果期分别增加21.92%、47.94%、52.18%,成熟期分别增加20.96%、49.46%、66.97%。相同CO2浓度下,C0处理下苗期和开花期的Pn大于幼果期和成熟期;增施CO2处理后,C2与C3处理下的番茄叶片Pn在幼果期与成熟期反而较苗期与开花期增势明显。
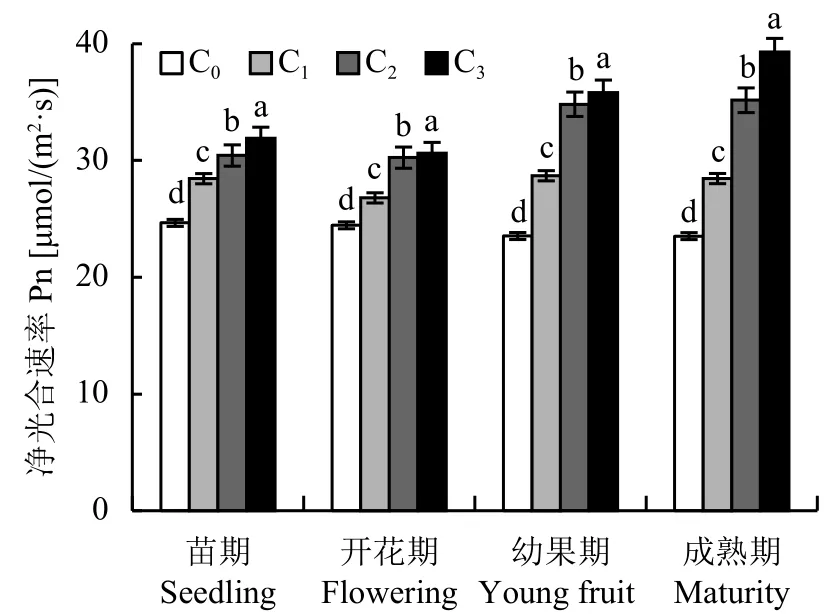
图2 不同CO2浓度下番茄不同生育期净光合速率的变化Fig.2 Photosynthetic rate of tomato of different growth stage under different CO2 concentrations[注(Note):C0—(400 ± 15) μmol/mol; C1—(600 ± 20) μmol/mol;C2—(800 ± 25) μmol/mol; C3—(1000 ± 30) μmol/mol.柱上不同字母表示处理间差异达 5% 显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
2.2.2 不同 CO2浓度施肥对番茄叶片气孔导度 (Gs) 的影响 从图3可以看出,C1、C2与C3处理的番茄叶片气孔导度 (Gs) 在苗期、开花期、幼果期及成熟期均显著低于对照处理C0,其中,C3处理降幅最大,C2处理次之。相同CO2处理时,从苗期到成熟期,对照C0下成熟期Gs虽大于苗期、开花期和幼果期,但差异未达到显著水平;而增施CO2后,C1与C2处理下各生育期Gs无明显变化,C3处理下苗期与幼果期的Gs显著低于开花期与成熟期。
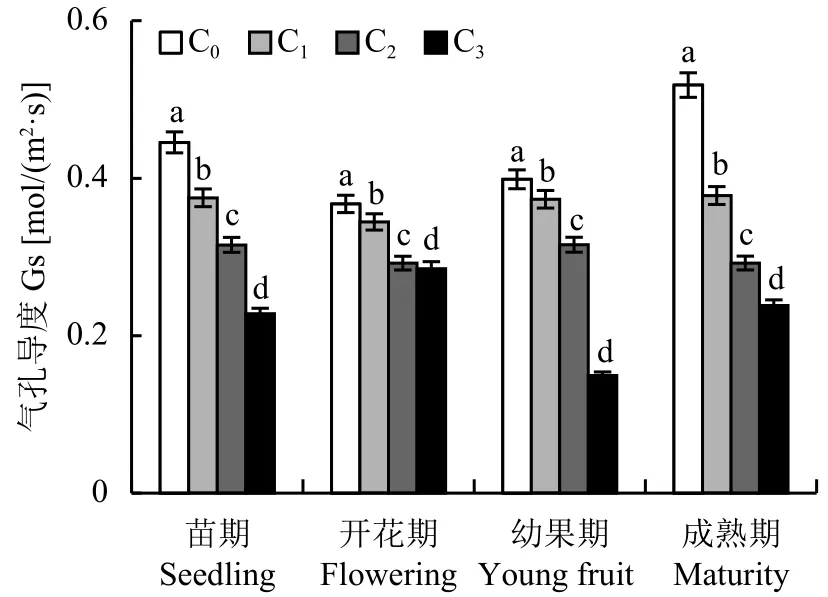
图3 不同CO2浓度下番茄不同生育期气孔导度的变化Fig.3 Stomatal conductance of tomato of different growth stage under different CO2 concentrations[注(Note):C0—(400 ± 15) μmol/mol; C1—(600 ± 20) μmol/mol;C2—(800 ± 25) μmol/mol; C3—(1000 ± 30) μmol/mol.柱上不同字母表示处理间差异达 5% 显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
2.2.3 不同 CO2浓度施肥对番茄叶片蒸腾速率 (Tr) 的影响 图3、图4结果表明,蒸腾速率的高低与气孔导度的大小密切相关,随着CO2浓度的升高,番茄叶片蒸腾速率 (Tr) 于各生育期均不同程度地显著降低 (图4)。不同CO2浓度处理下,生殖生长期的Tr较苗期下降幅度大,即增施CO2对苗期影响不大,而且,C2与C3处理的Tr下降幅度较C1处理明显。另外,相同CO2浓度时,C1处理下除了成熟期叶片Tr高于其它生育期,从苗期到幼果期均呈下降趋势,即成熟期 > 苗期 > 开花期 > 幼果期。
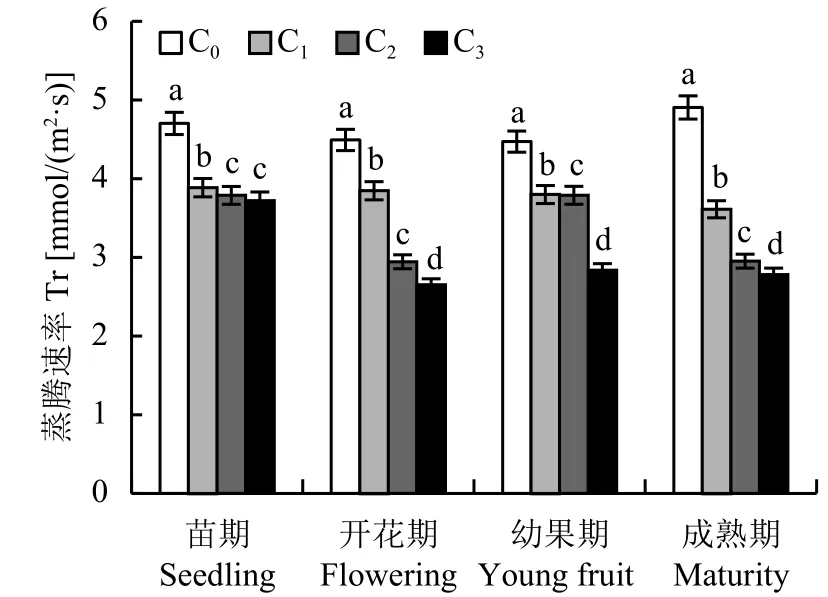
图4 不同CO2浓度下番茄不同生育期蒸腾速率的变化Fig.4 Transpiration rate of tomato of different growth stage under different CO2 concentrations[注(Note):C0—(400 ± 15) μmol/mol; C1—(600 ± 20) μmol/mol;C2—(800 ± 25) μmol/mol; C3—(1000 ± 30) μmol/mol.柱上不同字母表示处理间差异达 5% 显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
2.2.4 不同CO2浓度施肥对番茄叶片水分利用率的影响 由图5可知,随着CO2浓度的升高,番茄叶片水分利用率 (WUE) 在开花期与成熟期均随之显著增加,增施CO2对苗期影响较小,其增幅明显小于开花期、幼果期和成熟期。而且,从苗期到成熟期,C3处理对WUE影响最大,C2处理次之。CO2浓度不变时,对照C0下成熟期的WUE低于其它时期;增施CO2后,从苗期到成熟期,相同CO2浓度下的WUE呈不同程度的增加趋势,即增施CO2可提高番茄叶片WUE。
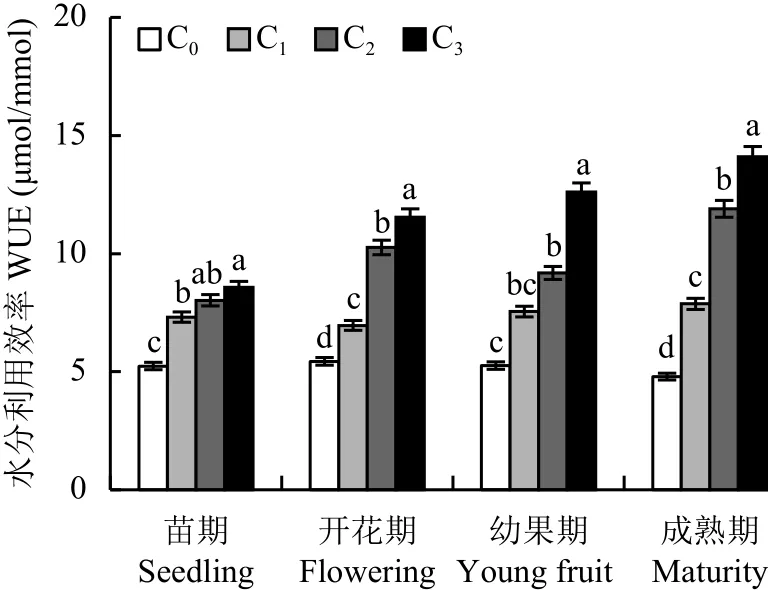
图5 不同CO2浓度下番茄不同生育期水分利用率的变化Fig.5 Water use efficiencies (WUE) of tomato at different growth stages under different CO2 concentrations[注(Note):C0—(400 ± 15) μmol/mol; C1—(600 ± 20) μmol/mol;C2—(800 ± 25) μmol/mol; C3—(1000 ± 30) μmol/mol.柱上不同字母表示处理间差异达 5% 显著水平 Different letters above the bars mean significant difference among treatments at the 5% level.]
2.2.5 不同CO2浓度施肥对番茄叶片胞间CO2浓度(Ci) 的影响 由表1可知,不同浓度CO2处理下,番茄叶片胞间CO2浓度 (Ci) 在苗期、开花期、幼果期及成熟期差异性均达到显著水平,并且随着CO2浓度的增高而增大。与对照C0处理相比,C1、C2与C3处理下番茄叶片Ci在苗期分别增加35.2%、123.5%、209.2%;在开花期分别增加56.5%、120.7%、208.3%;在幼果期分别增加90.4%、120.2%、137.1%;在成熟期分别增加51.7%、160.9%、214.8%。CO2浓度相同时,从苗期到成熟期,C0处理下Ci逐渐减小,增施CO2处理后Ci降低的情况有所改善。
2.3 不同浓度的CO2施肥对番茄光响应曲线特征参数的影响
由表2可知,光补偿点随着CO2浓度的升高而减小,光饱和点、最大净光合速率及表观量子效率均随着CO2浓度的升高而增大。与对照处理C0相比较,C3处理的光补偿点在苗期、开花期及成熟期分别显著降低25%、31.25%、27.27%,在幼果期无显著差异;光饱和点和最大净光合速率在苗期差异不显著,但在开花期、幼果期及成熟期,与对照C0相比,C3处理的光饱和点和最大净光合速率均呈显著增加。说明随着CO2浓度的升高,番茄叶片对弱光和强光的利用能力得到增强。

表1 不同CO2浓度下番茄不同生育期胞间CO2浓度 (μmol/mol)Table1 Intercellular CO2 concentration of tomato of different growth stage under different CO2 concentrations
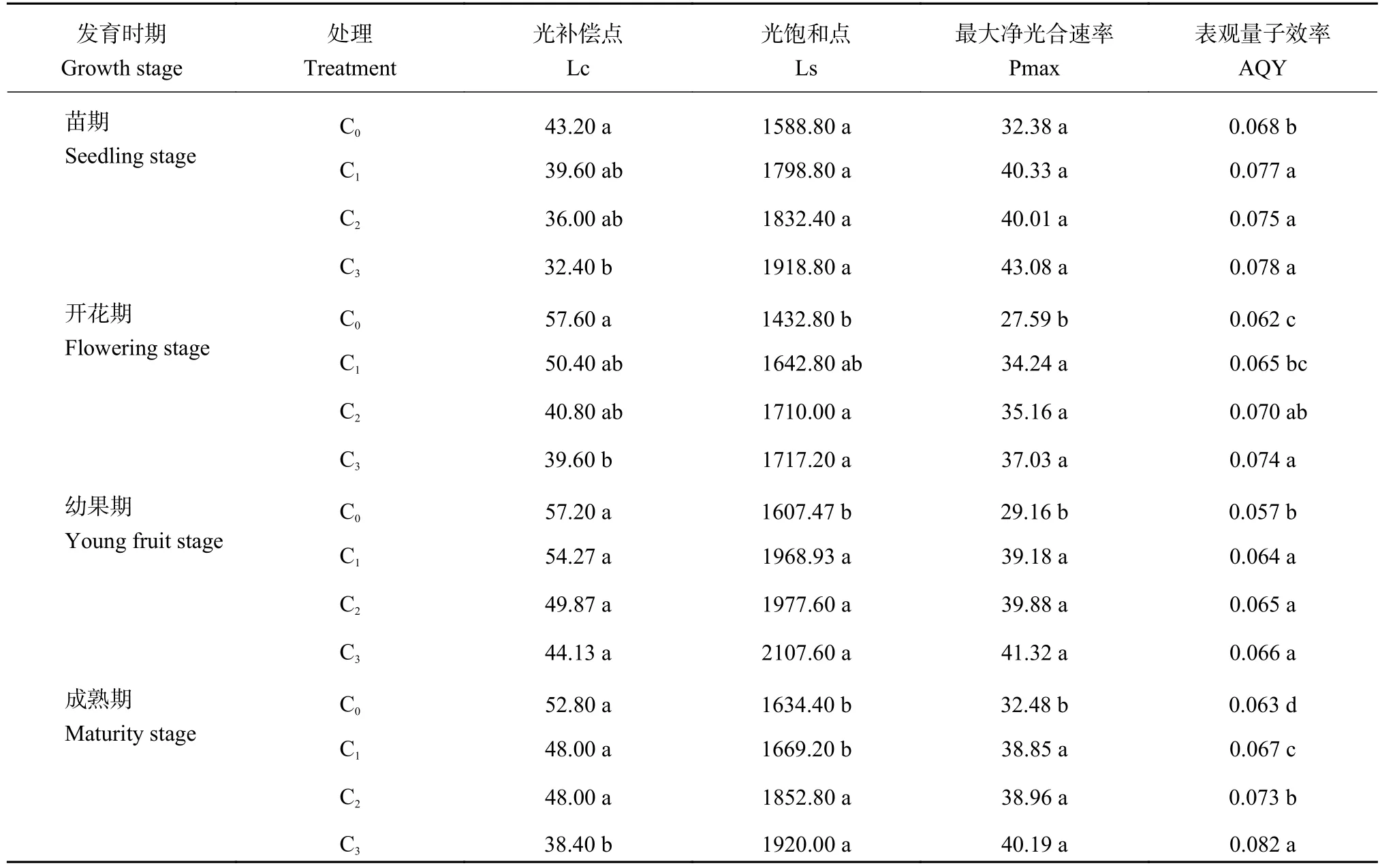
表2 不同CO2浓度下番茄不同生育期叶片的光响应参数 [μmol/(m2·s]Table2 Light-response parameters of tomato of different growth stage under different CO2 concentrations
2.4 不同浓度的CO2施肥对番茄产量的影响
表3显示,平均单果重、平均单株产量及产量均随着CO2浓度的升高而显著增加。与对照相比,增施CO2处理的番茄分别增产 10.8%、26.4%、33.9%。
3 讨论与结论
植物叶片中的光合色素是叶片进行光合作用的物质基础,其中叶绿素是叶片进行光合作用的敏化剂,植物叶片中包含的绝大部分叶绿素a和全部叶绿素b主要起捕捉和传递光能的作用,少量特殊状态的叶绿素a作用是将光能转化为电能,而且植株叶片中叶绿素含量与净光合速率呈极显著正相关[19];而类胡萝卜素能将吸收的光能传递给叶绿素a,也是光合作用中不可缺少的光合色素,即光合色素含量的高低可以反映植株叶片光合能力的强弱,而且生长于不同环境下的植株叶片中的光合色素素含量也会有所差异。本试验结果表明,不同CO2浓度处理间番茄叶片中光合色素含量虽然未达到显著差异,但随着CO2浓度的升高,光合色素均有不同程度的增加,且开花结果期增势较苗期和成熟期明显,即增施CO2能提高番茄各生育期叶片中的光合色素含量。该结果不仅与熊珺等[20]的研究结果一致,而且在其它作物上也得到了证实,如CO2浓度升高能提高辣椒[10,16]、豇豆[21]等叶片中的光合色素含量。而且,本研究表明1000 μmol/mol的CO2浓度效果最佳,800 μmol/mol次之。

表3 不同CO2浓度下番茄的产量Table3 Yield of tomato under different CO2 concentrations
CO2是植物进行光合作用的主要原料,CO2浓度升高对绿豆[22]、谷子[23]及水稻[24]等农作物生长发育的影响的研究表明,CO2浓度升高能通过提高作物的光合能力,进而提高其产量[25]。其浓度升高促进植物光合作用和增产的机制主要是通过提高RUBP羧化酶的活性,抑制RUBP加氧酶的活性,降低植物的光呼吸强度,提高植物的光合作用,从而促进植物光合产物的积累[26]。CO2浓度升高使植物叶片气孔导度和蒸腾速率降低,净光合速率、胞间CO2浓度和水分利用率增加[22,27–29]。但也有研究发现,CO2浓度升高对某些C4杂草的光合作用没有促进作用[30]。本试验结果表明,随着CO2浓度的升高,番茄叶片的净光合速率、胞间CO2浓度和水分利用率在番茄各生育期均显著增加,且以1000 μmol/mol浓度处理下的增幅最大;气孔导度和蒸腾速率均有不同程度的降低。有研究表明,短期CO2浓度升高能促进植物光合作用,但植物长期处于高浓度CO2环境中会出现光合适应现象[5,28,31]。本研究在增施 CO2的过程中,净光合速率始终处于上升阶段,说明该试验设置下的高CO2浓度对番茄的光合作用的促进效果没有随着施入CO2时间的延长而减弱,即试验期间未发生光合适应现象。其原因可能是由于本试验设置的CO2浓度低于番茄叶片CO2饱和点,或者是试验过程中氮肥充足,植株吸收的氮能够满足其光合作用需要,使得净光合速率长时间保持在较高的水平,抑制了光合适应现象[32]。也可能与本试验是在温度较低的冬春季节进行有关,试验期间昼短夜长,叶片在白天积累的淀粉和可溶性糖等光合产物在夜间被迅速转移或呼吸消耗,维持了番茄植株内的源库平衡,避免了光合的反馈抑制[33]的出现。有研究表明,增施CO2条件下,提供充足的光照可以进一步促进黄瓜叶片的光合作用[34]。因此,未发生光合适应现象也可能与该试验在测定光合作用有关参数时设置的光合有效辐射 (PAR) 有关。
通过光响应曲线可得出番茄叶片的光补偿点、光饱和点、最大净光合速率及表观量子效率。与对照处理相比,增施CO2处理能够降低光补偿点,尽管差异不显著,但在苗期、开花期、幼果期及成熟期分别平均降低了16.7%、24.3%、13.6%、15.2%,说明增施CO2能够提高番茄叶片对弱光的利用能力,其中开花期对弱光的利用能力较其它生育期强。而光饱和点、最大净光合速率和表观量子效率均随着增施的CO2浓度的升高而增大,其中,C3处理即CO2浓度为1000 μmol/mol时增幅达到最大。光饱和点反映植物对强光的忍受性[35],说明增施高浓度CO2能够增强番茄在生长发育过程中对强光的忍受性;叶片最大净光合速率反映了植物体具有的最大光合潜能[36],试验结果表明,与对照相比,增施CO2后,最大净光合速率在苗期、开花期、幼果期及成熟期分别平均增加了27.0%、29.0%、37.6%、21.1%,说明增施CO2能够有效提高番茄叶片的光合能力,在开花结果期番茄的光合潜能较苗期和成熟期强,而且该试验条件下以800、1000 μmol/mol的CO2浓度效果较好。
综上所述,本试验条件下,增施不同浓度CO2后,以浓度为 800、1000 μmol/mol CO2明显提高了番茄各生育期叶片光合色素含量和光合生理特性,有利于番茄产量的增加。即本研究可为增施CO2浓度条件下北方温室番茄及其它果菜生产提供一定的参考依据。但该试验设置的CO2浓度最高为1000 μmol/mol,加之番茄叶片的CO2饱和点较高,若是CO2浓度继续升高对番茄光合特性等生长发育的影响可能会与本试验结果有所不同,故对于CO2浓度高于1000 μmol/mol后对番茄生长发育的影响有待今后开展进一步的研究。
