转CarNAC1基因可提高棉花的抗旱性
2018-08-07王淑君曲延英倪志勇王莉萍高文伟陈全家
王淑君,曲延英,倪志勇,王莉萍,高文伟,陈全家
(新疆农业大学农学院,农业生物技术重点实验室,新疆 乌鲁木齐 830052)
非生物胁迫抑制棉花正常生长发育,制约产量,降低纤维品质,持续胁迫可能导致棉株死亡[1]。干旱是当前面临的主要非生物胁迫[2]。全球干旱地区耕地面积占总耕地面积的20%左右,约2.74亿公顷[3]。棉花是世界性重要的经济作物,它虽然是较抗旱的作物[4],但抗旱能力存在品种差异[5]。结合当前严峻形势,培育高抗旱棉花品种,可提高农民经济效益,促进农业可持续发展[6]。与传统育种相比,转基因技术可以通过导入目的基因,定向的改良棉花农艺性状,提高产量,加速棉花育种进程[7]。然而,关于转基因提高棉花抗旱性的报道较少[8]。
转录因子通过与目标基因启动子中特定的DNA序列即顺式作用元件结合,激活或抑制靶基因的转录表达,是基因表达中一类重要的调控因子[9]。研究表明,DREB、MYB、Bzip、锌指结构等转录因子在提高植物胁迫耐受性方面发挥着重要作用[10]。NAC转录因子家族被认为是植物所特有的一类转录因子,命名源于矮牵牛NAM和拟南芥ATAF1、ATAF2以及CUC2基因,广泛存在于各种植物中,而在其他的真核生物中还未发现[11]。
NAC转录因子参与高等植物顶端分生组织的形成、细胞周期的调控、种子和胚的发育、花药的发育、次生根的形成、次生壁的形成、叶片的衰老、激素信号转导等生物学过程,同时还具有调控逆境胁迫应答的功能[12]。关于NAC基因家族的研究主要集中在水稻[13]、小麦[14]、玉米[15]、大豆[16]、拟南芥[17]、番茄[18]、紫花苜蓿[19]等植物上。
鹰嘴豆是世界第三大豆科作物,主要生长于干旱或者半干旱地区,其生育期短、抗逆性强、基因组小,在解析植物的抗旱分子机制和挖掘抗旱基因方面具有重要意义[20]。前人从鹰嘴豆中克隆得到CarNAC1基因,并在拟南芥中证明该基因能提高植株抗旱耐盐能力[21]。本研究对15个稳定遗传的转CarNAC1基因棉花品系以及受体材料k62,通过室内、室外试验,比较转基因棉花品系与受体材料k62在干旱条件下光合指标、生理指标、纤维品质以及产量间的差异,筛选抗旱能力较强的棉花品系。
1 材料与方法
1.1 材料
受体材料K62及经检测能稳定遗传的15个转基因棉花品系:ZK62-1、ZK62-2、ZK62-3、ZK62-4、ZK62-5、ZK62-6、ZK62-7、ZK62-8、ZK62-9、ZK62-10、ZK62-11、ZK62-12、ZK62-13、ZK62-14、ZK62-15。新疆农业大学农业生物技术重点实验室提供以上供试材料。
1.2 试验方案
1.2.1田间试验 试验于2016年在石河子144团新疆农业大学试验田进行。该地区无霜期较长,气候条件适宜棉花种植。5月1日播种,每份材料一膜,株距为10 cm,种植5 m。同一膜相邻材料以50 cm的走道隔离开来。每个供试材料设两个处理,处理1正常灌水:全生育期正常浇水;处理2干旱胁迫:于7月5日至7月21日(花铃期)不浇水,后正常灌水;每个处理均重复3次,该试验用地面积1.32×103m2。复水前一日测定不同处理下供试材料各项光合指标(均以倒2叶为准)。9月16日(吐絮期)测定农艺性状,在不同处理下收获20铃测定单铃重和衣分含量,每份材料保存一定量的皮棉样品送往乌鲁木齐棉花质量监督检验测试中心测定纤维品质。
1.2.2室内试验 试验于2016年在新疆农业大学农业生物技术重点实验室进行。通过大田试验结果选择抗旱性具有差异的两个棉花品系:ZK-6、ZK-15以及受体材料K62进行室内试验。选用饱满且胚完整的种子,经消毒处理后播种于花盆中,待长出一片子叶后移入装有1/2Hoagland营养液的塑料盆。水培试验在光照培养室进行,光周期为12 h/12 h,昼夜温度为:26~28℃/20~22℃,每2~3 d按时更换营养液保证幼苗健康生长。水培至4~5片真叶时,用15%PEG-6000进行处理,分别在处理0、3、6、12、24 h后采取叶片并测定脯氨酸、丙二醛、可溶性糖、过氧化氢酶、过氧化物酶、超氧化物歧化酶的含量,每处理重复3次。
1.3 测定指标与方法
1.3.1 光合生理生化指标 采用英国(汉莎)CIRAS-3便携式光合测定系统,于7月13日以及7月21日10∶30-12∶30(此时间段为当地测量最佳时间,避免“光合午休”现象)进行光合测量。均以测定倒2叶为准,测定净光合速率(Pn)、气孔导度(Gs)。
1.3.2 农艺性状指标 每个品系不同处理下连续测定5株棉花的株高、第一果枝节位、始节高、果枝数、有效果枝数、总铃数、有效铃数;收取20铃,计算单铃重、衣分含量、皮棉产量、籽棉产量。
1.3.3 纤维品质指标 考种后保存的皮棉样品送至乌鲁木齐农业部棉花质量监督检验测试中心,测定纤维上半部平均长度、整齐度、马克隆值、伸长率、短纤维率、纺稳参数和断裂比强度等指标。
1.3.4 生理指标的测定 脯氨酸用茚三酮比色法测定;可溶性糖用蒽酮比色法测定;丙二醛用硫代巴比妥比色法测定;总超氧化物歧化酶用羟胺法测定;过氧化物酶用可见光法测定;过氧化氢酶用分光光度法测定;以上各指标均使用南京建成生物工程研究所订购的试剂盒测定。
1.4 数据处理及统计方法
所有数据的统计和分析均在SPSS 19.0和Excel 2013上完成,Excel 2013进行绘图。
数据处理时进行标准化处理,消除遗传背景对抗旱性研究的影响,参考王艳慧等[22]方法,采用(干旱值/正常值)相对值进行综合评定。
(1)
(2)
式中,Xi、CKi分别表示干旱、正常处理的性状测定值。
采用隶属函数法进行抗旱性综合评价。隶属函数值的计算如公式(3)所示。
X(u)=(x-xmin)/(xmax-xmin)
(3)
X(u)反=1-(x-xmin)/(xmax-xmin)
(4)
式中,x为各品种(系)的某一指标测定值,xmax和xmin分别为所有品种(系)某一指标的最大值和最小值。如果某一指标与抗旱(耐盐)性呈负相关,则用反隶属函数进行转换,如公式(4)所示。
综合指标权重:
(5)
式中,wi为第i个综合指标在所有综合指标中的重要程度即权重;pi为各基因型第i个综合指标的贡献率。
综合抗旱能力:
(6)
式中,D值为各棉花品系在干旱胁迫条件下用综合指标评价所得的抗旱性综合评价值。
2 结果与分析
2.1 干旱胁迫对棉花光合指标的影响
由图1和图2可看出,干旱胁迫后,转基因棉花品系气孔导度显著降低,均低于受体材料k62;转基因棉花品系净光合速率均高于受体材料k62。由此结果说明转基因棉花品系可能通过降低气孔导度,来提高棉花品系的抗旱能力。
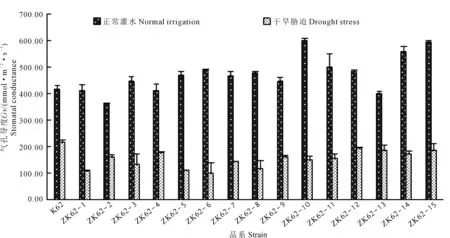
图1 干旱胁迫下棉花叶片气孔导度的变化Fig.1 Changes of stomatal conductance of cotton leaves under drought stress
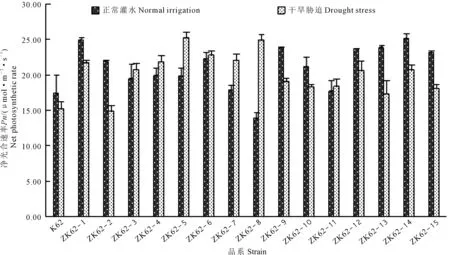
图2 干旱胁迫下棉花叶片净光合速率的变化Fig.2 Changes of net photosynthetic rate of cotton leaves under drought stress
由表1可知,干旱胁迫下,供试材料气孔导度在98.8~284.2 mmol·m-2·s-1之间,转基因棉花品系气孔导度显著低于受体材料K62,其中转基因棉花品系ZK62-6的气孔导度与受体材料K62差异性最显著,低于受体材料185.4 mmol·m-2·s-1;光合速率在14.93~25.29 mmol·m-2·s-1之间,除ZK62-2外,转基因棉花品系的光合速率均显著高于受体材料K62,其中转基因品系ZK62-6的净光合速率与受体材料K62差异性最显著,高于受体材料10.067 μmol·m-2·s-1;

表1 干旱胁迫下转基因棉花与受体材料光合指标的比较Table 1 Comparison of photosynthetic indexes of transgenic cotton and receptor materials drought stress
注:*代表0.05显著水平;**代表0.01显著水平。下同。
Note:*represent 0.05 significant level;**represent 0.01 significant level. The same below.
2.2 干旱胁迫对转基因棉花农艺性状和产量性状的影响
2.2.1 干旱胁迫下转基因棉花品系与受体材料农艺性状和产量性状比较 由表2可得,干旱胁迫下,各转基因棉花品系与受体材料K62性状差异达显著水平所占比例分别为:株高占60%、第一果枝节位占60%、果枝数占53.3%、有效果枝数占20%、总铃数占20%、有效铃数占40%、单铃重占100%。由此可知,干旱胁迫下转基因棉花品系单铃重、株高、始节高、果枝数差异幅度较大。转基因棉花品系单铃重均高于受体材料K62,ZK62-15与K62差异最大,平均高于对照0.657g;除ZK62-10外,其它转基因棉花品系株高均低于受体材料K62,ZK62-5与K62差异最大,平均低于对照8.8 cm;转基因棉花品系第一果枝节位均低于受体材料K62,ZK62-6与K62差异最大,平均低于对照7.5 cm;转基因棉花品系果枝数均多于受体材料K62,ZK62-14与K62差异最大,平均多于对照3.5个。
2.2.2 基于农艺性状和产量性状主成分分析 对9个单项指标的抗旱系数进行主成分分析(表3),前2个综合评价指标F1和F2的累积贡献率为76.619%,因此,F1和F2基本能反映9个农艺性状及产量性状指标的变化。
2.2.3 基于农艺性状和产量性状指标抗旱性综合评价 由表4可知,CDC值与D值结果基本一致。在干旱胁迫下,大部分转基因棉花品系综合抗旱能力均优于受体材料k62。综合抗旱能力排在前三的品系分别为ZK62-10、ZK62-6、ZK62-1。综合抗旱能力相对较差的品系为:ZK62-13、ZK62-9、ZK62-4。

表2 干旱胁迫下转基因棉花品系与受体材料农艺性状和产量性状比较
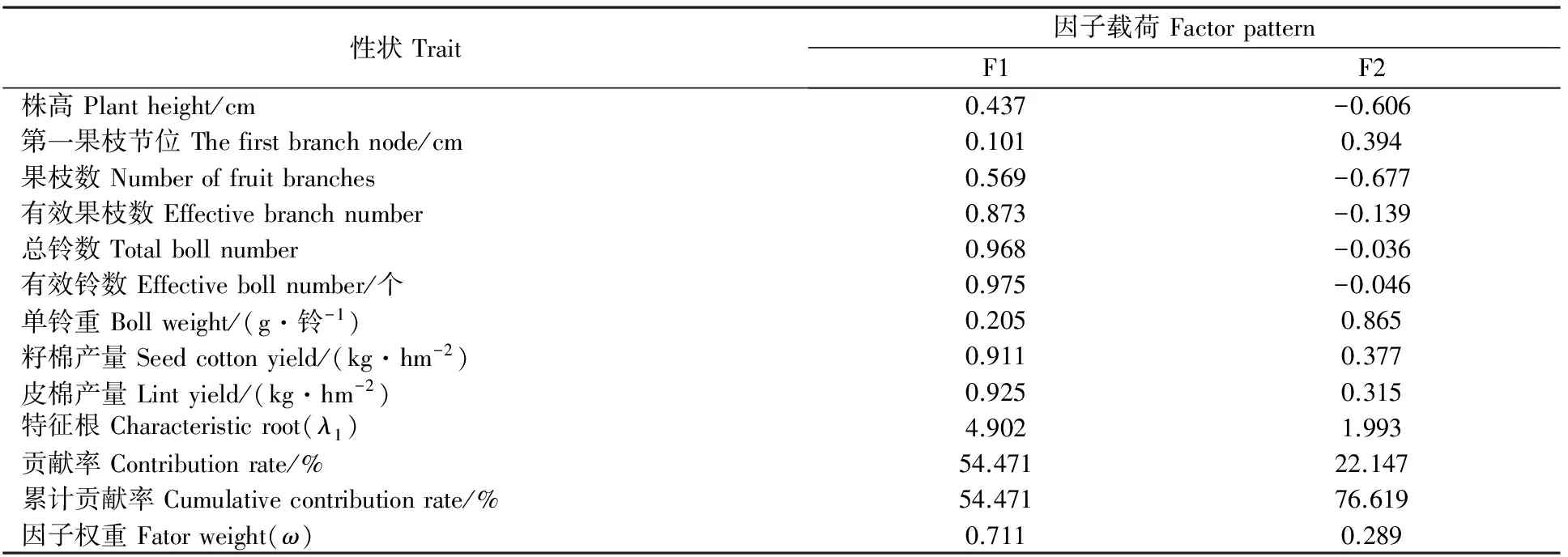
表3 干旱胁迫下转基因棉花品系农艺性状和产量性状指标因子载荷矩阵及方差贡献率Table 3 The factor loading matrix and the variance contribution rate of agronomic characterrs and yield traits of transgenic cotton strains under drought stress
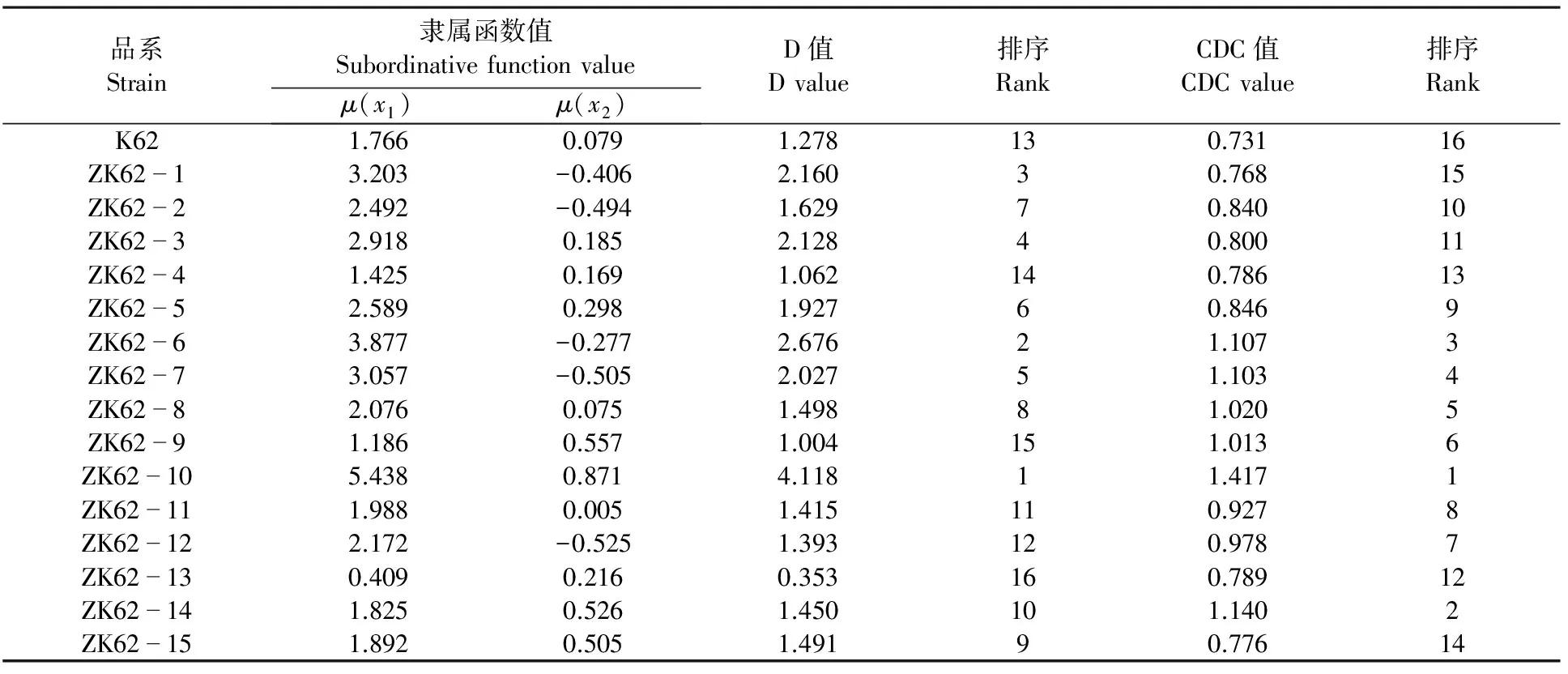
表4 干旱胁迫下转基因棉花品系农艺性状和产量性状的D值、CDC值及排序Table 4 D value, CDC value and the rank of agronomic characters and yield traits of transgenic cotton strains under drought stress
2.2.4 基于农艺性状和产量性状指标聚类分析 通过最近邻元素法进行聚类分析,结果如图3所示,干旱胁迫下,将16个棉花品系分为三类,第一类为极抗旱材料1份:ZK62-10;第二类为中抗旱材料5份:ZK62-1、ZK62-7、ZK62-5、ZK62-3、ZK62-6;第三类为敏感材料10份:ZK62-8、ZK62-11、k62等。
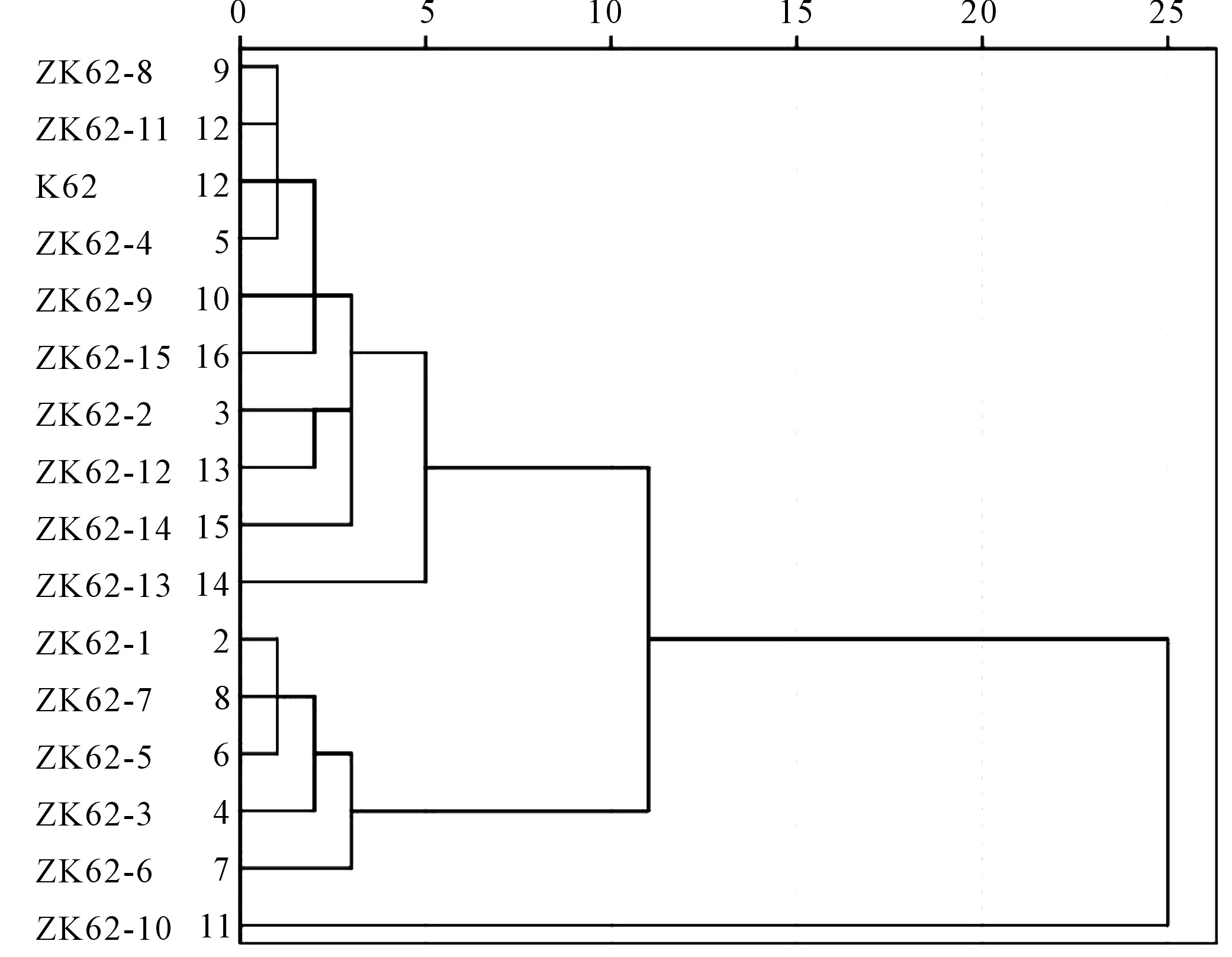
图3 基于农艺性状和产量性状指标D值的聚类Fig.3 Clusters based on D value of agronomic traits and yield
2.3 干旱胁迫对棉花纤维品质的影响
2.3.1 干旱胁迫下转基因棉花品系与受体材料纤维品质比较 由表5可知,干旱胁迫下,各转基因棉花品系与受体材料K62性状差异达显著水平所占比例分别为:纺纱系数占80%、马克隆值占100%、上半部长度占100%、长度整齐度占73.3%、短纤维率占86.7%、断裂比强度占80%、断裂伸长率占93.3%。由此可知,干旱胁迫下转基因棉花品系与受体材料K62在马克隆值、上半部长度、断裂伸长率的差异幅度较大。马克隆值在3.12~5.69之间,其中K62-4与受体材料K62差异性最大,差值为1.79;上半部长度在27.2~30.46 mm之间,其中K62-4与受体材料K62差异性最大,差值为2.0 mm;断裂伸长率在3.17~6.68 CN·tex-1之间,其中K62-5与受体材料K62差异性最大,差值为2.62 CN·tex-1。
2.3.2 基于纤维品质指标主成分分析 由表6可知,对7个单项纤维品质指标的抗旱系数进行主成分分析,前3个综合评价指标F1、F2、F3的累积贡献率为91.395%。因此,前3个综合评价指标基本能反映纤维品质状况。
2.3.3 基于纤维品质指标抗旱性综合评价 由表7可知,CDC值与D值结果基本一致。在干旱胁迫下,大多数转基因棉花品系综合抗旱能力均优于受体材料K62。干旱胁迫下纤维品质依然能保持较高水平的品系分别为ZK62-6、ZK62-12、ZK62-15。纤维品质受干旱胁迫影响较大的品系为:ZK62-10、ZK62-1、K62。
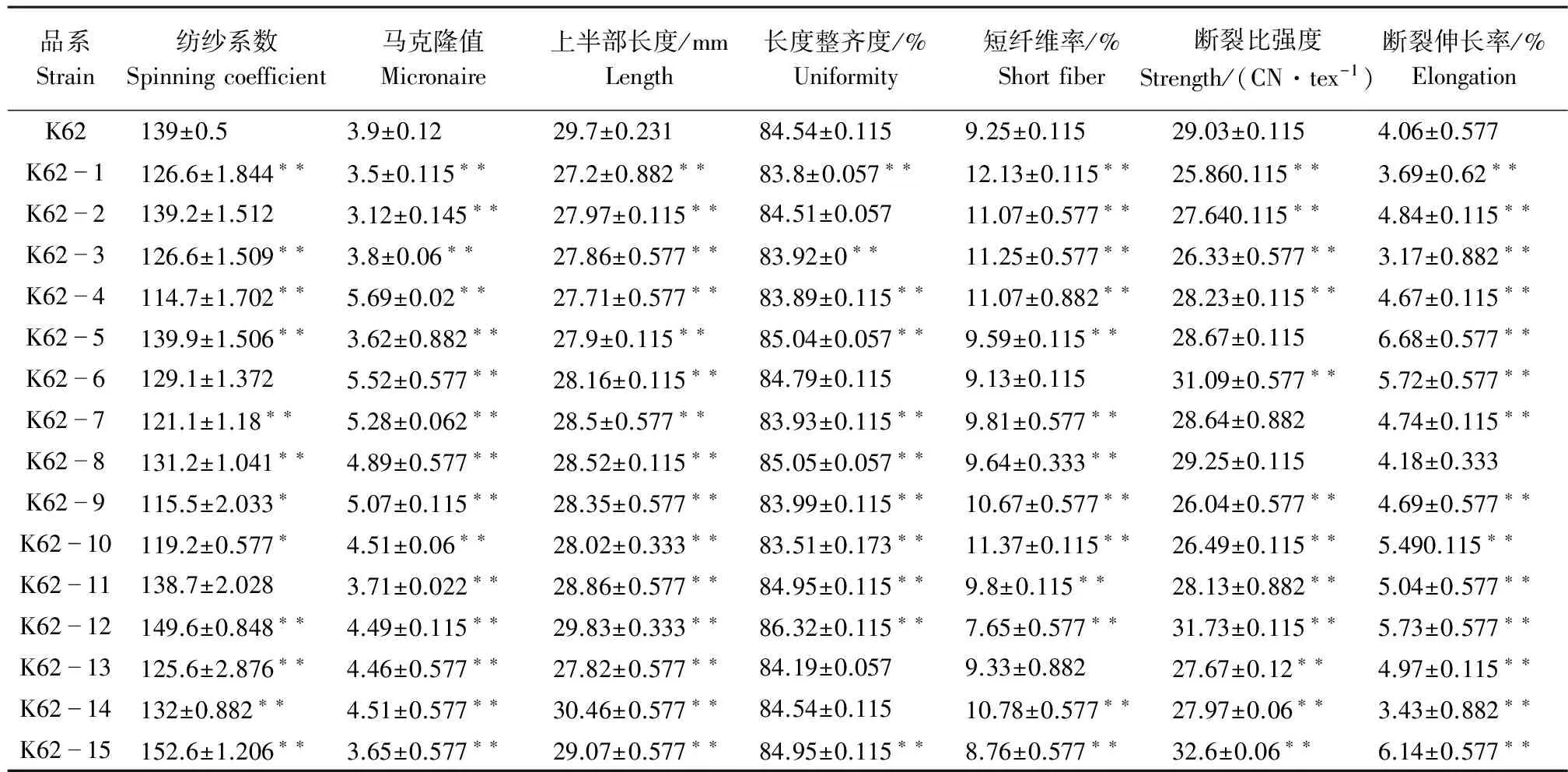
表5 干旱胁迫下转基因棉花品系与受体材料纤维品质比较

表6 干旱胁迫下转基因棉花品系纤维品质指标因子载荷矩阵及方差贡献率Table 6 The factor loading matrix and the variance contribution rate of fiber quality index of transgenic cotton strais under drought stress
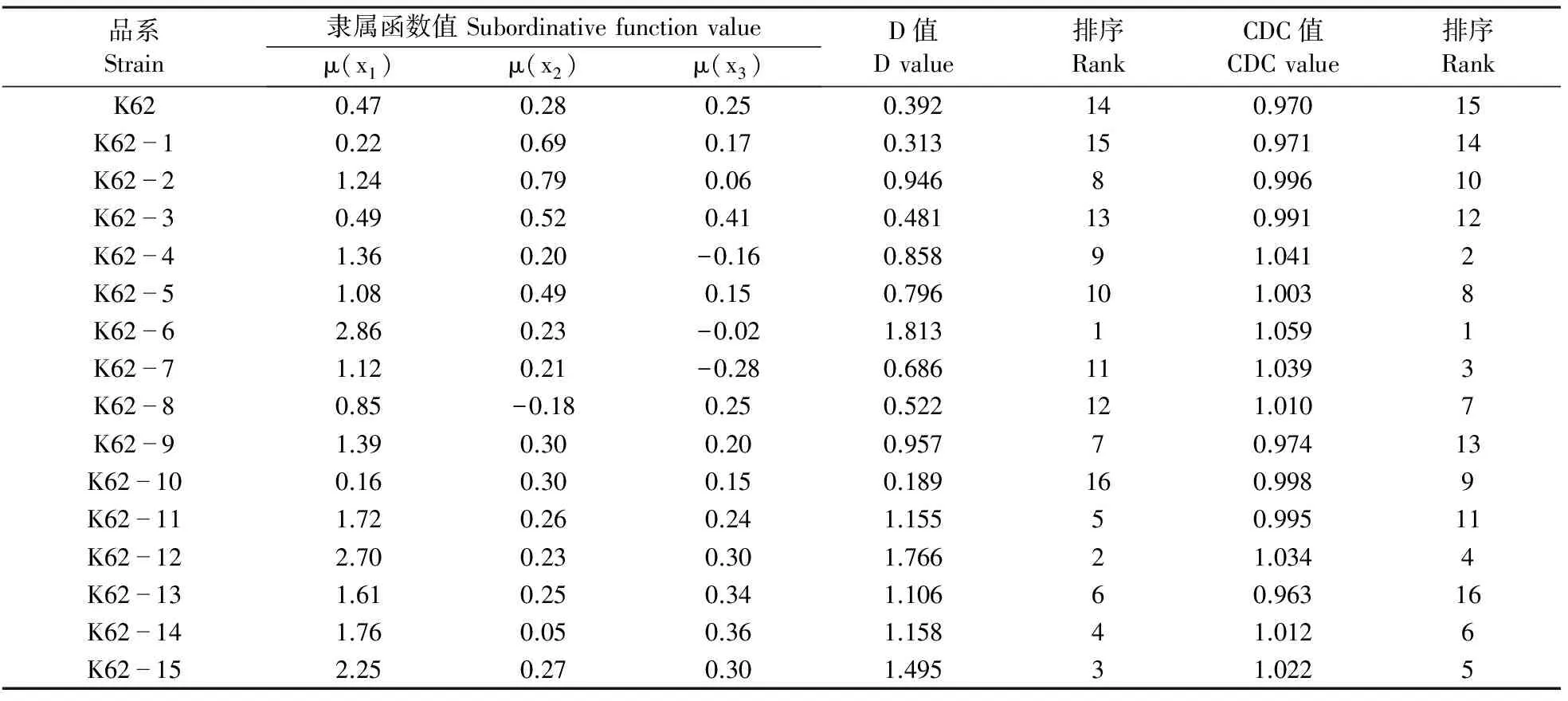
表7 干旱胁迫下转基因棉花品系纤维品质指标的D值、CDC值及排序Table 7 D value, CDC value and the rank of fiber index of transgenic cotton strains under drought stress
2.3.4 基于纤维品质指标聚类分析 通过最近邻元素法进行聚类分析,结果如图4所示,干旱胁迫下,将16个棉花品系分为三类,第一类为极抗旱材料3份:ZK62-6、ZK62-12、ZK62-15;第二类为中抗旱材料8份:ZK62-9、ZK62-13、ZK62-11等;第三类为敏感材料5份:K62、ZK62-10、ZK62-3、ZK62-1、ZK62-8。
2.4 室内模拟苗期干旱胁迫对棉花生理指标的影响
由图5可知,棉花在受到干旱胁迫后,丙二醛含量、脯氨酸含量、可溶性糖含量、总超氧化物歧化酶活力随着胁迫时间的增长均呈上升趋势,过氧化氢酶活力、过氧化物酶酶活力呈现先升高后降低的趋势。除丙二醛含量为转基因棉花品系低于受体材料K62外,其余各指标均为转基因棉花品系高于受体材料K62。
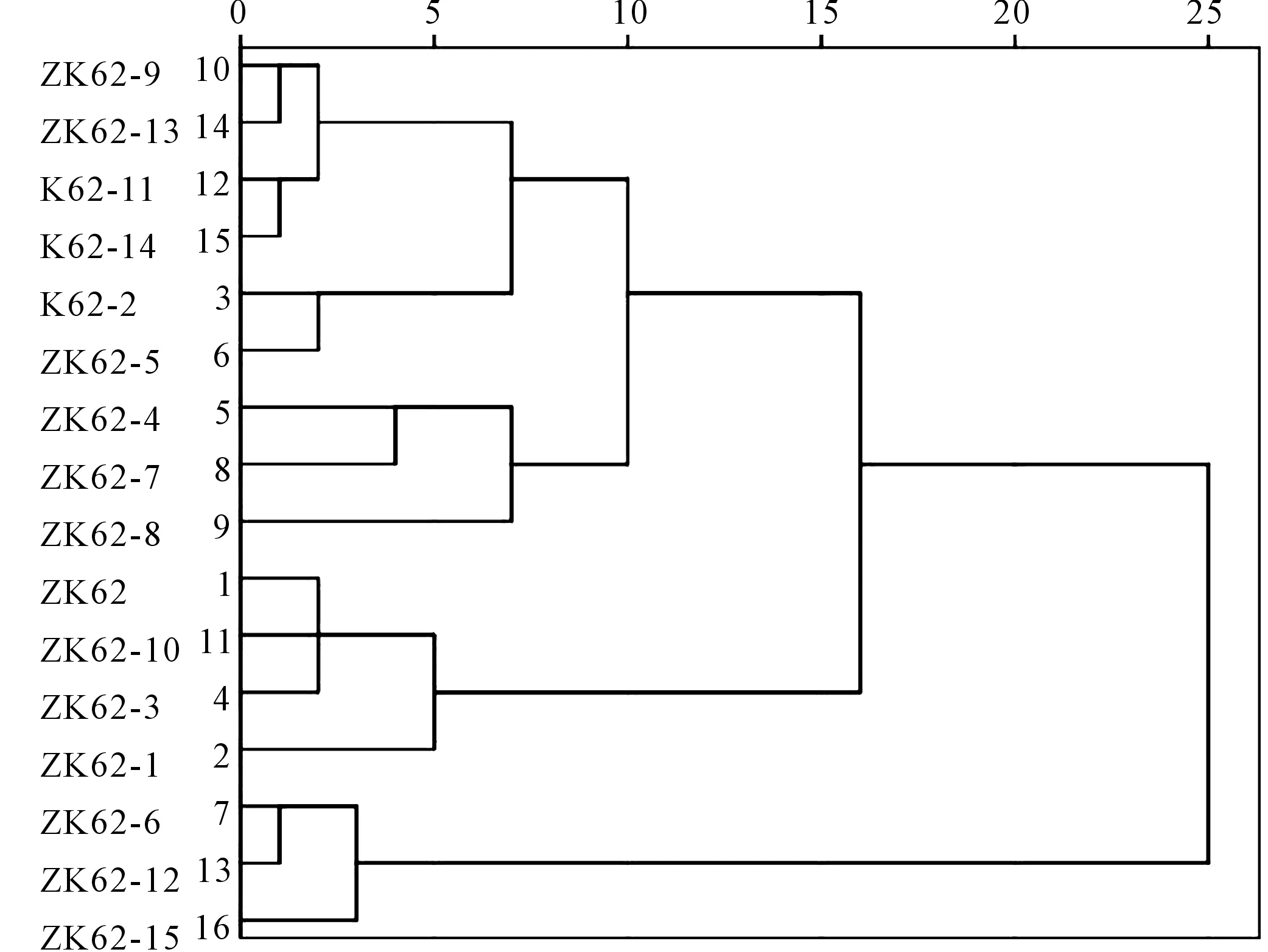
图4 基于纤维品质指标D值的聚类Fig.4 Clusters based on D value of fiber quality index
由表8可知,棉花苗期在室内进行模拟干旱,转基因棉花品系与受体材料K62在胁迫24 h时,丙二醛含量差异最大,ZK62-6丙二醛含量比K62低15.534 nmol·ml-1,ZK62-15丙二醛含量比K62低2.869 nmol·ml-1;转基因棉花品系ZK62-15脯氨酸含量在12 h时,与受体材料K62差异最大,高于受体材料5.71 μg·g-1,转基因棉花品系ZK62-6脯氨酸含量在24 h时,与受体材料K62差异最大,高于受体材料12.42 μg·g-1;转基因棉花品系ZK62-15可溶性糖含量在6 h时,与受体材料K62差异最大,高于受体材料2.92 mg·g-1,转基因棉花品系ZK62-6可溶性糖含量在24 h时,与受体材料K62差异最大,高于受体材料6.69 mg·g-1;转基因棉花品系ZK62-15过氧化氢酶活性在12 h时,与受体材料K62差异最大,高于受体材料6.91 U·mg-1,转基因棉花品系ZK62-6过氧化氢酶活性在干旱胁迫3 h时,与受体材料K62差异最大,高于受体材料18.27 U·mg-1;转基因棉花品系ZK62-15超氧化物歧化酶活性在干旱胁迫24 h时,与受体材料K62差异最显著,比受体材料高33.17 U·mg-1,转基因棉花品系ZK62-6超氧化物歧化酶活性在干旱胁迫12 h时,与受体材料K62差异最显著,比受体材料高65.65 U·mg-1;转基因棉花品系过氧化物酶活性在干旱胁迫24 h时,与受体材料K62差异最显著,ZK62-15比受体材料高85.88 U·mg-1,ZK62-6比受体材料高208.95 U·mg-1。
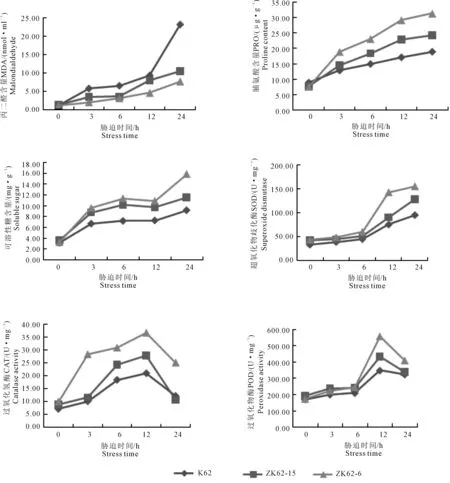
图5 干旱胁迫下棉花生理指标随胁迫时间的变化Fig.5 Changes of physiological indexes of cotton with stress time under drought stress
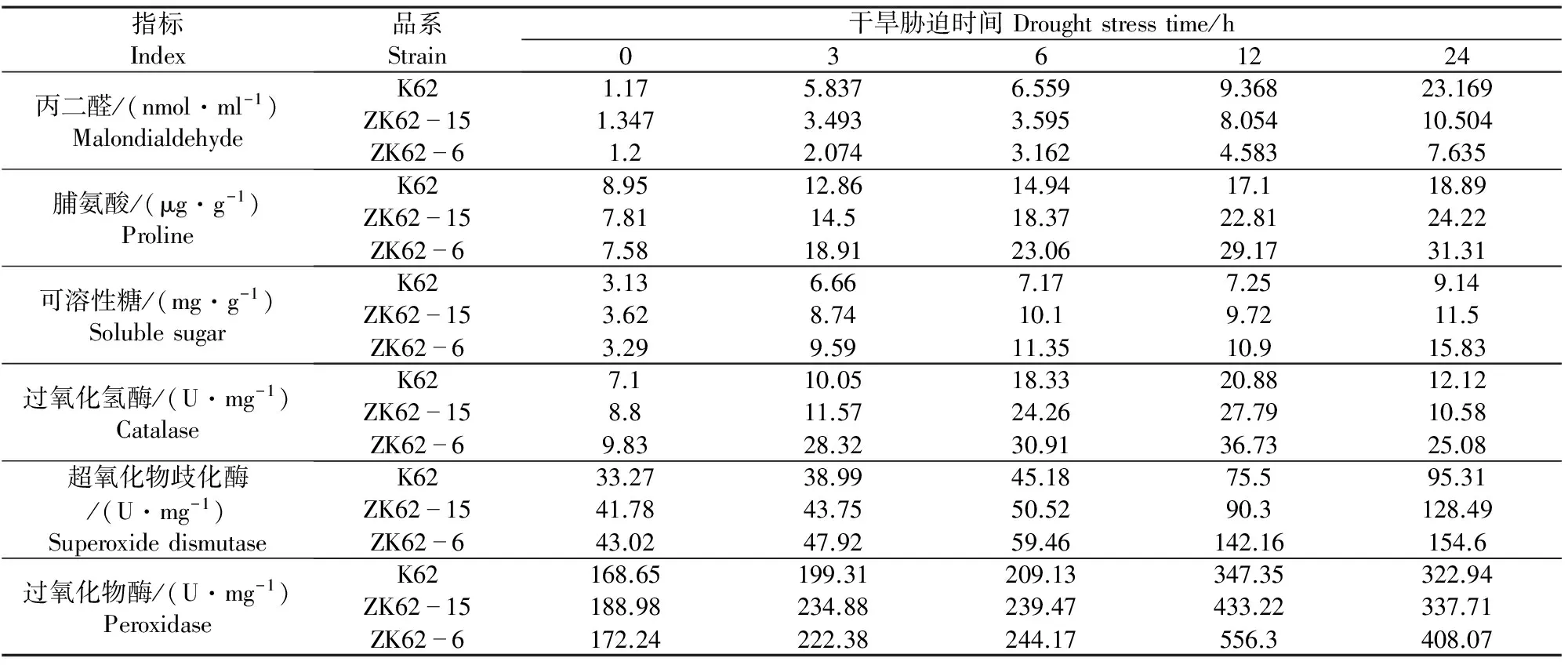
表8 干旱胁迫下转基因棉花品系与受体材料生理指标的比较Table 8 Comparison of physiological indexes of transgenic cotton and receptor materials under drought stress
3 讨 论
干旱是影响植物生长发育的重要逆境因子。研究发现[23],NAC转录因子直接参与或通过调节参与干旱、高盐、冷害应答基因的表达,在植物非生物逆境胁迫中起重要作用。本研究通过多角度实验证明,在棉花生长关键期——花铃期遭遇干旱时,转CarNAC1基因棉花抗旱能力在产量和纤维品质上均高于受体植株。有研究表明[24],OsNAC10基因过量表达,显著提高水稻抗旱耐盐性,且正常情况下,转OsNAC10基因使水稻产量增加5%~14%,干旱条件下相对于对照增产25%~42%,该结果与本试验结果一致。
棉花纤维的长度、强度、马克隆值等品质是评价棉花品质的重要指标。培育纤维品质较好的棉花品种是当前棉花育种工作重点之一[25]。目前,NAC基因的研究集中在拟南芥[26]、水稻[27]等植物上,在棉花中的研究较少。Zhong[28]研究报道了NAC转录因子SND1参与调控拟南芥纤维次生壁合成,在木质部纤维和茎维管束纤维特异表达,SND1过表达或抑制造成纤维次生壁的加厚或缺失。本研究表明,在干旱胁迫后,转CarNAC1基因棉花在提高其抗旱性的同时,与受体材料相比,增加棉花产量并保持相对较高的纤维品质水平。由此推测,转CarNAC1基因棉花的纤维品质受干旱胁迫影响较小。有研究表明[29]NAC基因参与调节棉纤维细胞次生壁的合成。CarNAC1基因是否参与调控棉花纤维细胞的合成有待于进一步研究。
光合作用是植物生长和产量形成的生理基础[30],气孔作为水分和CO2进出叶片的通道,通常在干旱、高温等逆境胁迫下,叶片气孔都有不同程度的关闭[31]。研究表明[32],水稻SNAC1基因在干旱胁迫下,气孔保卫细胞特异表达,促进气孔关闭,降低水分蒸腾速率,但不影响光合速率,因而植株抗旱性大为提高。本试验结果显示,转CarNAC1基因棉花在受到干旱胁迫后,气孔导度迅速下降,可能是由于CarNAC1在干旱胁迫下,气孔保卫细胞特异性表达,保持了相对较高的光合能力。通过隶属函数法对16个棉花品系进行综合评价,发现大多数转CarNAC1棉花品系在干旱胁迫下的综合抗旱能力均高于非转基因棉花,并一定程度上提高了植株的生物量和籽棉产量,但少数转CarNAC1棉花品系在干旱胁迫下的综合抗旱能力低于非转基因棉花。该现象可能是由于响应干旱胁迫时,NAC转录因子除了被诱导表达外,也可能存在表达受抑制的情况。如Han[33]研究结果表明,番茄SlNAC3基因在盐、干旱和ABA胁迫处理下其转录表达受抑制。
渗透调节是植物耐受干旱环境的重要生理机制[34]。丙二醛(MDA)、脯氨酸(PRO)、可溶性糖、过氧化氢酶(CAT)、过氧化物酶(POD)、总超氧化物歧化酶(SOD)都是与旱胁迫密切相关的理化指标。本实验研究表明,室内培养条件下,在棉花苗期用15%的PEG干旱模拟胁迫后,供试材料的丙二醛含量随胁迫时间增加均呈现上升趋势,但转基因棉花的丙二醛含量显著低于受体材料K62。转基因棉花品系的脯氨酸、可溶性糖、过氧化氢酶、过氧化物酶以及总超氧化物歧化酶活力均显著高于受体材料K62。由以上结果可以推测外源CarNAC1基因可能提高了棉花抗旱性,增强了棉花自我渗透调节能力。
综上所述,本文通过室内试验和田间试验多方面证明,转CarNAC1基因提高了棉花抗旱能力,并筛选出综合抗旱能力较强的品系ZK62-6,将有助于推动干旱区棉花转基因育种工作。