小麦氮、磷胁迫响应基因研究进展
2018-08-02潘剑君
王 峻,宋 科,薛 永*,潘剑君
(1.上海市农业科学院 生态环境保护研究所/农业部上海农业环境与耕地保育科学观测实验站/上海农业环境保护监测站/上海低碳农业工程技术研究中心,上海 201403;2.南京农业大学 资源与环境科学学院,江苏 南京 210095)
小麦是世界上种植面积最大、总产量最高的粮食作物[1]。小麦是植物蛋白质的主要来源,其产量和品质一直备受人们的关注。氮、磷是蛋白质构成的重要元素,对小麦生长发育和产量形成具有十分重要的作用[2-3]。
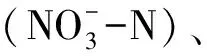
基于以上严峻的现实问题,寻求有效的解决方法变得尤为重要。提高肥料利用率,减少化肥投入,做到减量不减产、增效又增质是人们的愿望和目标。种植养分高效的高产作物品种是行之有效的途径,而探明作物高效吸收利用养分的分子机制是培育高效高产新品种的重要基础。控制作物养分吸收利用的基因复杂多样。有研究表明,一种基因可以调控植物对多种营养元素的应答[8-9];一种营养元素的变动也会引起植物多种基因的响应[10]。在养分胁迫条件下,不同作物或同种作物不同基因型抗逆能力也存在较大差异[11]。这在一定程度上增加了探究养分吸收利用分子机制的难度。本研究主要从miRNA(microRNA)、LBD (Lateral organ boundaries domain)、MAPK(Mitogen-activated protein kinase)等基因方面综述了小麦氮胁迫响应基因,从bHLH(Basic-helix-loop-helix)、CDPK(Calcium-dependent protein kinase)、MAPK等基因方面综述了磷胁迫响应基因,旨在探明小麦氮、磷养分吸收利用的分子机制,为挖掘小麦氮、磷养分高效基因奠定基础。
1 氮胁迫下小麦响应基因研究进展
氮素是植物必需的营养元素,对植株的生长发育发挥重要调控作用,也是影响农作物产量和品质的主要因素之一[12]。在氮素供应不足时,植物体感知并传递胁迫信号,启动相关基因做出响应和应答,以抵御这种逆境环境。目前,发现的这类基因主要有miRNA、LBD、MAPK等。
1.1 miRNA基因
miRNA是生物体内普遍存在的一种非编码内源小分子RNA,能通过序列反向互补方式与其作用靶基因结合,在转录后水平或翻译水平对其靶基因进行调控。大多数植物种属miRNA作用的靶基因为编码调控蛋白基因,在植株生长发育调控和介导植株应答、抵御各种生物、非生物逆境中发挥着重要的作用[13]。目前,已发现的参与小麦氮胁迫的miRNA基因主要有TaMIR1129[13-14]、TaMIR1118[15]、TaMIR1136[15]、TaMIR444a[16-17]。
小麦TaMIR1129基因受低氮胁迫诱导表达,其表达量随氮浓度降低和处理时间延长不断升高[14]。同样,TaMIR1118和TaMIR1136基因也受低氮胁迫诱导表达[15]。在低氮条件下,转小麦TaMIR1129基因烟草植株形态增大,单株干质量显著增加,TaMIR1129基因表达上调,烟草植株抵御低氮胁迫的能力显著增强[13]。
在低氮胁迫下,小麦根和叶中TaMIR444a基因表达上调,其表达量随处理时间增加而升高,在处理48 h后达到最大值;其通过调控靶基因的转录和翻译,进一步改变TaMIR444a/靶基因模块下游基因的转录;通过分析,在超表达TaMIR444a基因烟草中鉴定了1 733个差异表达基因,其中有921个基因表达上调、812个基因表达下调;另外,低氮胁迫下,转TaMIR444a基因烟草中硝酸转运蛋白(NRT)基因NtNRT1.1-s、NtNRT1.1-t、NtNRT2.1和抗氧化酶基因NtCAT1;1、NtPOD1;3、NtPOD4表达量明显上调[16]。说明MIR444a可以在转录水平对大量基因进行调控。
1.2 LBD基因
LBD基因家族是一类植物特有的转录因子家族,参与调控植物侧生器官和分生组织的建成,对植物体内激素积累和氮素代谢等具有重要的调控作用[18-22]。目前,已发现的参与小麦氮胁迫的LBD基因主要为TaLBD[23-24]。
研究发现,在低氮、低磷胁迫条件下,小麦TaLBD基因表达量上升,在胁迫处理后9 h达到峰值,之后表达量下降[23]。Pant等[24]研究也发现,低氮胁迫下,TaLBD基因的表达量呈先上调后下调的趋势,并且与野生型植株相比,超表达TaLBD的烟草植株长势明显增强,单株干、鲜质量,叶面积和根量显著增加。表明TaLBD通过调控植株根系发育改善植株的养分状况,在介导植株抵御低氮逆境中发挥重要作用。
1.3 MAPK基因
MAPK是一类存在于真核生物、进化上高度保守的丝氨酸/苏氨酸类蛋白激酶。MAPK基因广泛参与植物细胞的信号转导过程,并且能够响应植物多种生物和非生物胁迫[25]。目前,在拟南芥[26]、苜蓿[27]、水稻[28]等多种植物中已发现大量MAPK基因,但在小麦中关于MAPK基因的研究比较滞后,尤其是氮胁迫方面的研究。郝琳[29]对小麦3个MAPK基因TaMAPK3、TaMAPK4和TaMAPK12;1的耐逆性进行研究,结果表明,在低氮逆境下,3个基因都产生明显应答,然而它们的表达情况却不尽相同,TaMAPK3和TaMAPK12;1基因在低氮胁迫下表达量下降,TaMAPK4基因在低氮胁迫下表达量增加。
1.4 其他基因
除了上述氮胁迫响应基因外,小麦中还存在一些其他的氮胁迫响应基因。研究发现,小麦G 蛋白β亚基基因TaRACK1对低氮胁迫产生明显的应答,在根和叶中的表达水平明显上调,说明TaRACK1可以作为响应低氮逆境的一个参考基因[30]。同样,Chen等[31]也发现,TaRACK1基因响应低氮胁迫,其表达量上调。另外,袁晓磊等[32]研究表明,在低氮条件下,小麦地上部谷氨酰胺合成酶基因TaGS1c、丙氨酸转氨酶基因TaAlaAT和丙酮酸磷酸双激酶基因TaPPDK的表达水平在抽穗期以后均显著增强,且氮高效小麦品种的表达水平显著高于氮低效小麦品种,表明TaGS1c、TaAlaAT和TaPPDK基因在决定小麦氮利用效率方面发挥重要作用。Shrawat等[33]、Good等[34]也得到类似的结论。
2 磷胁迫下小麦响应基因研究进展
土壤有效磷偏低是全球普遍存在的问题,施用磷肥虽然对农业生产起到一定的作用,但磷肥施入土壤后作物当季利用率仅为10%~25%,近75%~90%积累在土壤中,致使土壤全磷含量较丰富,而有效磷含量很低,从而影响作物生长发育和产量的形成[35-36]。有研究表明,植物对低磷胁迫的响应,在分子水平上是低磷信号转导通路组分在转录和翻译水平协同作用的结果[37]。目前,响应磷胁迫的基因主要有bHLH、CDPK、MAPK等。
2.1 bHLH基因
bHLH转录因子是一类超结构域蛋白家族,在植物和动物体内广泛分布[38]。在植物体内,bHLH转录因子参与对环境信号的感受等生物过程,在调控植物生长发育以及应答和抵御非生物逆境中发挥着重要作用[39]。目前,已发现的参与磷胁迫的小麦bHLH基因主要为TabHLH1[39-40]。
张珉等[40]利用cDNA-AFLP技术鉴定了1个对低磷胁迫产生明显应答的小麦bHLH基因TaHLH1,通过比对分析表明,TaHLH1编码蛋白与水稻、大麦和玉米的同源蛋白在保守基序的氨基酸组成上高度同源;TaHLH1基因对低氮、干旱和高盐等逆境产生明显应答,但对低钾、低钙、脱落酸(ABA)和低温不产生应答。表明TaHLH1在介导低磷等非生物逆境的信号转导中可能发挥着重要作用。刘芳朋[39]利用RT-PCR技术对小麦TabHLH1基因在低磷胁迫下的表达特征进行研究发现,小麦根系中TabHLH1基因对低磷胁迫产生明显应答,且表达水平随着处理时间的延长呈不断增高趋势,在处理9 h时表达量最高;另外,小麦根系中TabHLH1基因对低氮胁迫也产生应答,但是对低钾和低钙胁迫不产生应答。由此推断,小麦TabHLH1转录因子在调控植株应答和抵御低磷、低氮胁迫过程中可能发挥着重要作用。
2.2 CDPK基因
CDPK是目前植物中研究较多、了解较为清楚的一类丝氨酸/苏氨酸蛋白激酶,在植物发育及生物和非生物胁迫信号转导中执行不同的生物学功能[41-42]。在小麦响应低磷胁迫方面,路文静等[43]鉴定了2个应答低磷胁迫的小麦CDPK基因TaCPK1A和TaCPK10,在低磷胁迫1~24 h时根系中TaCPK1A基因响应明显,且表达水平不断增强,叶中该基因在1 h内明显被诱导表达,之后表达水平保持稳定;在低磷胁迫条件下,TaCPK10基因在根、叶中的表达水平均呈低—高—低的变化趋势,在胁迫1 h时表达被诱导,之后表达量又逐渐降至胁迫前水平;另外,TaCPK1A和TaCPK10基因对氮、钾胁迫无应答。此外,TaCPK1和TaCPK5基因对低磷胁迫也产生应答,表达上调[44]。这暗示着有些CDPK基因可能参与小麦植株对低磷逆境信号的转导、响应或抵御过程。
2.3 MAPK基因
MAPK基因不仅对低氮胁迫产生应答,对低磷胁迫也产生应答。TaMAPK3、TaMAPK4和TaMAPK12;1基因除了对低氮逆境产生明显应答外,对低磷等非生物胁迫也产生响应[29]。在低磷胁迫下,TaMAPK3基因表达量下降,TaMAPK4和TaMAPK12;1基因表达量增加,其中TaMAPK3和TaMAPK4基因的表现与低氮环境相同,说明TaMAPK3和TaMAPK4基因可能在氮胁迫和磷胁迫交互响应方面起重要作用。另外,MAPK可调控下游基因的表达,在低磷胁迫下,转基因烟草植株中部分磷酸盐转运蛋白基因的表达水平发生改变,表明这些基因可能通过对下游磷酸盐转运蛋白基因转录过程的调控,来调节植株抵御磷胁迫的能力[29]。路文静等[45]研究响应低磷胁迫的小麦MAPK基因发现,TaMPK1a-1基因在小麦根、叶中均受到明显诱导,表明该基因可能会增强小麦适应低磷胁迫的能力。文延丽等[46]克隆了1个小麦MAPK基因TaMAPK7-1,该基因在低磷和高盐胁迫下上调表达,但在低氮和干旱胁迫下表达量没有发生变化,表明小麦MAPK基因可能通过增强表达及进一步磷酸化过程在介导植株应答和抵御低磷及盐分胁迫中发挥重要作用。
2.4 其他基因
磷饥饿诱导(IPS)基因已经分别在蒺藜状苜蓿、拟南芥和水稻中被克隆,并发现其只有在缺磷情况下才能表达,氮素缺乏以及干旱、低温和高盐胁迫均不能诱导IPS基因的表达[47-49]。这使得IPS基因成为一个理想的诊断植物缺磷的标记基因。李彦龙等[50]研究发现,缺磷显著增加了小麦根系中3个TaIPS1基因和地上部TaIPS1.1基因的表达,中度上调了根系中2个TaIPS2基因的表达,轻度上调了地上部TaIPS1.2和2个TaIPS2基因的表达。通过比较,认为TaIPS1.1是一个较理想的用于诊断小麦植株磷素丰缺的基因。方韦博等[51]在丰磷条件下小麦根系cDNA差减文库中鉴定了1个依据酵母蛋白和人类逆转录病毒受体蛋白共有的保守基序命名的基因TaSPX129,并在非生物逆境下对其进行研究发现,TaSPX129基因对低磷逆境产生明显应答,对低氮和高盐胁迫也产生明显应答。初步说明TaSPX129基因在介导植物适应和抵御上述非生物逆境中可能发挥着重要的作用。在小麦中,对低磷胁迫产生明显应答的基因还有转录因子基因PTF、锌指蛋白基因ZAT10、参与磷素吸收的磷转运蛋白基因PHT3和PT2[52]等。
3 展望
氮、磷是作物生长所必需的营养元素,生产中每年都会投入大量的肥料来满足作物的生长需要,同时也引起了一系列问题。所以,从作物营养高效与生态环境保护的角度看,无论是氮肥、磷肥还是其他肥料的投入,都应该紧紧围绕“高效、生态、安全”的目标进行。揭示小麦养分胁迫相关响应基因及其调控机制,从基因表达水平来反映养分的亏缺情况,将会极大促进对小麦养分高效吸收利用机制的理解,对其中关键基因进行遗传操作将有可能培育养分高效利用的新品种,这样不仅在遗传资源的收集、高效品种的筛选,而且在提高肥料效益、经济效益和生态效益方面具有至关重要的作用。目前,关于小麦氮、磷胁迫响应基因的研究已经取得了一些进展,但相比于拟南芥、水稻等模式作物,在小麦上的研究相对较少。另外,非生物胁迫因子的研究多集中于干旱、低温及盐胁迫,对氮、磷胁迫的报道相对较少。目前所掌握的小麦氮、磷胁迫响应基因并不是很多,还需要进一步对相关基因进行挖掘。小麦的基因组庞大且复杂,目前已鉴定出多种氮、磷胁迫响应基因,但是哪种基因是关键应答基因还需要进一步探讨。
