松嫩草地豆类草群落土壤细菌碳源利用研究
2018-07-28,,,,
, , , ,
(1. 吉林农业大学,吉林 长春130118;2. 黑龙江八一农垦大学,黑龙江 密山 163319; 3. 东北师范大学植被生态科学教育部重点实验室,吉林 长春 130024)
草原生态系统中,植物是物质循环和能量流动的中枢,土壤细菌则是生物地球化学循环的重要驱动者,植物与土壤细菌之间的联系较为复杂,植物可以通过改变土壤的物理化学条件对细菌群落的代谢活性和代谢多样性产生一定影响,土壤细菌可以迅速响应并适应周围环境的变化,调节植物生长以及植物营养元素活性等方面[1-3]。而在一定条件下,土壤细菌的多样性则可以反映出土壤环境的历史情况,在草地生态系统中具有重要作用[4-6]。
豆科植物在草地生态系统中具有重要生态功能,在改良土壤的结构、提高土壤的肥力和促进植被演替等方面具有一定的功能作用[7-8]。土壤细菌群落多样性与生态系统营养之间密切相关[9]。有研究表明,豆科植物在一定程度上可以改善土壤细菌的功能,提高其群落的代谢多样性[1,10]。
通过Biolog-ECO微平板培养技术可描述土壤细菌功能多样性与代谢活性,从土壤细菌对单一碳源的利用程度来进一步反映土壤细菌群落水平的生理轮廓[11-13]。我们曾对松嫩草地羊草群落土壤细菌碳源利用进行了研究,关于豆类草群落与土壤细菌多样性关系的研究还很少[14]。本试验以盐碱化严重,且羊草占主要优势的松嫩放牧草地为背景,探讨了草木犀和胡枝子两种豆类草群落对土壤细菌碳源利用的影响,同时与羊草群落进行了比较。研究了不同植物群落与土壤细菌之间的关系,为草地可持续发展道路提供更多的理论依据。
1 材料与方法
1.1 试验小区概况
试验地点为位于松嫩草地的西南部的东北师范大学草地生态研究站(35°23.1′~35°34.1′ N,123°30′20.5″~123°30′45.0″ E)。羊草(Leymuschinensis)和杂类草群落为草地的主要群落类型,这类群落主要分布于土壤碱化比较严重的地段。除优势种羊草外,还有亚优势种芦苇(Phragmitisaustralis),及其他伴生种如虎尾草(Chlorisvirgata)、拂子茅 (Calamagrostisepigeios)等,在植物群落间形成镶嵌分布的格局,间有兴安胡枝子(Melilotusdaurica)和草木犀(Melilotusofficinalis)等组成豆类草群落分布[15]。
1.2 试验方法
1.2.1植被调查 植被调查于2014年8月牧草生长最为旺盛时期进行。样地大小为1 m×1 m,重复3次。分别记录植物种类、高度、盖度、密度以及生物量等数据。
1.2.2土壤样品采集 在试验小区内按照羊草、草木犀和胡枝子群落取样,采取“五点混合法”取土壤样品,三次重复。土钻直径为2 cm,取地面下0~20 cm土壤。除去土样中杂质,5 点混匀后将样品装入无菌塑料袋中,分为2份带回实验室。一份置于-20℃冰箱保存,用于进行Biolog实验;另一份在室内进行风干,用于测定土壤理化性质。
1.2.3土壤理化性质的测定 烘干称重法测定土壤含水量;用雷磁PHS-3C pH计测定土壤pH ;用DDS-307电导仪测定土壤电导率;凯氏定氮法测定土壤氮含量;重铬酸钾容量法测定土壤有机质。
1.2.4Biolog-ECO微平板培养 将10 g过2 mm筛的土壤加到90 ml无菌NaCl溶液中,用振荡器混匀(5 min,180 r·min-1),按照10倍稀释法配制成10-3倍土壤稀释液,然后用移液枪注入到ECO微平板中,最后在26℃的培养箱中连续培养10 d,每隔12 h用Biolog仪读数一次。
1.3 数据分析
数据分析软件为Microsoft Excel 2007;方差分析软件为SPSS 17.0;冗余分析(RDA分析)软件为Canoco for Windows 4.5。
土壤细菌代谢强度采用单孔颜色平均值(average well color development,AWCD)来描述。
计算方法为:AWCD=∑(C590-C750)/31
式中: C590为生态板中碳源孔在590 nm下的吸光度值减对照孔,C750为碳源孔在750 nm下的吸光度值减对照孔。
采用Shannon-Wiener指数、Pielou均匀度指数以及Gini多样性指数来描述土壤细菌群落的多样性:
(1)Shannon-Wiener指数(H):H = -Σ(Pi×lnPi)
(2)Pielou均匀度指数(E):EH = H/Hmax = H/lnS
(3)Gini多样性指数(D):D = 1 - ΣPi2;
式中:Pi= C590-C750/∑(C590-C750),S 为 ECO 板中颜色变化孔的数目。
2 结果与分析
2.1 植物群落特征
植物群落中植物组成变化对植物群落特征影响较大,各项指标均存在显著差异(P<0.05)。羊草群落主要由羊草、芦苇、虎尾草、紫花苜蓿(Medicagosativa)等植物组成;草木犀群落主要由草木犀、羊草、芦苇和碱地肤(Kochiasieversiana)等植物组成;胡枝子群落主要由兴安胡枝子、羊草、广布野豌豆(Viciacracca)和全叶马兰(Kalimerisintegrifolia)等植物组成。胡枝子群落植被高度最高,其次为羊草群落,最后为草木犀群落;草木犀群落和胡枝子群落在植被盖度、密度和地上生物量上均小于羊草群落,且二者之间差异不显著(见表1)。
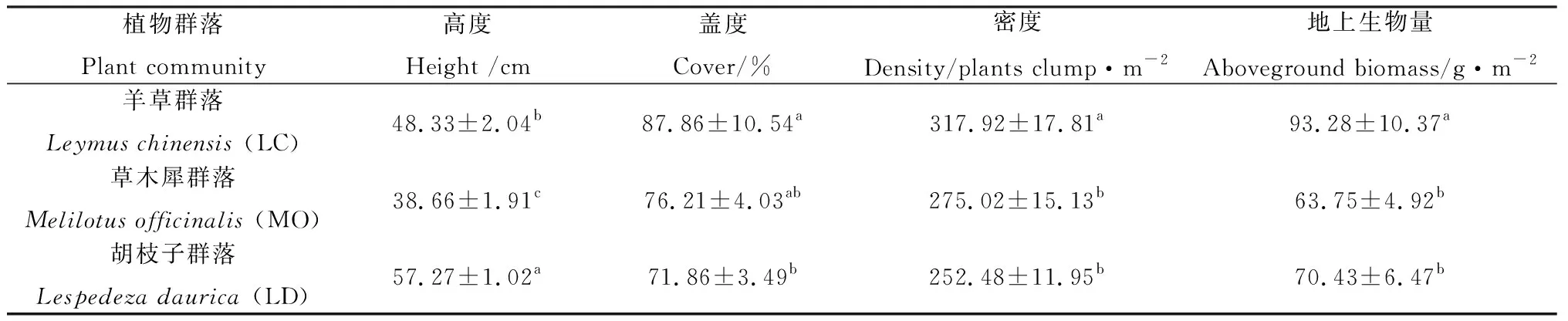
表1 植物群落特征Table 1 Characteristic of plant community
注:同列不同字母表示差异显著(P< 0.05),数据为平均值±标准误,下同
Note: Values with different lowercase letters show significant differences at the 0.05 level, the data is average value ± standard error. The same as below
2.2 土壤理化性质
3种植物群落的土壤理化指标均存在显著差异(P< 0.05)。其中,羊草群落的含水量、电导率和pH均最高;而草木犀和兴安胡枝子群落的有机质和全氮含量则明显高于羊草群落(见表2)。
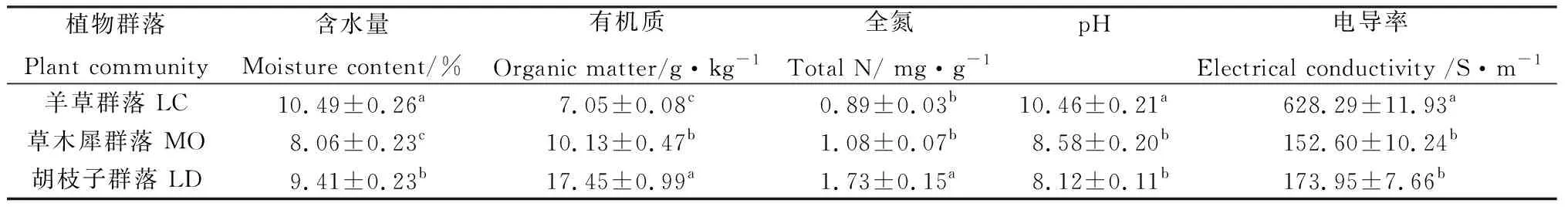
表2 土壤理化性质Table 2 Physicochemical properties of soil
2.3 土壤细菌的平均吸光度值
结果表明,3种植物群落土壤细菌的AWCD值均随培养时间的延长而上升(见图1)。培养前36 h,3种植物群落土壤细菌的AWCD值变化不明显;96~180 h之间土壤细菌的代谢活性增加,消耗碳源的速度加快;180 h后,平缓增长,代谢功能下降。兴安胡枝子群落土壤细菌的AWCD值上升较快,且增长幅度明显高于羊草和草木犀群落;而羊草群落和草木犀群落则上升较为缓慢且增长情况相近。兴安胡枝子群落土壤细菌的AWCD值最高,说明其土壤细菌对碳源利用情况最好。而草木犀群落的AWCD值与羊草群落接近,但稍低于羊草群落,说明二者的土壤细菌代谢情况与活性接近。

图1 土壤细菌平均吸光值变化曲线Fig.1 Curve of AWCD of soil bacteria
2.4 土壤细菌生理碳代谢指纹图谱分析
从Biolog代谢指纹图谱可以更加直接和全面地了解土壤细菌群落代谢功能。结果表明,松嫩草地羊草群落土壤碳源利用的数量最少,对L-天门冬酰胺、D-甘露醇和吐温等4种碳源的利用程度最好,对2-羟基苯甲酸和衣康酸等8种碳源几乎没有利用;草木犀群落土壤碳源利用的情况好于羊草群落,对D-甘露醇、D-纤维二糖和L-丝氨酸等8种碳源的利用程度较好,对D, L-α-磷酸甘油等4种碳源几乎没有利用;胡枝子群落土壤碳源利用数量最多,大多数碳源利用情况较好,只对γ-羟丁酸、α-环式糊精和苯乙胺这3种碳源几乎没有利用(见图2)。
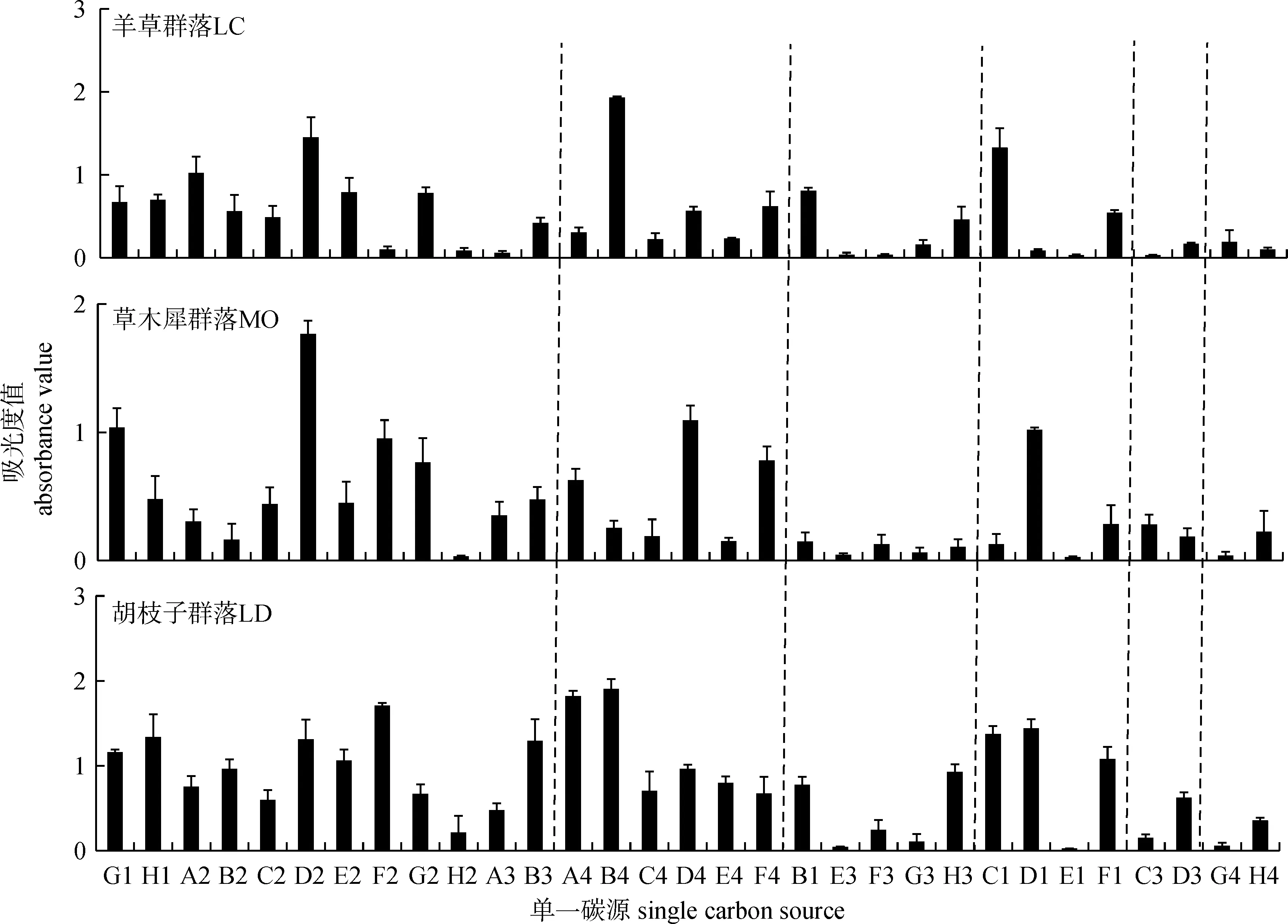
图2 土壤细菌生理碳代谢指纹图谱Fig.2 Metabolic fingerprint of carbon physiological profiles of soil bacteria注:G1-B3为糖类,包括G1.D-纤维二糖;H1.a-D-乳糖;A2.β-甲基-D-葡萄糖苷;B2.D-木糖;C2.I-赤藻糖醇;D2.D-甘露醇;E2.N-乙酰基-D-葡萄胺;F2.D-氨基葡萄糖酸;G2.α-D-葡萄糖-1-磷酸;H2.D,L-α-磷酸甘油;A3.D-半乳糖酸内脂;B3.D-半乳糖醛酸;A4-F4为氨基酸类,包括A4.L-精氨酸;B4.L-天门冬酰胺;C4.L-苯基丙氨酸;D4.L-丝氨酸;E4.L-苏氨酸;F4.甘氨酰-L-谷氨酸; B1-H3为羧酸类,包括B1.丙酮酸甲酯;E3.γ-羟丁酸;F3.衣康酸;G3.α-丁酮酸;H3.D-苹果酸;C1-F1为多聚物类,包括C1.吐温;D1.吐温80;E1.α-环式糊精;F1.肝糖;C3-D3为酚酸类,包括C3.2-羟基苯甲酸;D3.4-羟基苯甲酸;G4-H4为胺类,包括G4.苯乙胺;H4.腐胺
2.5 土壤细菌对不同碳源的利用
Biolog-ECO微平板的31种碳源分6类,分别是糖类、氨基酸类、羧酸类、多聚物类、酚酸类和胺类。对180 h的AWCD值进行6类碳源利用分析,胡枝子群落对6类碳源的利用情况均高于其他两种植物群落;羊草群落对糖类和氨基酸类利用较好,对酚酸类利用最差;草木犀群落对羧酸类利用最差;3种植物群落土壤细菌对氨基酸、糖类等的利用程度较好,对胺类的利用较差(见图3)。
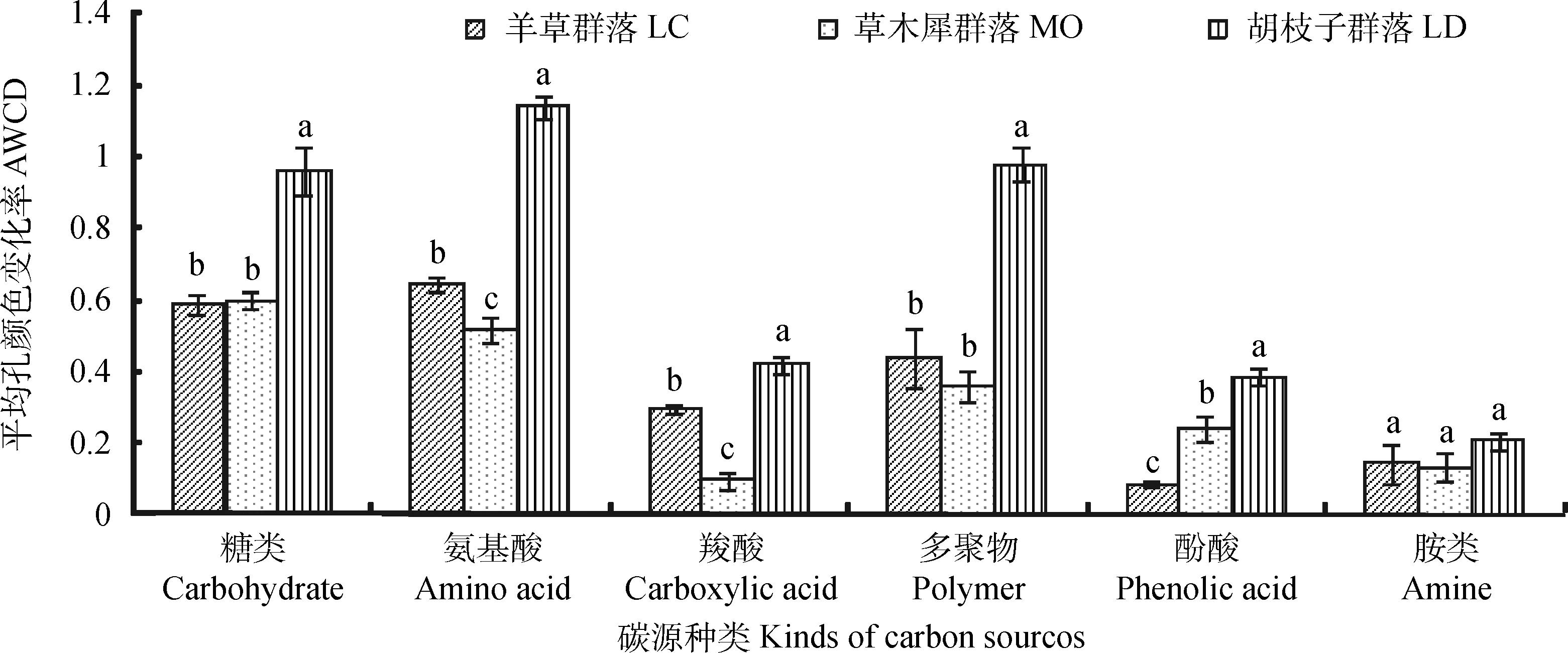
图3 土壤细菌对六类碳源的利用情况Fig.3 AWCD change of the soil bacteria with 6 carbon sources
2.6 土壤细菌碳源利用多样性指数
对180 h的AWCD值进行多样性指数分析,不同植物群落下土壤细菌群落的碳源利用多样性指数不同,胡枝子群落的Shannon-wiener指数,Pielou均匀度指数以及Gini多样性指数最高,说明其多样性与均匀度最好;草木犀群落的多样性与均匀度最低(图4)。

图4 土壤细菌碳源利用多样性指数Fig.4 Diversity index of carbon source metabolism of soil bacteria
2.7 土壤细菌碳源利用与土壤理化指标冗余分析
对180 h的AWCD值进行细菌碳源利用的RDA分析,可以看出不同植物群落土壤细菌碳源利用与土壤理化指标关系(见图5)。各个样方按照植物群落不同而呈有规律分布,碳源利用差异主要由植物群落决定。所有土壤变量可以解释土壤细菌群落变化总变量的78.2%(蒙特卡罗置换检验999,P= 0.01),第1轴作为约束轴能解释环境变量的37.8%,综合箭头方向和长度分析,土壤的有机质和全氮含量是影响这3种植物群落土壤细菌碳源利用的最主要因素,而从这2种环境因素分析对3种植物群落对碳源利用情况大小排序依次为,胡枝子群落、草木犀群落和羊草群落。
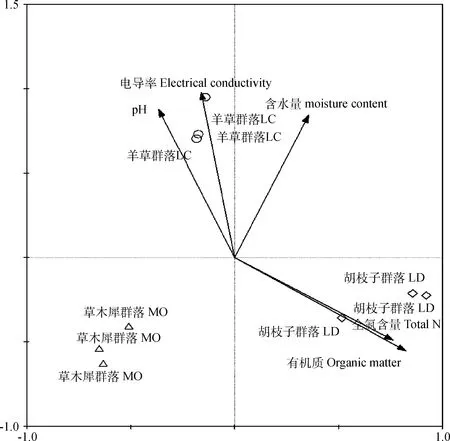
图5 土壤细菌31孔吸光度值与土壤理化性质冗余分析Fig.5 Redundancy analysis of the 31 hole absorbance value of soil bacteria and physicochemical properties of soil(RDA)
3 讨论
松嫩草地的优势种羊草占的比例最多,也分布一些豆科植物和杂类草,但其中羊草的营养价值高、适口性良好等一系列特征导致在放牧过程中被消耗的最多[15-16]。经过短期放牧后,出现了胡枝子和草木犀为代表的2个主要豆科植物群落,在盖度、密度以及地上生物量上均比羊草群落低,植物群落丰富程度略低,说明放牧改变了草地植物群落的组成。
豆科植物可以作为绿肥,提高土壤含氮量,且在植物生长发育过程中,细根及根瘤的死亡脱落,以及枯落物的腐解等,同样使得土壤中有机质的含量升高[17]。3种植物群落的土壤理化指标同样差异明显,其中2种豆科植物群落土壤的有机质与全氮含量高于羊草群落。RDA分析表明,影响植物群落土壤细菌碳源利用的最主要因素为土壤的有机质含量及全氮含量。从中可以看出,豆科植物会对土壤条件有显著的改善作用,从而进一步影响土壤细菌的碳源代谢情况。
Biolog-ECO微平板技术可以获得大量数据和有价值的信息,是研究土壤细菌群落功能多样性的简单、有效的主要方法[18]。土壤细菌利用单一碳源的AWCD值均随时间延长而上升,与土壤细菌的代谢情况呈相近的趋势。Biolog-ECO微平板培养土壤细菌的整体培养时间通常为10 d,共240 h。0~24 h为土壤细菌适应微平板环境的滞后期;24 h后进入快速增长的指数期;通常在156~192 h之间会有一个拐点时间,使其进入生长的平缓期[19-20]。本试验的3种植物群落土壤细菌的AWCD值随时间变化曲线符合这一生长规律。
地上植物不同,土壤碳源种类和数量也可能不同,土壤中存在的代谢不同碳源的细菌在数量和分布上也不相同。Biolog-ECO生态板中,至少有9种碳源是根系分泌物的组分,因此不同植物群落的土壤细菌对碳源的利用程度存在差异[21-23]。胡枝子群落土壤细菌利用碳源的数量和种类最多,对糖类、氨基酸和多聚物这3类碳源能进行很好的利用,对胺类的利用率最低;羊草群落和草木犀群落土壤细菌利用的碳源数量较少,对碳源利用好的种类也较少。说明在胡枝子群落中,土壤中所含有的代谢不同碳源的细菌的类型和数量较多。
多样性指数在反映土壤细菌的群落组成以及个体数量分布上有一定作用,通过土壤细菌群落的多样性和均匀度,可以进一步反映土壤细菌群落的组成结构及其稳定程度[24-25]。胡枝子群落土壤细菌群落的多样性指数最高,说明具有较高代谢活性细菌的种类和数量较多。已有研究表明,豆科植物可增加土壤微生物的数量和代谢多样性[10]。在松嫩放牧草地中,豆科植物在改善草地植物格局、提高饲草产量和质量等方面具有重要的生态功能[16]。
4 结论
综上所述,不同植物群落组成及其地下土壤理化指标导致了土壤细菌碳源利用的不同,不同植物群落土壤细菌群落功能多样性有很大差异。豆类草群落下的植被盖度、密度和地上生物量均小于羊草群落,但豆科植物的固氮作用导致有机质和全氮含量明显增加,改善了土壤条件,使得土壤细菌碳源利用较好。2种豆科植物群落植物组成不同,对土壤碳源利用存在差异,反映了其土壤细菌代谢能力及活性不同。土壤的有机质和全氮含量是影响土壤细菌碳源利用的最主要因素。
