封育年限对伊犁绢蒿荒漠土壤微生物及酶活性的影响
2018-07-26孙宗玖李培英江沙沙
孙宗玖, 李 琦, 李培英, 江沙沙
(1.新疆农业大学 草业与环境科学学院, 新疆 乌鲁木齐 830052; 2.新疆草地资源与生态重点实验室, 新疆 乌鲁木齐 830052)
新疆地处亚欧大陆腹地中温带干旱荒漠区,可利用天然草地面积4.80×107hm2,划分为11个草地类,其中荒漠草地是其重要组成部分,面积约为2.69×107hm2,生产中多作为春秋场或冬场利用,在季节牧场轮换利用中具有重要的地位[1]。以伊犁绢蒿(Seriphidiumtransiliense)为建群种的荒漠是新疆荒漠草地的典型代表,面积约为1.14×106hm2,广泛分布于新疆天山北麓山前洪积扇和低山丘陵区[2]。由于所处生态环境脆弱、放牧压力大等原因,伊犁绢蒿荒漠退化现象十分普遍,并成为新疆草地退化的重灾区,对草地生物多样性的保护、畜牧业生产和绿洲生态安全造成了严重威胁,因此恢复与治理退化的伊犁绢蒿荒漠已成为当前亟待解决的首要问题之一。
围栏封育已成为新疆退化草地恢复的重要举措,可使退化的草地的植被、土壤得到一定程度的改善[3]。目前,围绕封育过程中退化伊犁绢蒿荒漠恢复效果已开展了大量研究[2,4-6],主要集中在草地植被群落特征及其多样性、土壤常规养分及土壤碳组特征等方面的对比分析,相对而言对其土壤微生物组成数量特征及土壤酶活性的探讨较少。土壤微生物和土壤酶是衔接土壤有机物—无机物—生物相互作用的重要参与者与推动者,在土壤有机物分解、腐殖质的形成、养分转化等环节中具有重要作用[7-8],因此研究封育过程中土壤微生物和土壤酶的响应机制对深入了解退化草地土壤质量的恢复具有重要指导意义。基于以上认识,本文拟以轻度退化的伊犁绢蒿荒漠为对象,通过对不同封育年限下草地土壤有机质、土壤微生物数量及土壤酶活性的测定,探讨其对封育年限的响应规律,以期为退化蒿类荒漠草地的改良、复壮以及生态系统的恢复提供参考依据。
1 材料与方法
1.1 研究区概况
研究区位于新疆维吾尔自治区乌鲁木齐市米东区芦草沟乡的博格达山北坡低山带(87°47′—87°46′E,43°53′—43°49′N),海拔840~1 110 m。试验区为典型温带大陆气候,四季明显,冬季寒冷漫长,年均温6.4 ℃;年降水量约为236 mm,四季分配不均,春季占全年降水量的30%。土壤类型为灰漠土,土层较厚。该草地主要优势植物有伊犁绢蒿(S.transiliense)、木地肤(Kochiaprostrata)、叉毛蓬(Petrosimoniasibirica)、角果藜(Ceratocarpusarenarius)等,每年春(3月中旬至6月中旬)、秋(9月上旬至11月中旬)两季放牧利用,且地表有一定程度的裸露,总体处于轻度退化状态。
1.2 样地选择
试验设置2个封育处理,即封育3 a和11 a,分别在2013,2005年采用网围栏进行封育,面积均为2 000 m2,且封育区内全年禁止放牧利用。封育区基本相连(小于10 m),且土壤状况、地形地貌、降水、温度等生态因子基本保持一致;同时封育区外围设为对照区(CK),为传统的自由放牧区,放牧畜种多为新疆细毛羊。
1.3 土壤样品采集
2016年5月,每个围栏样地沿坡向平均分为3个条带区,每个条带区内沿对角线采用蛇形取样(Z字型),每隔10 m设置1个土样取样点,共5处。每处用直径为7 cm的土钻,按土层深度0—5,5—10,10—20 cm分层钻取2钻,同层混匀形成混合样,放入做好标签的自封袋冷藏带回室内。对照区则在封育区附近随机设置样地3处,按相同方法获取土样。室内,剔除植物根系、石砾等杂物后,将混合样分成2部分,1部分放置4 ℃冰箱贮存,用于土壤微生物数量的测定,另1部分置于室内自然风干,磨碎,过0.25,1,2 mm筛后保存,用于土壤有机质及酶活性的测定。
1.4 测定方法
土壤有机质采用重铬酸钾外加热法[9]。过氧化氢酶活性测定采用高锰酸钾滴定法,脲酶活性测定采用苯酚钠比色法[10]。土壤微生物数量测定采用稀释平板法,细菌用牛肉膏蛋白胨琼脂培养基,真菌用马丁氏孟加拉红琼脂培养基,放线菌用淀粉铵盐琼脂培养基培养法[11]。
1.5 数据分析
利用SPSS 17.0数据统计软件的one-way ANOVA及Duncan法进行相关数据的统计分析,结果以“均值±标准误”的形式表示;利用Microsoft Excel 2013进行相关表格的制作。
2 结果与分析
2.1 封育对土壤有机质的影响
表1看出,随封育年限增加,0—5 cm土层有机质含量呈上升趋势,5—10 cm,10—20 cm土层则呈先降后升趋势,封育3 a最低。与对照比,封育3 a和11 a的0—5 cm土层有机质含量分别增加7.7%,14.2%(p<0.05),而封育3与11 a间差异不显著;5—10 cm土层封育3 a有机质含量下降了6.6%(p>0.05),至封育11 a则时增加15.4%(p<0.05);10—20 cm土层有机质增降不明显。随土层深度增加(表1),0—20 cm土层有机质含量呈逐渐下降趋势,且0—5 cm土层显著高于5—20 cm土层,且至封育11 a时,5—10 cm土层有机质才显著高于10—20 cm土层(p<0.05)。

表1 封育年限对伊犁绢蒿荒漠土壤有机质的影响g/kg
2.2 封育对土壤微生物数量的影响
表2看出,随封育年限的增加,伊犁绢蒿荒漠0—20 cm土层微生物总数量均呈降低趋势,但各封育处理间差异不显著,且无论封育与否,土壤微生物数量构成上均以细菌最高,放线菌次之,真菌最低,如封育3 a样地,0—5 cm土层中细菌、放线菌依次占其微生物总数的85.1%,14.7%,而真菌仅为0.02%。进一步对土层中不同微生物种类组成的分析发现(表2),封育年限对不同土层的细菌、放线菌、真菌数量的影响并不一致,需要区别对待。0—5 cm土层,随封育年限的增加,细菌、真菌数量呈降低趋势,放线菌数量呈增加趋势,且至封育11 a细菌、真菌较对照依次显著降低47.3%,25.5%,而放线菌则显著增加15.5%(p<0.05);5—10 cm土层,放线菌数量封育3 a显著高于对照和封育11 a,而真菌数量则对照、封育3 a显著高于封育11 a(p<0.05),但封育对细菌数量影响不显著;10—20 cm土层,放线菌、真菌数量随封育年限增加呈增加趋势,封育3,11 a间差异不显著,且均显著高于对照(p<0.05),而封育对细菌数量影响仍不显著(p>0.05)。随土层深度的增加(表2),封育3 a及11 a的5—10,10—20 cm土层细菌、放线菌、真菌数量及其微生物总量均显著高于0—5 cm土层,而5—10,10—20 cm土层间差异不显著;对照区5—10 cm土层放线菌、真菌数量则显著高于0—5,10—20 cm土层(p<0.05),而细菌及微生物总数则各土层间差异不显著(p>0.05)。
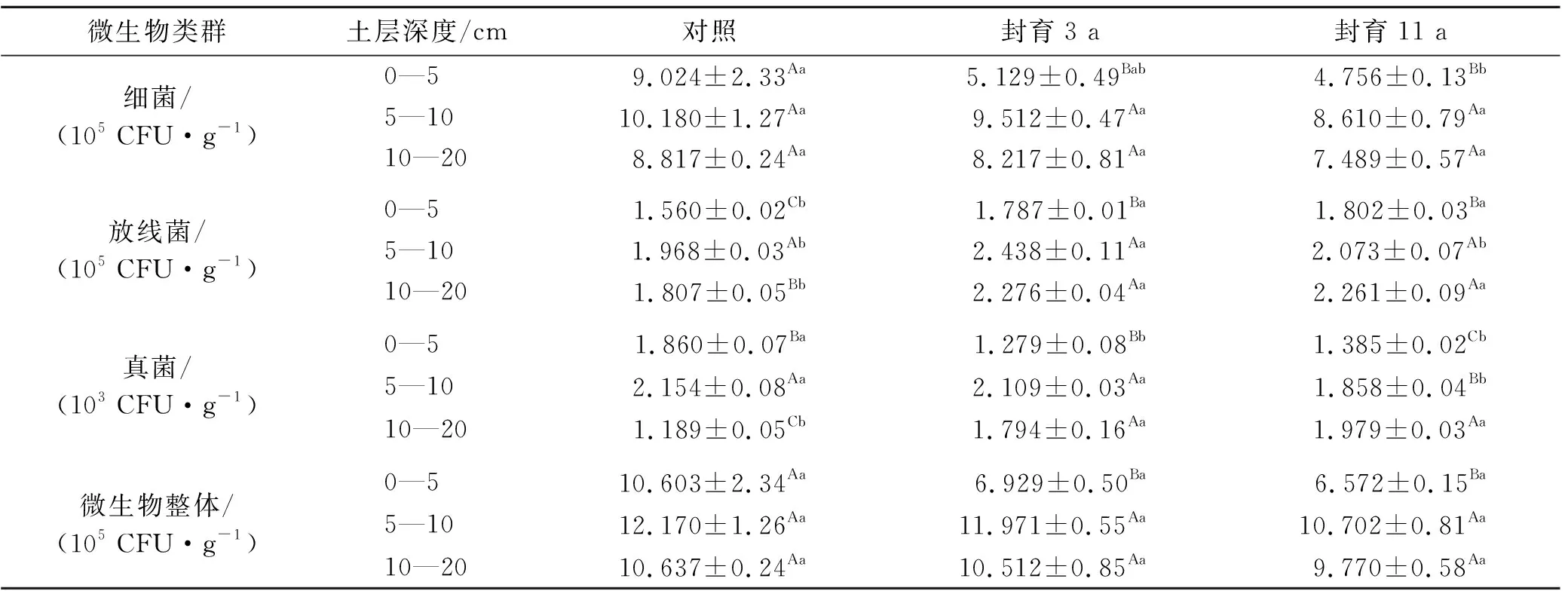
表2 封育年限对伊犁绢蒿荒漠土壤微生物数量的影响
2.3 封育对土壤酶活性的影响
表3看出,随封育年限增加,0—20 cm土层过氧化氢酶活性呈下降趋势,但仅封育11 a的10—20 cm土层过氧化氢酶活性较对照显著降低,降幅为26.1%(p<0.05),而封育3 a处理则介于两者之间;0—10 cm土层脲酶活性呈增加趋势,且封育11 a较对照显著增加15.9%~41.5%,而10—20 cm土层脲酶活性呈现降低,且封育11 a较对照显著降低39.7%。随土层深度增加,过氧化氢酶活性呈上升趋势,且10—20 cm土层高于0—5 cm土层(p<0.05),而封育后脲酶活性则呈现降低,且10—20 cm土层基本低于0—5 cm土层(p<0.05)。

表3 封育年限对伊犁绢蒿荒漠土壤酶活性的影响
3 讨 论
退化草地封育后可显著提高草地植物群落的高度、盖度,并在封育5~10 a后达到最高,但土壤质量恢复较为缓慢,短期内变化不明显,只有经过长时间封育才会得以显著恢复,且其恢复效果与封育区域所处环境及封育前草地的本底直接相关[12]。Hu等[13]整合分析了中国326个封育样地的监测数据,认为封育后92%的退化草地土壤有机碳出现增加,8%土壤有机碳出现降低或变化不明显。如李丽君等[14]指出,草地封育26 a后天山中部高寒草甸、高寒草甸、高寒草原表层土壤有机碳分别提高11.37%,3.26%和2.21%,但仅在高寒草甸出现显著增加,而杨新国等[15]认为,短期封育条件下荒漠草原沙化灰钙土有机碳含量变化不显著。本研究结果表明,退化草地不同土层有机质对封育的恢复响应存在一定的差异。与对照比,退化伊犁绢蒿荒漠封育3 a后0—5 cm土层有机质呈现显著增加,而5—10 cm土层则出现不明显降低,且封育3 a样地的有机质显著低于封育11 a样地,但10—20 cm土层则至封育11 a土壤有机质仍增加不显著。初步说明退化草地土壤有机质的恢复可能按照先表层后深层的顺序逐步有序恢复,且长期封育均利于土壤有机质的积累。同时本研究结果也与杨合龙等对轻度退化蒿类荒漠[16],董乙强等[5]对中度退化蒿类荒漠及杨静等[17]对重度退化的沙质荒漠的研究结果“封育早期(1~4 a)草地土壤有机质含量均会出现不同程度的降低,后期才逐渐增加”基本吻合,说明退化荒漠草地植被初期恢复过程中地上植被的快速复壮,净生产力增加过多,可能会导致植物对土壤养分的吸收量远远高于其返还量,同时干旱环境下返还的凋落物并不能及时分解所致,封育后期土壤有机质的提高可能通过逐年地上植被净生产力(凋落物、死的根系)的积累,及外界小环境尤其是土壤水分条件的改善加速了土壤有机质输入所致,同时也与封育后截留了放牧条件下家畜将植物碳向系统外的输出有关。
土壤微生物是土壤养分循环和转化过程中的重要组成部分,影响着植物对养分的吸收与利用[18]。目前,有关草地土壤微生物的研究已有较多研究,多集中在微生物种类组成及其数量特征方面[19-22],但研究结果存在一定争议。如姚拓等[19]、柴晓红等[22]分别对天祝高寒草地、夏河县桑科高寒草地土壤微生物数量的研究均表明,细菌最多,放线菌次之,真菌最少,且封育地大于放牧地。郭继勋等[21]认为吉林省长岭县羊草草原上羊草群落、杂类草群落、碱茅群落、碱蓬群落的土壤微生物均以放线菌数量最多,而榆树疏林群落以真菌数量最多,但赵吉等[23]则认为锡林河流域羊草草原土壤微生物数量以细菌最多,放线菌次之。本研究结果表明,无论封育与否,伊犁绢蒿荒漠土壤微生物数量以细菌最多,放线菌次之,真菌最少,这与前人的研究结果相一致[19,22-23],同时也验证了范燕敏等[20]认为,退化伊犁绢蒿荒漠土壤微生物数量以细菌为主的研究结果,但与郭继勋等[21]研究结果不一致。可能是由于研究区所处生境条件及植被群落类型的差异导致其生土壤理化性质的差异所致,伊犁绢蒿荒漠气温相对较高,而生境土壤pH值为微碱性(8.5~8.7),均有利于细菌及放线菌的生长繁育。伊犁绢蒿荒漠0—20 cm土层微生物总量虽然随封育年限增加呈不显著性降低,但封育显著影响了细菌、放线菌及真菌的数量组成比例。如封育11 a 0—5 cm土层细菌、真菌较对照显著降低25.5%~47.3%,而放线菌却显著增加15.5%;5—20 cm土层封育对细菌数量影响不显著,而对真菌、放线菌数量影响显著。这可能与土层温度变化密切相关,封育后伊犁绢蒿荒漠植被盖度增加,表层土壤温度出现一定程度的降低,而真菌、细菌及放线菌生长繁育所需最适温度并不一致,导致其种群消长存在较大差异所致。高寒草原的研究表明[19],封育后细菌、放线菌及真菌数量均出现不同程度的增加,这与本研究结果存在一定差异,可能与其研究区域所处生境条件及土壤有机质含量的高低相关。从土层垂直变化看,无论封育与否,5—10 cm土层细菌、放线菌、真菌数量及其微生物总量均显著高于0—5 cm土层,可能是表层土温相对较高,土壤含水量少,抑制了微生物正常生长发育所致。
土壤酶是土壤的组成部分,可作为生物催化剂,在土壤物质循环和能量转化过程中起着重要作用。一般认为,土壤酶在很大程度上起源于土壤微生物,也可能来源于植物和土壤动物[24]。与对照比,封育后伊犁绢蒿荒漠0—20 cm土层过氧化氢酶活性呈下降趋势,且仅封育11 a的10—20 cm土层呈现显著降低,而0—10 cm土层脲酶活性呈现增加,封育11 a增加显著。其原因可能是: ①封育后引起土壤理化性质的变化,如土壤温度、水分、pH值等,导致土壤微生物数量及种类的变化,进而引起其分泌酶类的差异。 ②封育后伊犁绢蒿草地组成植物种类成分、结构和生物量变化引起其向土壤输入的凋落物及植物残体的化学计量学特征发生变化,导致土壤酶的种类及活性可能也会产生差异。
4 结 论
长期封育有利于退化伊犁绢蒿荒漠土壤有机质的积累,且封育后0—5 cm土层有机质的增加速率高于5—20 cm土层,封育3 a的0—5 cm土层有机质含量就呈现显著增加。
伊犁绢蒿荒漠土壤微生物数量以细菌最多,放线菌次之,真菌最少,且封育对3种微生物数量的影响并不一致。随封育年限的增加,0—5 cm土层细菌、真菌数量呈降低趋势,放线菌数量呈增加趋势,5—20 cm土层细菌数量变化不显著,而放线菌及真菌数量多呈增加趋势。从整体看,封育对土壤细菌、放线菌、真菌的总体数量影响不显著。
封育对伊犁绢蒿0—20 cm土层过氧化氢酶活性影响不显著,而0—10 cm土层脲酶活性随封育年限增加呈增加趋势,10—20 cm土层则呈降低趋势。
