LED株间补光对日光温室番茄产量及光合作用的影响
2018-07-25闫文凯张雅婷张玉琪杨其长
闫文凯,张雅婷,张玉琪,杨其长,李 涛
(中国农业科学院 农业环境与可持续发展研究所,北京 100081)
光照是植物光合作用的动力源,也是农作物健康生长和产量形成的基础。光对植物的形态建成、生理代谢、品质形成有重要的调节作用[1]。寡日照通常为限制日光温室蔬菜作物高产稳产的主要因素[2]。弱光环境通常导致植物叶片净光合速率下降,影响植株光合产物的合成与积累,其下降程度受CO2浓度、大气温度、相对湿度等环境因子共同作用,也与品种自身的生理特性差异有关[3]。由于弱光环境下光合作用受限,植株干物质积累受到抑制,进而导致花芽分化延迟,开花节位提升,对茄果类作物(如番茄)的开花授粉和坐果都造成严重影响,最终导致产量下降[4-5]。同时,弱光条件下植株干物质分配也会发生变化,弱光会导致向叶片分配的光合产物比例轻微增加,这是以降低向果实干物质分配为代价的[6],植株对弱光的响应会直接或间接影响植株的生长及产量、品质的形成。弱光也会影响植株形态,弱光条件下叶片变大变薄,叶色变淡,叶面积增大,以达到截获更多光能的目的。因此,提供适宜、充足的光照环境是植物健康生长及产量形成的基础和保障。
随着日光温室在中国北方的广泛应用,基于日光温室光环境的研究也越来越多。日光温室具有良好的蓄热保温和节能高效特性[7],有效利用日光温室能够延长蔬菜作物生育期,在一定程度上解决了冬季低温对蔬菜生产的影响。日光温室的东西面和北面为保温蓄热墙,仅有南侧拱形屋面覆盖塑料薄膜接受自然光照,并且在夜间有保温被覆盖,这最大限度地发挥了其保温蓄热的功能,但由于其围护结构、薄膜透光率、太阳入射角等多因子的限制,温室内的光照强度只有室外的60%左右,光照时数也远低于室外。与露地栽培相比,光照强度低、光照时间短、日光温室内南北方向及东西方向光照分布不均且日变化大。同时由于日光温室果菜种植密度较高,植株中下层叶片受到上层叶片的遮挡,光照强度很低,严重限制了植株整体光合产物的合成和转运。株间弱光环境不仅在冬季自然光照较弱的条件下存在,即使在自然光照充足的夏季也存在。研究证实光照强度在植物冠层分布符合朗伯比尔定律[8],即光照强度随着冠层深度的增加呈指数递减。有研究结果显示,黄瓜植株中层叶片接受到的辐照度只有顶部叶片的30%[9]。因此株间弱光是限制植株整体光合作用的障碍之一。此外,中下层叶片成熟度较上层叶片更高,而且番茄果实生长主要依靠光合产物向下输送及就近供给,对于中下层部位的果实成熟,与其相邻的上部叶片较顶层叶片起着更为重要的作用[10]。
人工补光是弥补自然光照不足的有效手段,不仅能够增加作物产量,而且会影响蔬菜作物品质,促进及协调植物器官的生长,提高植株壮苗指数和抗病性[11]。人工补光方式主要有顶部补光和株间补光。而株间补光是将补光光源安装在相邻两垄植物中间。LED发热量较低,因此可以作为良好的株间补光光源。此外,由于人工光源随着照射距离的增加光强衰减明显,因此,株间补光较顶层补光有更高的光源利用效率[12]。与上层叶片相比,中下层叶片受到的光照强度更低,在光合作用光响应曲线中处于光限制区域,因此株间补光比顶层补光对光合速率的提升更高效。近些年,国外学者对温室果菜株间补光研究较多,在温室黄瓜、甜椒等作物中开展株间补光试验,研究其对果实产量的影响,发现株间补光能够显著增加作物早期产量,并且能增加果实的大小[13]及果实数量[12];提高第一穗果的数量及干物质含量[14]。国内学者在温室黄瓜中进行株间补光试验,研究其对果实生长及品质的影响,发现补光处理提高了黄瓜产量,却降低果实中糖分含量[15]。综上所述,前人研究多集中在光照强度、果实产量及品质方面,而对光源红蓝光比例、植株干物质分配、叶片光合作用及光学特性的研究较少。为此,本试验采用红蓝LED光源,开展日光温室番茄生长株间补光试验,以未补光植株为对照,研究不同红蓝光比例(4∶1,7∶1,9∶1)对番茄产量、干物质分配及光合作用的影响。
1 材料与方法
1.1 试验材料
供试番茄品种‘瑞粉882’,种子由瑞克斯旺种子有限公司提供。
1.2 试验方法
日光温室南北跨度10 m,试验区东西长30 m,试验地南北向起垄,基质袋覆土栽培,如图1所示。垄间距为1.6 m,株距20 cm。采用单垄双行栽培模式,行距为0.8 m,种植密度为3.65株/m2。试验番茄幼苗于2016年2月3日定植,采用滴灌供给水肥(营养液),并于中午前打开通风口,控制日光温室内的温湿度。当株高约为50 cm时开始吊蔓,将植株均匀布置于基质袋两侧,日常管理过程中每周去除侧芽,保持顶端优势,并在开花后采用振动器每周授粉2次。当果实直径达到1 cm 时进行疏果,保留长势良好、大小一致的果实,每穗留4个果实。在果实开花成熟期,待成熟果实采摘后及时去除冠层下部老叶。

1.LED株间补光光源;2.基质袋;3.地膜;4.土垄1.LED interlighting light source;2.Cultivation bag;3.Mulch;4.Ridge with compacted soil
待番茄植株第1穗果坐果时开始补光处理,到最后一次破坏性测量为止。使用条形LED光源(上海菁非光电),单支光源长度为1 m,出光面在光源两侧,距光源20 cm处的光合有效辐射为70 μmol/(m2·s),补光灯置于基质袋正上方,即两垄植株中间,悬挂高度随番茄生长进行调整。补光处理设置不同的红光(R)和蓝光(B)比例,R/B分别为4∶1,7∶1和9∶1,以未补光植株为对照,每个处理3垄。为平衡日光温室东西向不同位置对番茄生长的影响,3种补光处理间隔布置,补光灯工作时间为06:00-17:00。
1.3 测定指标及方法
1.3.1 产 量 以每穗果的4个果实转色为成熟节点,分别于2016年5月13日、5月20日、6月1日和6月13日分4次采摘成熟果实。每处理随机选取10株取样,统计4次成熟果实鲜质量之和,即为试验阶段的单株产量,并根据种植密度计算单位面积产量。
1.3.2 干物质量 分别于2016年4月18日和5月18日去除冠层基部老叶,并记录各处理老叶干质量,计入植株叶片总干质量。在每次采收成熟果实时,从各处理随机选取6穗果实(每穗4个)分别测定鲜质量和干质量,计算果实干物质含量,以此测算各处理中成熟果实总干质量,并与试验期结束前破坏性测量所得干质量累积,计算整个试验期干质量。
在补光结束之前(2016年6月15日)开展破坏性测量,各处理选取4株植物,将植株茎、叶、果各器官分开,测定鲜质量,然后用电热鼓风干燥箱(DHG-9620-A)于105 ℃烘干至恒质量,记录各器官干质量。叶片总干质量为破坏性测量叶片干质量与2次去除的基部老叶干质量之和。果实总干质量为破坏性测量果实干质量与4次采摘果实的干质量之和。茎、叶、果的干、鲜质量之和为试验期地上部分总干鲜质量,并计算各器官干物质分配比例和干物质含量。
各器官干物质分配比例=(各器官干质量/总干质量)×100%;
干物质含量=(各器官干质量/各器官鲜质量)×100%。
1.3.3 光合作用响应曲线 在番茄植株开花结果期测定番茄叶片光合作用的光响应曲线。光响应曲线测量采用配备荧光叶室的光合仪(LI-6400XT),于晴天09:00-15:00选取株间LED补光位置的叶片进行测定,叶室光源选择90%红光+10%蓝光,叶温设定为25 ℃,CO2浓度设定为400 μmol/mol,VPD保持在0.5~1 kPa,在叶室光源400 μmol/(m2·s) 下适应20 min后开始测量,叶室光照强度(PAR)设定依次为400,200,100,50,0,600,900,1 200,1 600 μmol/(m2·s)。各处理选取4株植物作为重复。测定值通过非直角双曲线模型进行拟合,得到光合作用光响应曲线及参数(表观量子效率(a)、最大净光合速率(Pmax)、暗呼吸速率(Rd)、凸度(θ))。非直角双曲线公式[16]如下:
式中:Pn为净光合速率,μmol/(m2·s);a为表观量子效率;I为光合有效辐射,μmol/(m2·s);Pmax为最大净光合速率,μmol/(m2·s);Rd为暗呼吸速率,μmol/(m2·s);θ为光响应曲线凸度。
1.3.4 叶片光学特性 各处理随机选取株间LED补光位置叶片6片,立即用浸湿的棉布包裹以防失水,带回实验室进行测量。采用光纤光谱仪(Ocean Optics USB 2000+)结合积分球(FOIS-1,ISP-REF,Dunedin)分别测量叶片在可见光波段的反射率和透过率,计算叶片可见光吸收率。
1.3.5 比叶面积 在补光处理结束后,选取补光位置叶片(两垄番茄内侧,补光灯所在位置的叶片)、对照处理中内侧叶片及所对应的外侧叶片,用直径1.6 cm的打孔器在选取叶片上避开主叶脉并随机打孔,每次取20个圆片样本,每个处理重复4次,电热鼓风干燥箱烘干至恒质量,测定其干质量,计算比叶面积。
比叶面积=叶面积/干质量。
1.4 数据统计与分析
试验数据统计与分析采用Excel和SPSS软件,处理间的多重比较采用Duncan法。
2 结果与分析
2.1 LED株间补光对番茄产量的影响
由图1可以看出,与对照相比,LED株间补光处理显著提高番茄产量,R/B=4∶1、R/B=7∶1、R/B=9∶1株间补光处理分别增产25.9%,28.5%和22.7%,其中R/B=7∶1补光处理增产效果最为明显,较对照提高了3.28 kg/m2。
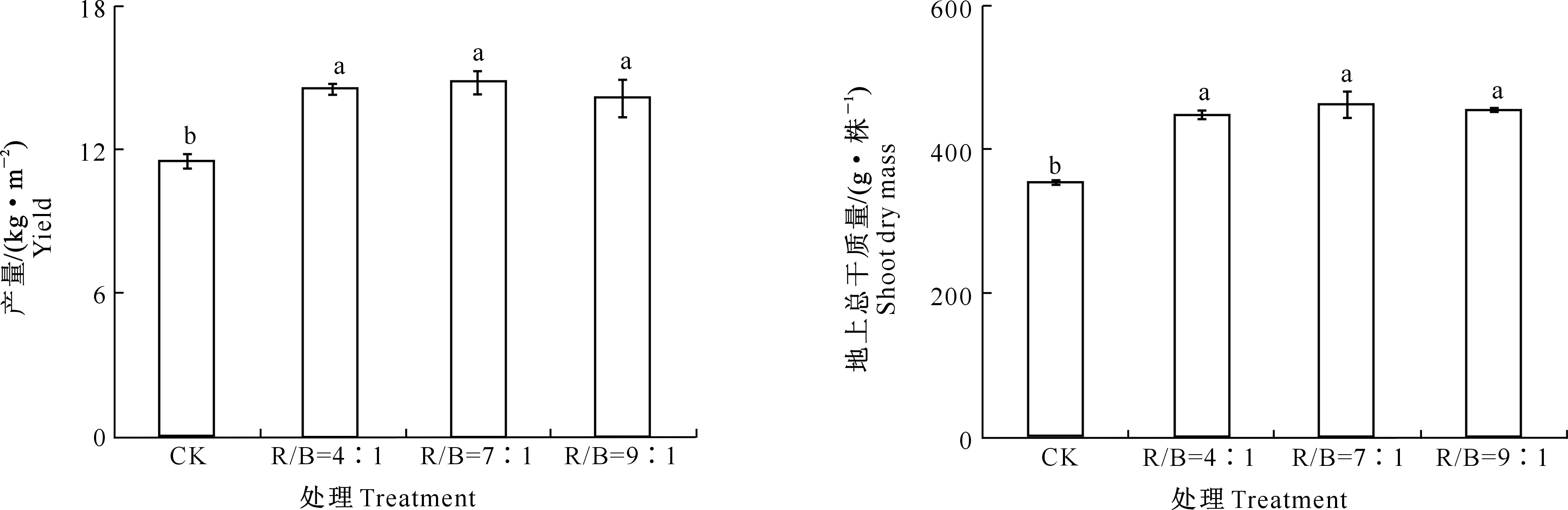
图柱上不同小写字母表示不同处理间差异显著(P<0.05)。下同Different lowercase letters represent significant difference among treatments (P<0.05).The same below图1 不同株间补光处理对日光温室番茄产量的影响Fig.1 Effects of interlighting on tomato yield in greenhouse
2.2 LED株间补光对番茄干物质分配及含量的影响
由图2可以看出,与对照相比,LED株间补光显著增加了番茄地上总干质量,R/B=4∶1、R/B=7∶1和R/B=9∶1处理番茄地上总干质量较对照分别显著提高了26.2%,30.4%和28.4%。
由表1可以看出,株间补光处理番茄果实干质量较对照显著提高,3种补光处理下果实干质量较对照分别提高31%(R/B=4∶1),38.6%(R/B=7∶1)和36%(R/B=9∶1),其中R/B=7∶1显著高于R/B=4∶1处理。株间补光提高了果实干物质分配比例,但差异不显著。
由表2可以看出,LED株间补光显著提高了果实干物质含量,其中R/B=9∶1补光处理最高。

表1 不同株间补光处理对日光温室番茄干物质分配的影响Table 1 Effects of interlighting on dry matter partitioning of tomato in greenhouse
注:同列数据后不同小写字母表示不同处理间差异显著(P<0.05)。下同。
Note:Different lowercase letters represent significant difference among treatments (P<0.05).The same below.

表2 不同株间补光处理对日光温室番茄各器官干物质含量的影响Table 2 Effects of interlighting on dry matter content (DMC) of tomato organs in greenhouse %
2.3 LED株间补光对番茄叶片光合能力及光学特性的影响
不同株间补光处理下番茄冠层叶片光合作用的光响应曲线及参数见图3和表3。由表3可以看出,与对照相比,株间补光显著提升番茄补光位置叶片最大净光合速率,但不同株间补光处理间的最大净光合速率差异不显著。株间补光对表观量子效率a、暗呼吸速率Rd和光响应曲线凸度θ均无显著影响。
由图4可以看出,株间补光处理提高了补光位置叶片对波长在500~650 nm可见光吸收率;株间补光对叶片在550 nm波段的绿光吸收率影响最大,3种红蓝光比R/B=4∶1、R/B=7∶1、R/B=9∶1株间补光较对照分别提高了4.0%,4.4%和5.8%。对于植物光合作用利用较多的红橙光(600~650 nm)平均吸收率分别较对照提高了1.5%,2.2%和2.3%。

图3 不同株间补光处理对番茄冠层叶片光合作用光响应曲线的影响Fig.3 Effects of interlighting on photosynthetic light response curves of tomato leaves

处理Treatment表观量子效率a暗呼吸速率/(μmol·m-2·s-1)Rd最大净光合速率/(μmol·m-2·s-1)Pmax凸度θCK0.078±0.006 a1.81±0.30 a 17.6±0.6 b0.66±0.14 aR/B=4∶10.078±0.003 a1.96±0.21 a22.7±1.4 a0.64±0.08 aR/B=7∶10.073±0.002 a1.75±0.14 a20.9±0.6 a0.76±0.06 aR/B=9∶10.064±0.008 a1.41±0.14 a21.7±0.9 a0.73±0.06 a
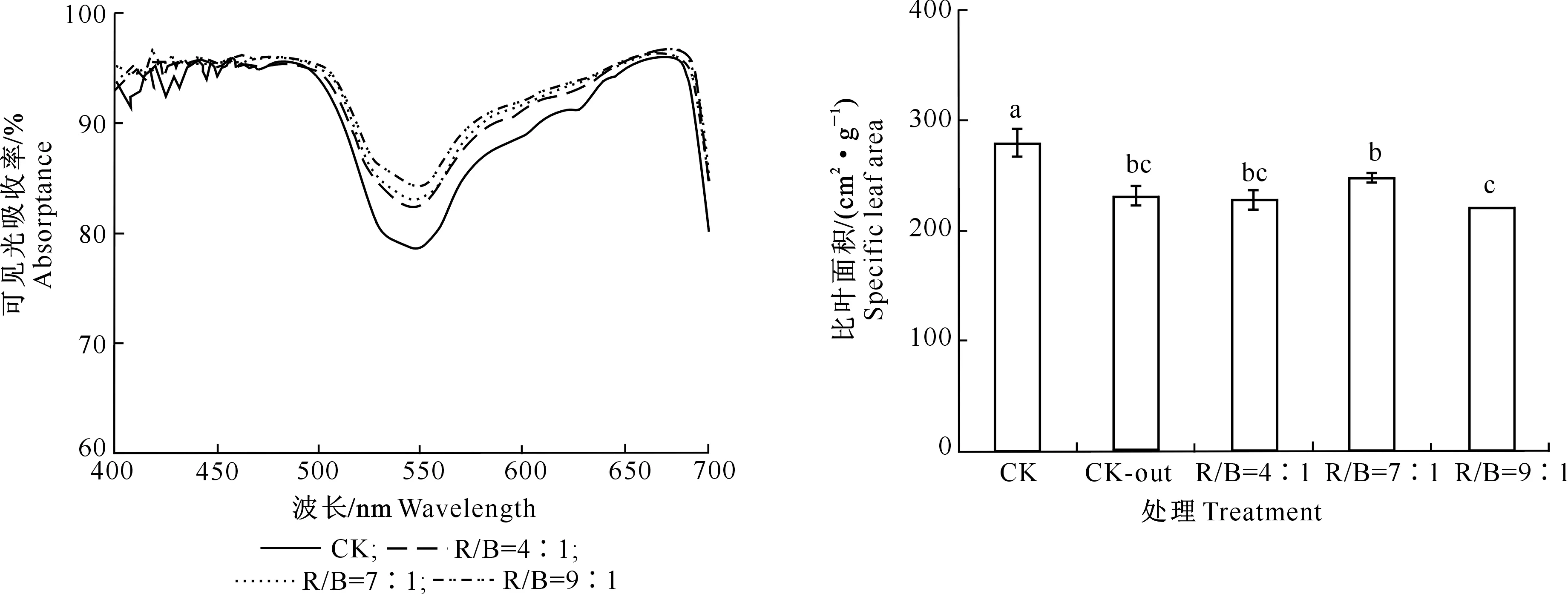
图4 不同株间补光处理对番茄叶片可见光吸收率的影响Fig.4 Effects of interlighting on tomato leaf absorptance in the visible light spectrum
2.4 LED株间补光对番茄叶片形态特征的影响
由图5可以看出,与对照相比,LED株间补光处理显著降低了番茄叶片的比叶面积,3种红蓝光比R/B=4∶1、R/B=7∶1、R/B=9∶1株间补光处理叶片比叶面积较对照分别降低18.5%,11.2%和21%,其中R/B=9∶1处理比叶面积最小。对照处理中两垄之间叶片(CK)比叶面积显著高于垄外侧对应位置的叶片(CK-out),对照处理两垄外侧叶片比叶面积与补光处理位置叶片比叶面积无显著差异。
3 讨 论
根据朗伯比尔定律,随叶面积指数增加,冠层光照强度呈指数关系下降。在温室番茄生产中随着冠层深度的增加,由上而下累积叶面积指数为1时的光照强度只有冠层顶部的40%;累积叶面积指数为2时的光照强度只有冠层顶部的20%[17]。大量研究证实,成熟番茄植株长期处于源库不均衡状态,即光合产物的合成不能满足植株各器官潜在生长需求[18],减少番茄果穗留果数能够增大剩余果实的大小也证实了这一点[19]。因此,株间弱光环境限制了番茄植株群体光合作用与产量的形成,也是番茄生殖生长时期处于源库不均衡状态的原因之一。人工补光作为调节番茄源库关系的有效手段,能够促进作物的光合作用,提高产量。因此,使用LED株间补光,研究其对番茄生长特性及产量的影响具有非常重要的意义。
本研究结果表明,LED株间补光处理显著提高了番茄产量,这主要是由于株间补光处理提高了番茄冠层中下部位的光照强度。温室作物补充光照强度能够增加叶片净光合速率、促进植株生长、增加果实产量、提高果实品质[20]。本研究中株间补光处理下,番茄产量、地上部分总干质量、果实干物质含量、最大净光合速率均显著提高。Hao等[21]的研究结果显示,补光处理能够增加黄瓜果实干物质分配比例,减少茎的干物质分配比例;Trouwborst等[9]的研究结果显示,株间补光能够增加黄瓜叶片干物质分配比例,而减少果实干物质分配比例。产生相反的结果可能是由于补光光源的使用造成处理间温度差异,以及不同的补光方式造成的。而本试验结果显示,株间补光对番茄各器官干物质分配无显著影响,仅有增加果实干物质分配比例的趋势。这可以从株间补光处理下各器官干质量变化中得到证实(株间补光处理下果实干质量较对照显著提高,而茎干质量和叶干质量没有显著变化)。本试验结果与Hao等[14]研究结果产生差异的原因在于,补光处理的时期及时长不同,并且补光光源对植物生长的影响效果也取决于补光时期自然光照的水平,这也是本试验中株间补光对各器官干物质分配无显著影响的一个原因。株间补光不仅能够提高番茄的产量,而且能够显著提高番茄果实干物质含量,这与Hao等[20]对黄瓜补光试验结果一致,更高的果实干物质含量有利于提升果实的品质,延长货架期,改善果实口感。
在光强较弱水平下,表观量子效率a和暗呼吸速率Rd是影响作物光合作用及产量的主要因素,而与最大净光合速率Pmax无关[9]。本试验结果表明,株间补光并未影响到植物叶片的表观量子效率和暗呼吸速率,只对最大净光合速率有显著影响,从株间补光处理下番茄冠层叶片光合作用光响应曲线可以看出,只有光合有效辐射高于400 μmol/(m2·s) 时,株间补光与对照处理的叶片净光合速率有差异。在温室环境条件下,番茄植株中下层叶片由于受到上层叶片的遮挡,叶片接受到的自然光强较弱,同时株间补光LED的光强相对较低,因此番茄中下层叶片仍处于相对弱光环境,在光响应曲线中处于光限制区域。随着室外光照强度的变化,当株间补光光源使中下层叶片受到的光强在光照周期内始终高于光补偿点时,将促进中下层叶片的光合作用,进而促进果实产量的增加。因此,株间补光对番茄产量及干物质的提高与叶片最大光合能力的改变并无关系,番茄植株产量及干物质的增加是由于株间光强的增加引起的。此外,植物叶片比叶面积也与光环境息息相关,比叶面积是叶片面积与叶片干质量的比值,叶片越厚比叶面积越低,反之则越高。叶片比叶面积的大小受光照强度、红光/远红光、水分、紫外光等环境因素的影响,其中光照强度对比叶面积的影响最大[21],在高光强下叶片比叶面积下降是叶片适应环境变化的结果[22]。本研究进一步证实了前人的结论,株间补光处理下叶片比叶面积显著降低,这是由于光强升高的结果;另外,对照处理垄内侧叶片(CK)比叶面积显著高于垄外侧叶片(CK-out),而株间补光处理下叶片比叶面积与CK-out没有显著差异,说明补光处理垄内侧与对照处理垄外侧叶片接受到的光照强度相似。
叶片光吸收率在蓝紫光区域和红光区域都达到了90%以上,而在绿光区域的吸收率降低到70%~80%,在绿光区域叶片反射和透射率更高[23]。本研究中,株间补光增加了叶片对波长在500~650 nm绿光及部分红橙光的吸收率。有研究显示,叶片对光的吸收率与叶片叶绿素含量呈正相关,其相关性在绿光区域更显著[24]。这可能是株间补光处理下叶片光吸收率提高的原因。植物叶片光量子效率与光吸收率有关[23]。本试验中叶片光吸收率的提高并未改变光量子效率,主要是由于绿光区域的光量子效率很低,而轻微提高绿光波段的光吸收率对可见光波段的光量子效率影响甚微。
不同波段光谱或光质比例补光对植株干物质分配及光合作用的影响不同,红光LED补光的黄瓜叶片净光合速率高于蓝光LED补光处理[25],补充红光能够促进光合产物向其它器官的运输能力,而补充蓝光增加光合器官自身的保留能力[11]。因而不同红蓝光比例LED株间补光对番茄叶片光合作用及干物质分配会产生不同程度的影响。而本试验中不同光质株间补光处理间叶片光合能力及各部位干物质分配比例差异并不显著,这是因为对于植株整体接受到的光照环境而言,无论是光质还是光强,都是以自然光为主,不同比例红蓝LED光源对株间光质环境影响有限,因此本研究不同光质株间补光处理仅有改变番茄植株各部位干物质分配比例的趋势,处理间差异并不显著。
4 结 论
株间补光改善温室番茄冠层光环境,提高冠层及植株整体光合能力,番茄产量增加22.7%~28.5%,果实干物质提高31%~38.6%。株间补光位置叶片在500~650 nm波段光吸收率和最大净光合速率Pmax均高于未补光植株;株间补光降低了补光位置叶片的比叶面积。不同红蓝光比例株间补光对番茄总干物质的积累和产量的形成均有显著的促进作用,但不同红蓝光比例处理差异不显著。
