灰胸薮鹛活动区的无线电遥测研究
2018-07-25孔赤平吴永恒向明付义强
孔赤平, 吴永恒, 向明, 付义强
(1.新疆农业大学动物科学学院,乌鲁木齐830052; 2. 乐山师范学院生命科学学院,西南山地濒危鸟类保护四川省高校重点实验室,四川乐山614004; 3. 广西大学动物科学技术学院,南宁530005)
活动区是栖息地中动物个体或种群取食、婚配、育雏等活动的区域(Burt,1943)。动物对活动区的利用,是其空间利用行为的一种体现,能够揭示该种群的空间结构模式,是了解种群与环境关系的重要基础(Uhía & Briones,2002;Febreretal.,2006)。开展动物的活动区研究,对于了解濒危珍稀物种的行为生态及栖息地,并依此制定科学的保护与管理对策具有重要意义。
灰胸薮鹛Liocichlaomeiensis隶属于雀形目Passeriformes噪鹛科Leiothrichidae薮鹛属Liocichla,是我国特有的珍稀鸟类之一(郑光美,2017)。该鸟种群数量稀少,分布区域狭小,仅偶见于四川省中南部及云南省东北部有限的山区森林中,被世界自然保护联盟(IUCN)列为易危(VU)物种(约翰·马敬能等,2000;雷富民,卢汰春,2006;IUCN,2017)。迄今,关于灰胸薮鹛的研究主要集中在繁殖生态、栖息地选择及分子生物学等方面(Fuetal.,2011,2016;Yangetal.,2017),未见其活动区的研究报道。为深化对该物种的认识,2016年1—2月和6—8月,利用无线电遥测技术在四川省峨眉山和四川老君山国家级自然保护区,对灰胸薮鹛的活动区进行了初步研究,旨在探明灰胸薮鹛非繁殖季和繁殖季活动区大小、重叠情况及个体的日活动节律。
1 研究区域
四川老君山国家级自然保护区(103°57′~104°04′E,28°39′~28°43′N)位于四川省宜宾市屏山县境内,总面积约35 km2,海拔900~2 009 m。保护区地处全球生物多样性保护热点的凉山山系。属亚热带湿润性季风气候,全年雨量充沛,年均降水量>1 500 mm,年均气温12~14.7 ℃,平均相对湿度>85%(Fuetal.,2011)。生境类型包括原生林、次生林和人工林。优势树种包括丝栗栲Castanopsisfargesii、木荷Schimasuperba、山茶Camelliajaponica、柃木Euryajaponica、糖槭Acersaccharum及杜鹃Rhododendronsimsii。在繁殖季,与灰胸薮鹛同域分布的常见鸟种包括棕噪鹛Garrulaxberthemyi、红翅噪鹛Trochalopteronformosus、红嘴相思鸟Leiothrixlutea、强脚树莺Horornisfortipes、金色鸦雀Suthoraverreauxi等(杨杰,2007)。
峨眉山风景区(103°15′~103°28′E,29°31′~29°38′N)位于四川省峨眉山市境内,面积154 km2,最高峰海拔3 099 m,山脚海拔500~600 m。风景区处于四川盆地边缘,属亚热带季风气候,年均降水量1 500~2 000 mm,相对湿度在80%以上,年均气温17.3 ℃。植被呈明显垂直分布:1 500 m以下为常绿阔叶林带;1 500~2 000 m为常绿落叶阔叶混交林带;2 000~2 500 m以上为针阔混交林带;2 500 m以上为寒温性针叶带。优势树种有白辛树Pterostyraxpsilophyllus、木瓜红Rehderodendronmacrocarpum、鸡爪枫Acerpalmatum、稠李Prunuspadus、润楠Machiluspingii(谷海燕,李策宏,2006)。在非繁殖季,与灰胸薮鹛同域分布的常见鸟种包括红嘴相思鸟、白颊噪鹛Garrulaxsannio、画眉Garrulaxcanorus、白头鹎Pycnonotussinensis、紫啸鸫Myophonuscaeruleus、灰喉鸦雀Sinosuthoraalphonsianus、棕脸鹟莺Abroscopusalbogularis、蓝鹀Emberizasiemsseni等。
2 研究方法
2.1 野外捕捉与标记
在灰胸薮鹛活动频繁的区域布设粘网,利用录音回放诱捕。每小时查看1次,防止鸟在网上悬挂时间过长而受伤。捕捉到灰胸薮鹛野生个体后,快速测量体质量等形态学指标,并为其戴上彩色塑料脚环(方便辨认),采用背带法佩带发射频率为150~151 MHz、电池设计寿命3个月的无线电发射器(Biotrack Ltd,UK)。捕捉到的灰胸薮鹛的平均体质量是32.6 g±1.2 g(n=6,30.7~34.0 g),发射器质量约1 g,低于灰胸薮鹛体质量的3%,对其活动不会造成影响。佩带发射器在15 min内完成,然后原地释放捕捉个体。
2.2 无线电遥测
无线电遥测借助三元Yagi天线和TRX-1000s无线电接收器(Wildlife Materials Inc,USA),利用三角定位法对标记个体遥测定位(Kenward,2000)。遥测在日出后和日落前进行。每只个体每日至少遥测8次,时间间隔约1 h。此外,还对部分个体进行了夜间遥测,以确定其夜栖地点。为减少由于动物移动所引起的误差,在最短时间内(小于5 min)完成每组位点的定位,方位角的夹角保持45°~135°(Kenward,2000)。
2.3 数据分析
2.3.1活动区面积及日活动性在ArcGIS 10.2中,首先将遥测位点的经纬度坐标全部转换成直角坐标,输出坐标系为“WGS 1984 UTM Zone 48N”。将转换好的直角坐标导入R 3.23中计算活动区面积,利用adehabitatHR包中的MCP函数和Kernel函数,即100%最小凸多边形法(minimum convex polygon,MCP)和95%固定核空间法(fixed kernel,FK),并用60%FK法所求得的活动面积作为每只个体的核心活动区面积(邱阳等,2009)。在使用固定核空间法计算活动区和核心区的面积时,均使用最小二乘交互检验(least squares cross validation)作为平滑因子(smoothing factor)(Gitzen & Millspaugh,2003;Gitzenetal.,2006)。计算活动区面积一般要求个体的合格位点数超过30组(Litvaitisetal.,1987;Seamanetal.,1999;Laver & Kelly,2008)。此外,测量相邻2个遥测位点间的平均距离,并绘制遥测个体的日活动距离变化曲线(刘阳,2006)。
繁殖季捕捉的2只灰胸薮鹛(B1♂和B2♀)为1对配偶,在遥测过程中找到了它们的巢,根据灰胸薮鹛产卵和孵卵的节律可以推算出它们的孵卵期和育雏期。分别计算了B1♂孵卵期和育雏期的活动区面积;由于B2♀在遥测10 d(处于孵卵期)后,因发射器信号故障无法定位,只计算了其孵卵期活动区面积。
2.3.2活动区重叠参考Fieberg和Kochanny(2005)的方法,计算灰胸薮鹛活动区面积(95%FK法)的重叠情况。在R 3.23中利用adehabitatHR包中的kernel overlap函数,选择“method=HR”计算活动区重叠百分率,公式为:HR[i,j]=A[i,j]/A[i]。其中A[i,j]是2只个体活动区重叠面积,A[i]是动物i的家域面积(Calenge,2006)。
数据采用平均数±标准误(Mean±SE)表示。由于样本量小,故没有对原始数据做统计学分析。
3 研究结果
共遥测6只灰胸薮鹛。其中,非繁殖季4只(2♂2♀),繁殖季2只(1♂1♀)。遥测总天数非繁殖季31 d,繁殖季59 d,共获取6组有效位点数据。
“地平线欧洲”高度重视对具有市场前景的高风险、颠覆性科研创新、创业活动的支持,专设资金、机构来落实这一政策导向,改变“地平线2020”下科研成果难以走出实验室的局面、真正实现计划下科研成果的市场价值。与此同时,中国目前也在加大对“大众创业,万众创新”的支持。
3.1 活动区面积
灰胸薮鹛非繁殖季平均活动区面积为19 148 m2±3 904 m2(100%MCP)、9 659 m2±975 m2(95%FK),平均核心区面积为2 542 m2±263 m2(60%FK)(n=4)。其中,雄鸟平均活动区面积为21 610 m2±2 507 m2(100%MCP)、10 472 m2±144 m2(95%FK),平均核心区面积为2 745 m2±143 m2(60%FK)(n=2);雌鸟平均活动区面积为16 685±3 896 m2(100%MCP)、8 846 m2±431 m2(95%FK),平均核心区面积为2 338 m2±144 m2(60%FK)(n=2),非繁殖季每只个体的活动区和核心区活动面积如表1所示。

表1 灰胸薮鹛非繁殖季的活动区和核心区面积Table 1 Home range size and core area size of Liocichla omeiensis during non-breeding season
注:*使用“季节+序号+性别”进行个体标识, N表示非繁殖季, B表示繁殖季; 下同
Notes:*Use “season+sequence number+sex” for individual identification, N represents non-breeding season, B represents breeding season; the same below
雄鸟(B1♂)繁殖季活动区面积为28 075 m2(100%MCP)、17 285 m2(95%FK),核心区面积为2 544 m2(60%FK);雌鸟(B2♀)繁殖季活动区面积为13 930 m2(100%MCP)、8 541 m2(95%FK),核心区面积为2 236 m2(60%FK);B1♂和B2♀在繁殖季平均活动区面积为21 721 m2±8 986 m2(100%MCP)、13 040 m2±6 003 m2(95%FK),平均核心区面积为1 581 m2±1 362 m2(60%FK)(n=2)。雄鸟(B1♂)孵卵期活动区面积和核心区面积大于雌鸟(B2♀),并且发现B1♂进入育雏期后的活动区面积变小(表2)。
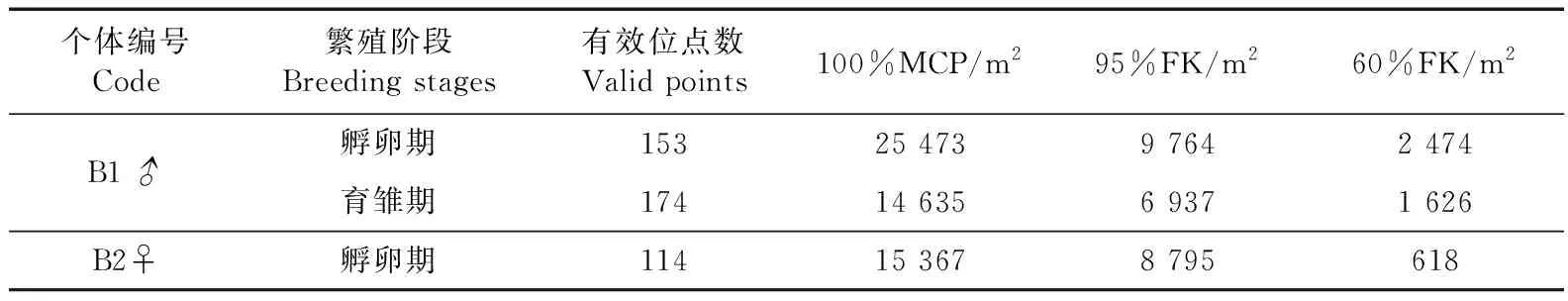
表2 灰胸薮鹛繁殖季的活动区和核心区面积Table 2 Home range size and core area size of Liocichla omeiensis in breeding season
3.2 活动区的重叠
非繁殖季遥测的4只个体,两两之间的活动区重叠百分率基本在50%以上。在遥测过程中,发现N1♂和N2♀可能为一对配偶,它们不仅白天的大部分时间在一起活动,晚上也在同一竹枝上夜栖,而这对灰胸薮鹛活动区重叠面积比例在70%以上(表3)。
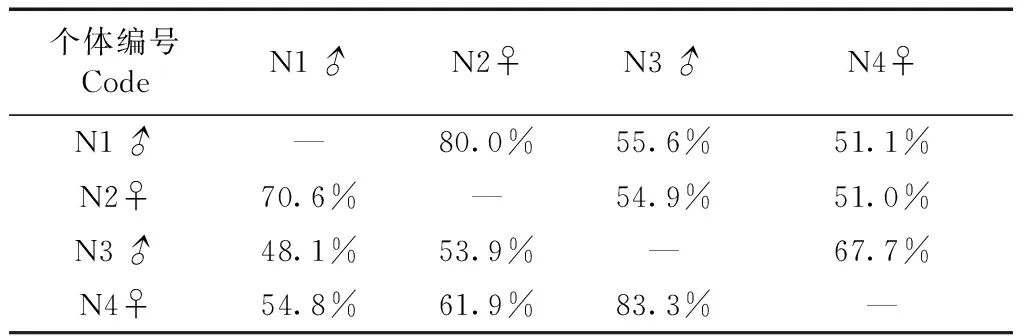
表3 非繁殖季遥测个体活动区的重叠指数(95%FK)Table 3 Overlap index of radio tracking individuals by means of 95% Fixed Kernel in non-breeding season
注: 对角线上、下方数值分别A[x, y]/A[y]、A[x, y]/A[x]; 下同
Note: The figures above and below the diagonal represent A[x, y]/A[y], A[x, y]/A[x], respectively; the same below
繁殖季捕捉到1对配偶(B1♂和B2♀),由于雌鸟只有孵卵期的遥测数据,因此,只计算孵卵期2只个体之间的活动区面积重叠指数。结果表明,二者重叠指数亦在70%以上(表4),与非繁殖季的N1♂和N2♀情况类似。
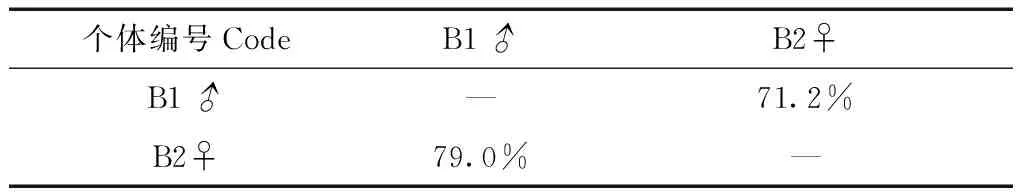
表4 繁殖季遥测个体活动区的重叠指数(95%FK)Table 4 Overlap index of radio tracking individuals by means of 95% Fixed Kernel in breeding season
3.3 个体的日活动性
对繁殖季的B1♂共进行了23个全天遥测,B2♀进行了4个全天遥测。结果发现,B1♂虽然有3个活动高峰期,但在各时间段内的位移距离波动不大。B2♀的日活动有2个高峰期,分别在10∶00—11∶00和15∶00—16∶00;对非繁殖季的N1♂和N2♀均进行了5个全天遥测。N1♂具有3个高峰期,分别出现在09∶00—10∶00、13∶00—14∶00和16∶00—17∶00。N2♀有2个高峰期,分别出现在09∶00—10∶00和17∶00—18∶00(图1)。
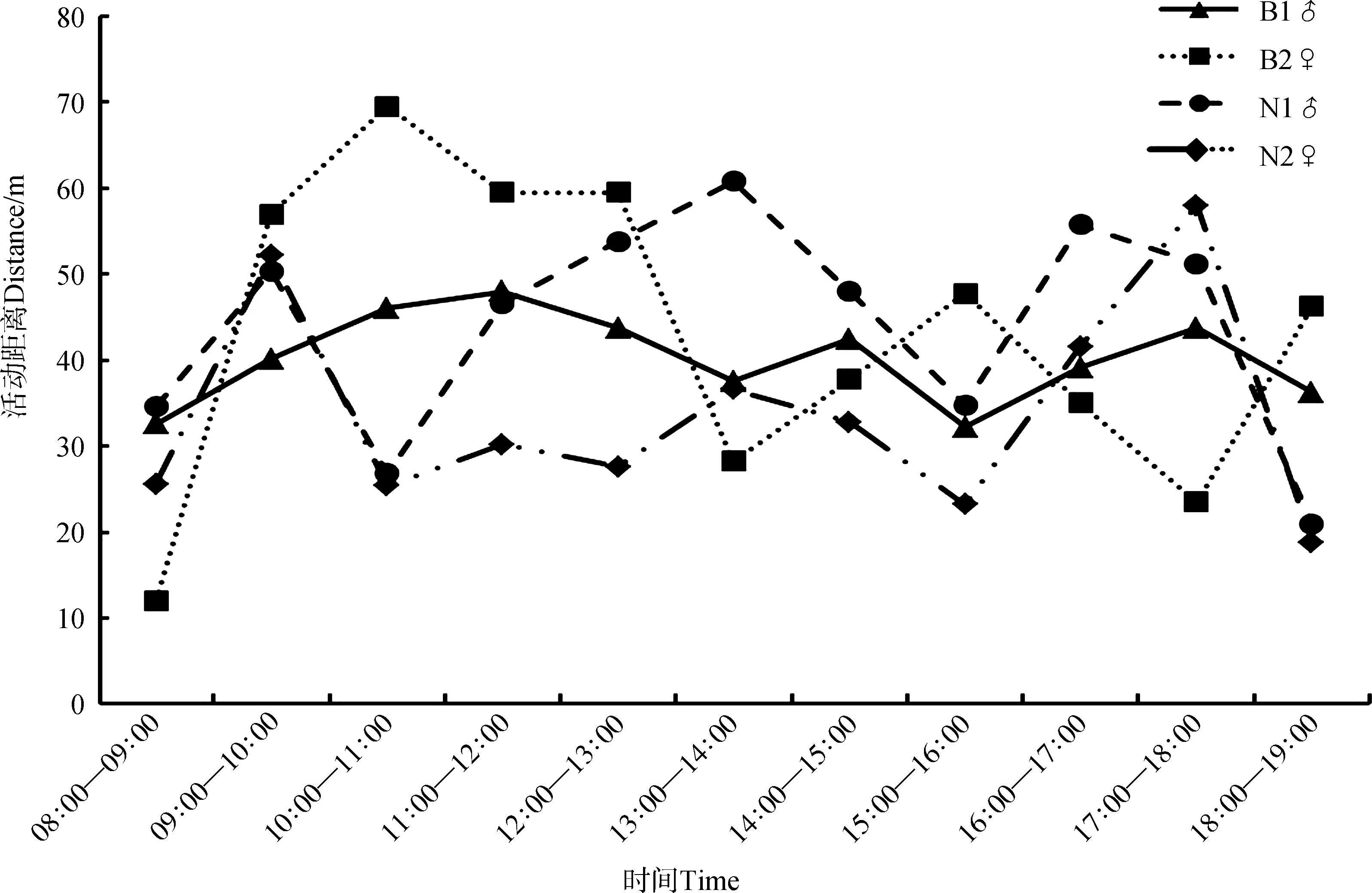
图1 不同灰胸薮鹛个体的日活动距离变化Fig. 1 Fluctuation of daily movement of different Liocichla omeiensis individuals
4 讨论
在非繁殖季,食物相对匮乏,鸟类需要扩大觅食范围以满足基本的能量需求。如,黄腹角雉Tragopancaboti夏季的活动区明显小于冬春季(孙悦华,郑光美,1992);白颈长尾雉Syrmaticusellioti冬季的月活动面积最大,繁殖期变小(蔡路昀等,2007)。本研究结果显示,灰胸薮鹛繁殖季的活动区面积大于非繁殖季,但核心区面积小于非繁殖季。在非繁殖季,野外工作点位于峨眉山伏虎寺周围,该寺庙的厨房有一条暗水沟通向庙外,每天有大量的食物残渣(如米粒、面条及蔬菜等)排出。另外,伏虎寺还有投喂野鸟的传统。非繁殖季遥测的4只灰胸薮鹛主要在此处活动。丰富且集中的食物可能是本研究中灰胸薮鹛非繁殖季活动区面积小于繁殖季的主要原因。例如,当停止对大山雀Parusmajor人工投食后,所有遥测个体的活动区面积均增大(van Overveld & Matthysen,2010)。而灰胸薮鹛繁殖季的核心活动区面积小于非繁殖季,则可能与其繁殖期领域防卫压力有关。在老君山二堰坪,丰富的灌丛与竹林是灰胸薮鹛、红嘴相思鸟、棕噪鹛及红翅噪鹛等鸟类理想的繁殖栖息地。推测繁殖季激烈的种内与种间竞争,可能会压缩生态位相近鸟类的领域空间。
本研究结果表明,灰胸薮鹛不同个体之间的活动区重叠程度较高。其中,非繁殖季灰胸薮鹛个体间的活动区重叠百分率基本在50%以上,该现象也见于褐马鸡Crossoptilonmantchuricum(王国栋,2006)。N1♂和N2♀活动区重叠百分率在70%以上,它们在一起夜栖,猜测可能为配偶关系。有研究证实,灰胸薮鹛的姐妹种——黄痣薮鹛Liocichlasteerii在非繁殖季亦维持配偶关系(罗柳墀,1987),灰胸薮鹛可能也具有该特性。在繁殖季遥测的1对灰胸薮鹛(B1♂和B2♀)确定为配偶关系,其活动区重叠百分率亦在70%以上。从野外观察来看,灰胸薮鹛配偶多成对活动,共同承担领域防卫及繁殖后代的任务,这是其活动区重叠百分率较高的主要原因。李景瑞(2005)也发现,在同一领域内出现的海南孔雀雉Polyplectronkatsumatae雌鸟和雄鸟活动区重叠面积很大,并且活动点经常交错在一起。
大量研究表明:鸟类的日活动具有显著的节律性(Rave & Baldassarre,1989;易国栋等,2010;钟雪等,2013)。许多野生鸟类晨昏各有1个觅食高峰期(柴璐艳等,2014;宋文宇等,2017)。在行为时间分配上,通常雌雄鸟有差异(徐纯柱等,2006;高志瑾等,2011;张延君等,2012)。本研究发现,灰胸薮鹛雌鸟在早、晚各有1个活动高峰期,可能主要与觅食活动有关;而雄鸟在早、中、晚各有1个活动高峰期,除了与觅食活动相关外,可能还与雄鸟担任主要的领域防卫任务有关。此外,还发现灰胸薮鹛繁殖期和非繁殖期的日活动节律大体相似,结合灰胸薮鹛配偶间的活动区重叠大,加之非繁殖季灰胸薮鹛具有典型的鸣唱歌声(Fuetal.,2013),表明该鸟在非繁殖季(如越冬期)可能存在领域行为,这在画眉科鸟类十分罕见。未来有必要进一步对灰胸薮鹛非繁殖季的日活动节律及其成因展开深入研究。
致谢:四川老君山国家级自然保护区为本研究提供了大力支持,陈文才参与了部分野外工作;戴波和娄尚灵老师分别对遥测及数据分析给予了很多指导与帮助;周博、思超、杨培培、雷红梅、曾兰义等人开展了大量前期工作,在此表示衷心的感谢!
