种群密度对大果虫实形态特征与异速生长的影响
2018-07-18范高华张金伟黄迎新神祥金禹朴家赵学勇
范高华,张金伟,黄迎新,神祥金,禹朴家,赵学勇
1 中国科学院东北地理与农业生态研究所, 长春 130102 2 中国科学院大学, 北京 100049 3 东北师范大学草地科学研究所 植被生态科学教育部重点实验室, 长春 130024 4 中国科学院西北生态环境资源研究院, 兰州 730000
植物不同组织、器官及其相互关系是植物生态学研究的重要内容,能反映出植物生长发育过程中株型、能量分配等重要信息。随种群密度变化,植物株型及生物量分配会呈现不同规律的生长变化[1-2]。株型涉及植株的根、茎、叶和繁殖等器官的特征及其在不同发育阶段的变化[3],以及各器官生物量的分配。作为生态系统生产力的重要指标,生物量是生态系统结构优劣和功能高低的直接体现,也是生态系统环境质量的综合体现[4]。生物量分配是植物生殖与生存平衡的结果,主要受植物遗传特性、环境条件和个体大小的影响[5],其分配模式在不同物种间差异较大。
当前,异速生长关系在刻画生物体器官结构与功能、器官生物量分配等特征上得到了大量应用[6- 10],其中以植物体生物学特征间的异速生长关系研究最为常见[11]。异速生长是指生物体的某些生物学特征(如个体形态指标、器官生物量等)与个体大小(重量)之间的幂函数关系。异速生长分析通过对数化大大减弱了单纯使用生物量学特征的绝对量或比例系数所带来的波动性,能揭示植物器官生物量分配特征间与尺度无关的内在规律[11]。已有研究表明,无论是一年生还是多年生草本植物,其地上、地下生物量间均存在异速生长关系,而且随密度增加,幂指数期望呈现近3/4的变化趋势。以往有关沙米、大果虫实等的异速生长的研究多侧重在水分及土壤营养等方面,而有关种群密度变化对植物形态特征与异速生长方面的影响研究较少,特别是繁殖分配以及繁殖器官与其他器官生物量间的异速生长关系[12]。此外沙漠化地区的植物研究相当匮乏[13- 17]。
大果虫实(Corispermummacrocarpum)为一年生草本,高20—50 cm[18-19]。大果虫实主要生长在流动、半流动和半固定沙丘上,是流动沙丘恢复过程中的主要物种之一,在沙漠化土地恢复的初期和中期比较常见。沙漠化恢复过程中,大果虫实种群密度往往发生剧烈变化,从每平米几株迅速增加到每平米上百株[20]。然而,在种群密度变化过程中,大果虫实器官生物量间是如何调整其生长及繁殖特性,是否具有异速生长规律,以及其异速生长规律是否发生变化均尚未明确。因此,本研究通过异速生长分析,分析种群密度对大果虫实形态特征、生物量分配和异速生长的影响,试图揭示大果虫实形态特征及器官生物量分配等功能性状变化的生态学机理,阐明其在退化土地恢复过程中的适应策略[21],以期为退化土地的恢复和荒漠化防治提供一定的理论基础。
其作用主要是强调了这场爱情的悲剧性,突出妻子的无结果付出的可悲情感,特意重复使用大段落,使得这种情绪更溢出听者心房。
1 研究地区与研究方法
1.1 研究区概况
研究区位于内蒙古通辽市奈曼旗中国科学院奈曼沙漠化研究站内,地处科尔沁沙地中南部,地理位置120°19′—121°35′E,42°14′—43°32′N,平均海拔360 m,具有多种不同的沙地类型和植被类型,土壤和植被梯度变化十分明显[22]。该区植被为温带半干旱草原植被,由于土地沙漠化严重,大部分地区半干旱草原植被为沙生植被所代替,植物种主要以沙米(Agriophyllumsquarrosum)、狗尾草(Setarriaviridis)、大果虫实(Corispermumelongatu)、糙隐子草(Cleistogenessquarrosa)、尖头叶藜(Chenopodiumacuminatum)和委陵菜(Potentillabifurca)等为主[21-22]。该区属温带半干旱大陆性季风气候,年均温6.3℃,≥10℃年积温在3161.2℃以上,无霜期约150 d。年均降水量365 mm,其中70%主要集中在6—9月,年蒸发量1935.4 mm。土壤类型为沙质栗钙土,沙土基质分布广泛,风沙活动强烈,年均风速3.5 m/s[21-22]。
不同植物根生物量分配随密度的变化有所不同[13, 15]。研究发现,大果虫实根生物量随密度的增大而显著减小,并与密度间呈极显著负相关(F=53.780,P<0.001)。根生物量D1-密度显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。随密度增大,大果虫实根生物量分配呈显著增大趋势,并与密度间呈极显著正相关(F=14.023,P<0.001)。根生物量分配D1—D4密度间差异显著(表4)。密度对大果虫实根冠比异速生长影响不显著(P=0.321)。密度对根:茎生物量间异速生长影响显著(P=0.028)。密度对根冠比、根:茎生物量间异速指数影响不显著(P=0.451;P=0.256),但对个体大小产生了极显著影响(P<0.001)(表1,图5)。随密度增大,根冠比、根/茎生物量呈增大趋势,二者与密度间呈极显著正相关(P<0.001)。根冠比D1—D2密度间差异不显著,其他密度间差异显著。根/茎生物量D2—D3、D2—D4密度间差异不显著,其他密度间差异显著(表1)。
选取太原市杏花岭区和小店区基层医院的在岗医务工作者为研究对象,于2016年8月1—31日区卫计委集中培训期间采用简单随机抽样法抽取180名进行调查研究。在获取研究对象知情同意后,使用自行设计的问卷进行面对面调查。本研究对象包括:妇女保健、儿童保健、预防保健或计划免疫科室(统称妇幼预防保健)及全科、临床各科、公共卫生、慢病及检验等其他科室就职的人员,不包括保洁、安保人员。在基层医务人员疫苗及安全接种知信行状况未知(P未知)的情况下,选取P=60%,允许误差 d=0.15P,α=0.05, 根据公式,计算得到样本量为119人。
研究发现,大果虫实繁殖器官生物量随密度增大而显著减小,繁殖器官生物量与密度间呈极显著负相关(F=43.584,P<0.001)。繁殖器官生物量D1-密度均显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。相应地,繁殖器官生物量分配也呈减小趋势,并与密度间呈极显著负相关(F=4.294,P<0.01)。D1—D2密度间差异不显著,其他密度间差异显著(表4)。研究还发现,除繁殖器官:根生物量间异速生长不随密度变化外(P>0.05) (表1,图2),密度对繁殖器官:地上部分、繁殖器官:茎及繁殖器官:叶生物量间的异速生长产生了显著影响(P<0.05),并对个体大小产生了极显著影响(P<0.001)。除密度对繁殖器官:叶生物量间的异速指数产生了极显著影响外(P<0.001),繁殖器官:地上部分、繁殖器官:根及繁殖器官:茎生物量间的异速指数不随密度变化而变化(P>0.05) (表1,图2)。此外,密度对繁殖器官/地上部分生物量影响不显著(P>0.05);对繁殖器官/茎生物量影响显著(P<0.05);对繁殖器官/根和繁殖器官/叶生物量的影响极显著(P<0.001),二者随密度增大而显著减小(表1,图2)。
1.2 试验设计
随外界环境的变化,植物各器官为更好地适应栖居地而发生了明显变化。茎不但为植株的生长输送必要的营养和水分,而且支撑叶片和花果,直接决定各器官的空间分布[3]。大果虫实茎生物量随密度的增大而显著减小,并与密度间呈极显著负相关(F=47.554,P<0.001)。茎生物量D1-密度显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。同样地,茎生物量分配随密度增大呈减小趋势,并与密度间呈显著负相关(F=3.148,P<0.05)。茎生物量分配D1—D4、D3—D4密度间差异显著,其他密度间差异不显著(表4)。密度对大果虫实茎:地上部分生物量间异速生长和个体大小的影响极显著(P<0.001)。密度对茎:地上部分生物量间异速指数影响不显著(P=0.102)(表1,图4)。茎/地上部分生物量不随密度变化而变化(P>0.05)。茎/地上部分生物量D1—D4、D3—D4密度间差异显著,其他密度间差异不显著(表1)。
显著性差异检验结果表明,株高与繁殖生物量异速系数在D1-密度与D2、D3、D4密度间无显著性差异(P>0.05),D2—D3、D3—D4密度间差异显著(P<0.01),D2—D4密度间差异极显著(P=0.005) (表3)。株高与根生物量异速系数在D3—D4密度间差异极显著(P=0.007),其他密度间差异不显著(表3)。株高与茎生物量异速系数在D1—D4密度间差异显著(P=0.047),D2—D4、D3—D4密度间差异极显著(P<0.01),其他密度间差异不显著(表3)。
1.3 数据分析
采用经典的异速生长方程:Y=βXa进行分析(式中:Y是某个需要研究的器官属性值,X是植物个体大小或另一个器官属性值,α是异速指数,即斜率;β是异速常数,即Y轴截距)。采用减小主轴回归(reduced major axis regression, RMA)拟合经对数转换后的数据,确定异速指数:αRMA=αOLS×r-1(αOLS 是最小二乘法得到的回归系数;r是相关系数),以去除变量偏差对回归系数的影响[21, 25]。用R软件的SMATR包,将数据log10转换后,进行标准主轴回归,分析密度对异速生长关系的影响,主要包括异速指数(斜率)、异速常数(截距)以及个体大小(X值变化),并比较异速指数与 3/4 的差异[26]。通过SPSS 17.0软件进行数据分析,采用单因素[种群密度(D)]方差分析比较各器官生物量特性以及生物量分配的可塑性。处理间差异显著性的检验采用F-检验,处理间平均数的比较采用最小显著性差异检验(LSD检验)。
1995年生效的《中华人民共和国国家赔偿法》(以下简称《国家赔偿法》),在很长时间内因鲜有申请人能成功获得国家赔偿而被谑称为“国家不赔法”。经过几次修改后,该法在赔偿程序和赔偿范围上有了很大进步,但关于司法赔偿的范围却一字未动。根据现行《国家赔偿法》的规定,司法赔偿仍仅限于刑事赔偿和法院因在民事、行政诉讼中违法采取强制措施、保全措施或者对判决、裁定及其他生效法律文书执行错误造成的损害,当事人有权利申请司法赔偿,将民事、行政错误裁判完全排除在司法赔偿范围之外。究其原因,主要有以下几点:
2 结果与分析
2.1 种群密度对大果虫实株高和器官生物量异速生长的影响
通过异速生长分析结果可以看出,种群密度对大果虫实株高:根生物量间异速生长具有显著影响(P<0.05),而对株高:地上部分、株高:繁殖器官及株高:茎生物量间异速生长影响不显著(P>0.05)(图 1,表1)。种群密度对大果虫实株高与器官生物量间异速指数和个体大小产生了极显著影响(P<0.01)(图 1,表1)。株高/根、株高/茎、株高/繁殖器官以及株高/地上部分生物量随密度增大而显著增大,四者在不同密度间差异极显著(P<0.001)(表1)。株高/根生物量D1—D4密度间差异显著。株高/茎、株高/繁殖器官生物量D4-密度均显著高于D1—D3密度,D1—D3密度间差异不显著。株高/地上部分生物量D1—D2、D2—D3密度间差异不显著,其他密度间差异显著(表1)。研究同时发现随种群密度增大,大果虫实株高呈减小趋势,株高与密度间差异极显著(F=70.602,P<0.001)。株高D1-密度均显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。

图1 大果虫实株高与器官生物量之间的异速生长关系Fig.1 The allometric relationship of Corispermum macrocarpum between height and organ biomass
于2008年8月29日进行破坏性取样,每个小区随机取样10株(为了避免取样的影响,每次取样前避免之前取样区域),每种处理共计40株。用清水将每株大果虫实的根冲洗干净,以保证获取较为完整的根系,并将每株植物分成根、茎、叶和繁殖器官(包括花和果)四部分,将植物放置于80℃烘箱内烘干称重,然后测得根、茎、叶、繁殖器官和地上部分的生物量,并计算株高/根、株高/茎、株高/繁殖器官、株高/地上部分、叶/地上部分、叶/根、叶/茎、根冠比、根/茎、茎/地上部分、繁殖器官/地上部分、繁殖器官/根、繁殖器官/茎及繁殖器官/叶生物量比,以及根、茎、叶、繁殖器官和地上部分的生物量分配,即各器官相对生物量[23-24]。
2.2 种群密度对大果虫实繁殖分配及其与其他器官生物量异速生长的影响
1、深化财政体制改革需要具体的财政事权清单。深化财政体制改革应在收入划分既定的基础上,列出财政事权的具体清单,将财政事权按照中央事权、地方事权和中央地方共担事权进一步划分,根据财政事权划分的属性原则,尽可能划分出具体清晰的事权清单。财政事权的清单,是与行政部门的行政事权清单有机统一,是“钱”与“事”的有机统一,所以每个领域的事权清单都要体现业务管理部门的主体责任,体现行业的特殊属性,与此相结合,形成钱事统一的事权清单。
2.3 种群密度对大果虫实叶片生物量分配及叶与其他器官生物量异速生长的影响
个体发育对植物器官生物量分配的影响因物种而异,反映了有机体在整个发育阶段各种功能的权衡,一般认为这是由物种内在的基因所决定的[30-31]。环境条件的变化会对植物分配产生重要影响[12]。本研究发现,大果虫实在不同竞争强度下会采取不同的生物量最优分配策略,即大果虫实器官生物量分配响应不同密度具有可塑性。不同器官生物量分配格局反映了大果虫实应对环境变化的策略。随密度增加,大果虫实各器官生物量呈减小趋势。各器官生物量D1-密度均显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。大果虫实各器官生物量变化量顺序为:繁殖器官>茎>根>叶生物量,而且,各器官生物量分配顺序也是:繁殖器官>茎>根>叶生物量分配,这说明大果虫实在权衡过程中,保持高的繁殖分配,以顺利完成生活史,也为后代在繁殖过程中更好地进化提供了条件;而且,随密度增加,繁殖器官的生长变化最大,茎其次,叶片最小,表明大果虫实地上器官对种群密度变化最敏感,处于高密度种群的植株可以获得更高的地上生物量分配。
表1种群密度对大果虫实各器官间生物量比特性的影响(平均值±标准误)
Table1TheinfluenceofCorispermummacrocarpumunderpopulationdensityonthebiomassratiocharacteristicsofthedifferentorgans(means ± SE)

性状Traits密度处理Densitytreatment/(株/m2)1644.4100400F值FvalueP1P2P3株高/根Height/root975.33±60.85a2918.10±254.82b4217.60±380.03c7562.80±785.66d49.609∗∗∗0.0330.003<0.00 株高/茎Height/stem180.29±16.53a846.96±92.32a1666.00±321.01a8798.80±3350.60b8.409∗∗∗0.0660.008<0.001 株高/繁殖Height/reproductiveparts114.57±16.49a505.74±84.18a903.34±97.69a2672.30±756.23b13.041∗∗∗0.8950.002<0.001 株高/地上Height/aboveground64.15±6.76a275.47±33.70ab490.93±57.48b1412.14±369.08c15.122∗∗∗0.6300.001<0.001 叶/地上Leaves/aboveground0.043±0.005a0.080±0.007b0.095±0.013b0.140±0.026c9.565∗∗∗0.0050.004<0.001 叶/根Leaves/root0.742±0.083a0.914±0.075ab0.919±0.186ab0.881±0.106b0.564ns0.0790.014<0.001 叶/茎Leaves/stem0.132±0.016a0.268±0.030a0.346±0.086a0.837±0.265a6.890∗∗∗<0.0010.05<0.001 根冠比Root/shootratio0.065±0.004a0.092±0.006a0.118±0.010b0.174±0.031c11.507∗∗∗0.3210.451<0.001 根/茎Root/stem0.184±0.011a0.298±0.025bc0.381±0.058b0.989±0.302c8.040∗∗∗0.0280.256<0.001 茎/地上Stem/aboveground0.357±0.016a0.328±0.018ab0.363±0.022a0.288±0.036b2.091ns<0.0010.102<0.001 繁殖/地上Reproductive/aboveground0.600±0.015a0.593±0.017ab0.542±0.017b0.573±0.032b1.868ns0.0380.125<0.001 繁殖/根Reproductive/root10.415±0.671a7.089±0.465a5.012±0.321a4.624±0.663a23.494∗∗∗0.0520.311<0.001 繁殖/茎Reproductive/stem1.789±0.094a1.974±0.134ab1.759±0.223b3.032±0.664b3.662∗0.0030.255<0.001 繁殖/叶Reproductive/Leaves19.079±2.208a8.669±0.755b7.974±1.037b5.781±0.711b16.897∗∗∗0.009<0.001<0.001
P1表示异速指数;P2表示异速常数;P3表示个体大小;P1 is the allometric exponent;P2 is the allometric constant;P3 is individual size

表2 种群密度对大果虫实株高及生物量特性的影响 (平均值±标准误)
不同小写字母表示种群密度间平均数的差异显著;处理间差异显著性的检验采用F-检验;nsP>0.05;**P< 0.01;***P< 0.001
2.4 种群密度对茎生物量分配及茎与地上部分生物量异速生长的影响
2008年5月初,在中国科学院奈曼沙漠化研究站内,选择质地均匀的沙质草地,通过人工播种,种植大果虫实,根据沙地上大果虫实自然种群随退化沙地恢复过程中的密度变化范围,对大果虫实进行密度处理,共设置4个水平,每个处理水平4个重复,共16个小区,小区面积(3 m×3 m):1) 16株/m2(D1);2) 44.4株/m2(D2);3) 100株/m2(D3);4) 400株/m2(D4)。在2叶期(大约2周左右)进行间苗,按照D1—D4密度处理定苗。整个生长阶段,除每3—4周进行一次人工剔除其他杂草外,无其他人为干扰。
表3不同密度下大果虫实株高与器官生物量各个斜率间显著性差异检验(平均值±标准误)
Table3UnderdifferentdensityonCorispermummacrocarpumeachslopeofsignificantdifferencebetweenplantheightandorgans′biomass(means±SE)

密度处理Densitytreatment株高/各器官生物量Height/organsbiomass44.4株/m2100株/m2400株/m2ABCABCABC16株/m2A0.690ns0.096ns0.062ns B0.995ns0.209ns0.082ns C0.930ns0.418ns0.047∗ 44.4株/m2A0.019∗0.005∗∗ B0.233ns0.086ns C0.230ns0.002∗∗ 100株/m2A0.045∗ B0.007∗∗ C0.005∗∗
A代表株高与繁殖生物量;B代表株高与根生物量;C代表株高与茎生物量;ns表示差异不显著;*表示P< 0.05;**表示P< 0.01;*** 表示P< 0.001
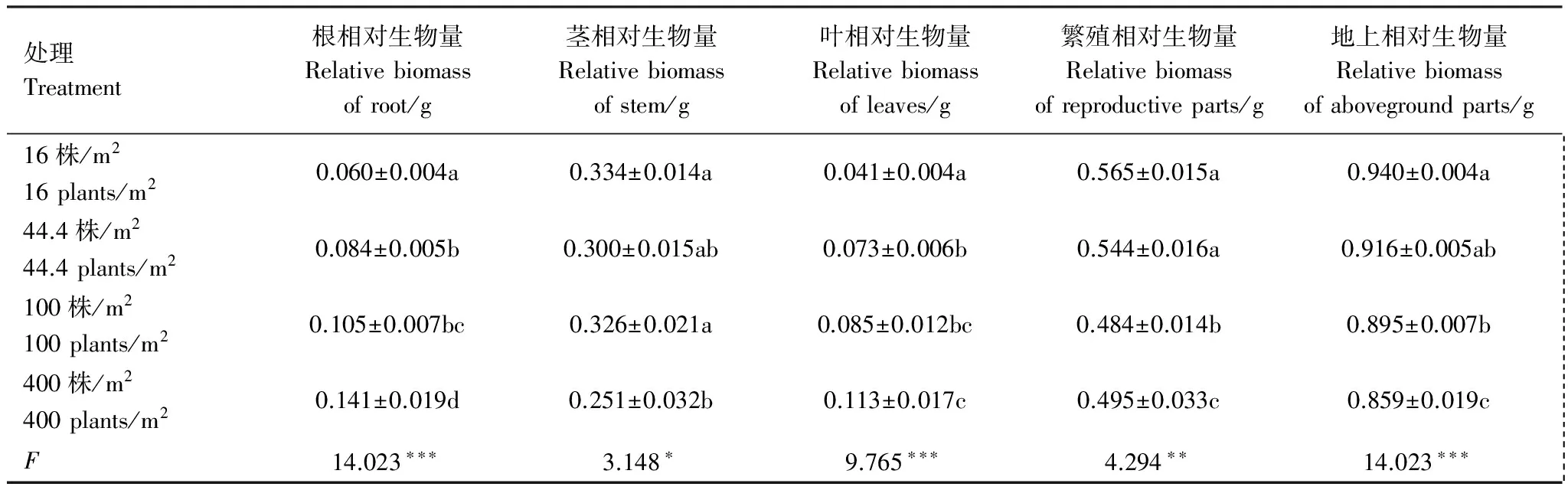
表4 种群密度对大果虫实生物量分配特性的影响 (平均值±标准误)
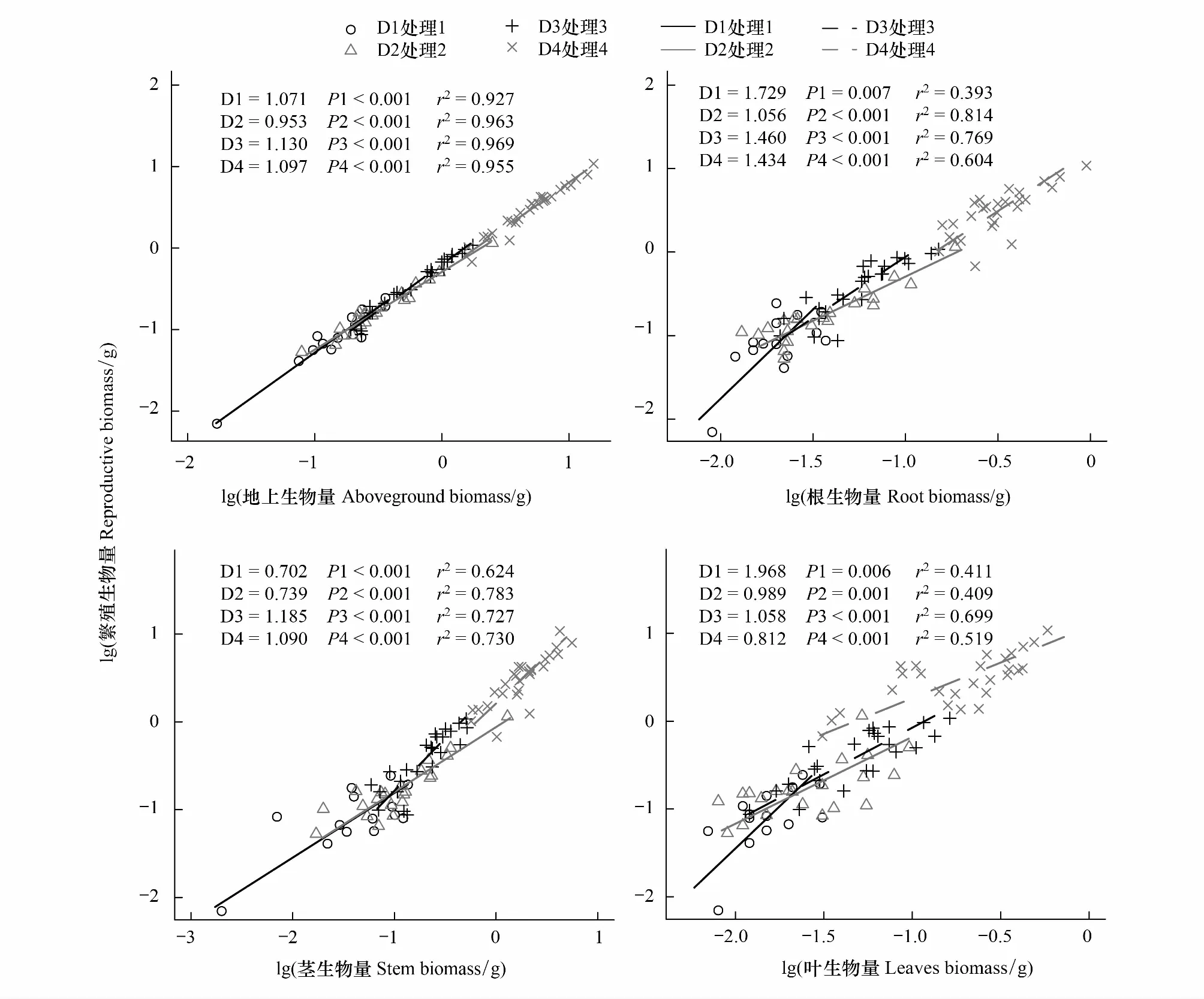
图2 大果虫实繁殖器官生物量与其他器官生物量之间的异速生长关系Fig.2 The allometric relationship of Corispermum macrocarpum between reproductive biomass and other organ biomassD1、D2、D3、D4分别代表16、44、100、400株/m2
2.5 种群密度对根生物量分配及根与其他器官生物量异速生长的影响
为了验证笔者设计的写作教学模式是否可以切实改善学生作文中句式、内容和语篇结构等方面的问题,提高写作课堂教学效率,笔者在学期初和学期末设计了前后测,于第一周和第十六周的写作课堂上进行。测试采用纸制考试的形式,学生需在30分钟内完成规定的写作任务,不得使用任何词典。为确保评分信度,特邀四位参加过全国大学英语四级考试阅卷工作的老师对作文分别进行评阅,从7个方面进行打分,包括篇章结构、内容切题、连贯性、词汇、语法、长句及写作规范,每项成绩都取四位老师评分的平均分。

图3 大果虫实叶片生物量与其他器官生物量之间的异速生长关系Fig.3 The allometric relationship of Corispermum macrocarpum between leaf biomass and other organ biomass

图4 大果虫实茎生物量与地上器官生物量之间的异速生长关系Fig.4 The allometric relationship of Corispermum macrocarpum between stem and aboveground biomass

图5 大果虫实根生物量与其他器官生物量之间的异速生长关系Fig.5 The allometric relationship of Corispermum macrocarpum between root and other organs biomass
3 讨论
3.1 大果虫实形态特征分析
高等植物通过调节顶端分生组织和侧生分生组织的活性建立地上株型系统,分生组织的活性受环境信号、发育阶段和遗传因素的综合调控,这些因素的综合调控使植物具备了发育可塑性和对环境的适应性[28]。以往研究表明,随密度增加,植物个体会将其高度维持在一最优值以使植物个体能最大化获取光资源,并且小个体相对于大个体能更早的将其高度维持在最优值[29]。本研究发现,随着种群密度增加,大果虫实植株呈不同的株型系统,其分枝数及分枝长度明显减小:D1-密度时,二、三级分枝较多,分枝数最高可达389,最低为45,总分枝长度最高可达4330 cm,最低为1328 cm;D4-密度时,大部分植株只有一级分枝,二、三级分枝几乎没有,分枝数最高只有18,总分枝长度最高只有536 cm。这表明密度变化对大果虫实的株型构建产生了显著地影响。这些变化很可能是受多种基因调控的,其主要是通过影响茎顶端分生组织的活动决定茎秆的形成与发育,进而调控大果虫实侧生器官的数目和形态[17]。
3.2 大果虫实器官生物量分配格局分析
叶生物量分配体现了植物在整个生长阶段应对外界环境变化的分配策略[27]。研究发现,随密度的增大,大果虫实叶片生物量呈减小趋势,并与密度间呈极显著负相关(F=36.370,P<0.001)。叶生物量D1-密度显著高于D2—D4密度,D2—D4密度间差异不显著(表2)。然而,叶生物量分配随密度的增大而增加,并与密度间呈极显著正相关(F=9.765,P<0.001)。叶生物量分配D1-密度显著低于D2—D4密度,D2—D4密度间差异显著(表4)。密度对大果虫实叶片:地上部分、叶:茎生物量间的异速生长产生了极显著影响(P<0.01)。然而,叶:根生物量与密度间异速生长关系差异不显著(P=0.079)(表1,图3)。密度对叶:地上部分生物量间的异速指数产生了极显著影响(P<0.001)。密度对叶:根及叶:茎生物量间的异速指数产生了显著影响(P<0.05),并对个体大小产生了极显著影响(P<0.001)(表1,图3)。叶/根生物量不随密度变化而变化(F=0.564,P>0.05),叶/地上、叶/茎生物量随密度的增大而增大,二者与密度间呈极显著正相关 (F=9.565,P<0.001;F=6.890,P<0.001)(表1)。
本研究分析发现,随密度增大,大果虫实根和叶生物量分配呈增大趋势,并且根、叶生物量分配随密度变化呈现一致的变化规律(表3),这与最优化分配理论中水、矿质营养和光资源受限时的情况一致;而茎、繁殖器官和地上部分生物量分配呈减小趋势,三者之间的生物量分配呈现异速生长规律(表3)。最优化理论预测表明:低光条件下,植物分配更多地资源给茎和叶的生长;低营养条件下,植物会将更多地资源优先分配给根的生长[32- 34]。植物对光、水分及矿质营养等资源竞争能力的高低是通过器官生物量分配权衡实现的[35]。这说明,对于基因型相同的植物而言,其各器官生物量分配响应不同密度具有可塑性;同时,也表明最优化分配理论与异速生长理论间并非没有关系可言,可能呈一种互补关系,二者综合分析能较好的解释大果虫实器官生物量间的权衡关系[36]。种群密度变化导致邻体植物间地上和地下部分对资源产生竞争,进而调节植物的资源配置。随密度增大,大果虫实植株增加地下部分生物量分配而减少地上部分生物量分配,表明大果虫实对水、矿质营养和物理空间等地下资源的竞争能力随密度增加而增加,而对地上光资源的竞争能力随密度的增加而降低。大果虫实根系生物量的增加促进其对水分和营养的吸收,这为大果虫实的正常生长提供了物质和能量保证。在器官水平上,根和叶生物量分配增加,茎和繁殖生物量分配减小,说明在高密度条件下,根生物量分配符合最优化理论中矿质营养受限的情况,而叶生物量分配的增加可能是叶的延长所致,这种表型变异是植物对遮阴的“躲避反应”,植物可以通过周围环境中红外:远红外光比率的变化感应相邻个体的存在[37-38]。当大果虫实密度过高、植株间相互遮蔽时,由于叶绿素对可见光的选择性吸收,会造成周围环境中红光:远红光比率(R∶FR)降低,并作为环境信号被由光敏色素基因编码的光受体接收,通过信号传递通路,进而调节一系列生理生化反应和发育程序[39- 41],即植物的“避荫”反应。因此,荒漠化生境中大果虫实在演替早期利用根和叶的优势充分获取地下地上部分的水分、矿质营养和光资源,将更多的资源投入到繁殖生长,顺利完成其生活史,维持荒漠化生境中的种子库,为荒漠化的改善提供了种源,为荒漠化生境的逆转奠定了基础。
3.3 大果虫实器官生物量间异速生长分析
研究发现,随密度增加,大果虫实株高与器官生物量间均呈现简单异速生长关系,并未出现复杂线性关系;但密度变化对株高与器官生物量间异速指数和个体大小均产生了极显著影响,说明密度的确改变了大果虫实株高的生长变化及其器官生物量的分配策略。株高随密度增加显著减小,而各器官生物量间则通过权衡生物量分配来完成生活史。密度对株高:根生物量间的异速生长具有显著影响,而对株高:地上部分、株高:繁殖器官及株高:茎生物量间异速生长影响不显著,说明密度变化引起了大果虫实种内对地下水和矿质营养的竞争,与最优化理论中地下营养受限的情况一致。
表型可塑性赋予生物个体在不同环境条件下通过产生不同的表型来维持其适合度的能力[42-43]。表型可塑性不仅是物种适应性进化的一个重要方面,也是应对环境变化而选择进化的产物,各种表型可塑性的存在对其生态适应和进化模式具有不可或缺的作用。以往的研究发现,不同植物响应密度处理的结果不尽一致,密度处理对反枝苋营养生长和繁殖生长间的异速关系无显著影响[44],密度同样对苍耳、荞麦和曼陀罗整个生长季中繁殖分配的异速生长无显著影响[45];但密度对刺沙蓬、沙米、角果藜、对节刺和尖头叶藜等1年生藜科植物的构件形态、生物量间的异速关系具有显著影响[23-24, 46]。本研究发现,大果虫实繁殖器官:根生物量间的异速生长不随密度变化而变化,而密度对繁殖器官/根生物量的影响极显著,这表明繁殖器官与根之间的变化是受个体大小制约的,而不是真正响应种群密度所发生的变化,属于表观可塑性[20-21,26]。密度对繁殖器官:地上部分生物量间的异速生长影响显著,却对繁殖器官/地上部分生物量影响不显著,属于真正可塑性。繁殖器官与其他器官生物量间的异速生长及其生物量比均受密度的极显著影响,表明密度变化的确影响了大果虫实繁殖器官的生长变化,随密度增大,大果虫实将更多地资源投入到繁殖生长,亦属于真正可塑性。对于大果虫实而言,真正可塑性是为更好的适应生长发育过程中密度变化所带来的影响,产生不同的表型确保大果虫实生活史的顺利完成[47]。繁殖器官:茎生物量间的异速指数在D1和D2-密度时与3/4差异不显著(P=0.925;P=0.886),符合生态代谢理论。繁殖器官:叶生物量间的异速指数在D2和D4-密度时与3/4差异不显著(P=0.119;P=0.570),亦符合生态代谢理论。
五是未建立有效的盈利模式。孵化器的收入与入驻企业的数量和购买能力紧密相关,而由于目标客户(入驻企业)数量及自身服务功能配置等因素的局限性,Z公司的主要收入来源为场地租赁收入及商务服务收入,存在收入来源单一并且不稳定的现象,不利于Z公司孵化器业务的可持续发展。
本研究中,大果虫实叶片:根生物量间的异速生长受密度的影响不显著,且密度对叶/根生物量无显著影响,属于表观可塑性[21, 31]。密度对叶与其他器官生物量间的异速生长及生物量比均有极显著影响,属于真正可塑性,表明密度变化对大果虫实叶片的生长变化产生了重要影响。叶∶根、叶∶地上部分生物量间的异速指数在D1和D2-密度时与3/4无显著性差异(P>0.05),符合生态代谢理论。叶:茎生物量间的异速指数在D2-密度时与3/4差异不显著(P=0.985),亦符合生态代谢理论叶与个体大小方面的预测。同时发现,根∶地上部分生物量间的异速生长不随密度变化而变化,而密度对根/地上生物量具有显著影响,属于表观可塑性。根∶茎生物量间的异速生长及其生物量比均受密度变化的显著影响,属于真正可塑性。密度对大果虫实茎∶地上部分生物量间的异速生长具有极显著影响,但茎/地上部分生物量不随密度变化而变化,亦属于真正可塑性。这表明密度变化在一定程度上影响了大果虫实根、茎的生长变化。
随物种的不同,物种对环境变化的敏感性、发生可塑性变异的能力,以及可塑性反应的模式不尽相同[43]。本研究中,大果虫实随密度增加,其株高和器官生物量分配都发生了明显的变化,其表型可塑性变异的发生与个体发育过程中由不同环境因素诱导的基因选择性表达密切相关[48]。通过基因的选择性表达,建立一种新的基因表达模式,进而改变大果虫实个体发育的进程和发育轨迹。这可能是大果虫实株高及器官生物量分配发生变化的主要原因。为应对特定的环境条件,不同种群的遗传结构或不同个体的基因型组成,在自然选择的调控下可能倾向于某些特殊的可塑性反应类型,结果不仅导致适应性表型可塑性变异的发生,同时也为可塑性的适应性进化提供了可能[47, 49-50],本研究中大果虫实即是在这种机制驱动下表现出不同的表型可塑性,也为大果虫实在极端环境条件下顺利完成生活史提供了保障。所以说,可塑性赋予了特定基因型的个体一定的适应性弹性,有助于提高种群的生存能力和选择进化的能力[51-52]。
4 结论
在所有种群密度下,大果虫实株高与器官生物量间呈异速生长关系;大果虫实的分枝数及分枝长度明显减小:从D1—D4密度,分枝数从最高的389减少到18,分枝长度从最长的4330 cm减小到536 cm。各器官生物量随密度增大呈减小趋势,具体器官变化量表现为:繁殖器官>茎>根>叶。虽然各器官生物量分配变化量也为:繁殖器官>茎>根>叶,但大果虫实根和叶生物量分配呈增大趋势,而茎、繁殖器官和地上部分生物量分配呈减小趋势,这与最优化分配理论中水、矿质营养和光资源受限时的情况一致。根∶地上部分、叶片∶根、繁殖器官∶根生物量间的异速生长属于表观可塑性。根∶茎、茎∶地上部分、叶与其他器官及繁殖器官与其他器官生物量间的异速生长属于真正可塑性。随密度的增加,大果虫实叶和繁殖器官生物量的变化是真正可塑性,说明大果虫实叶片分配及繁殖策略是由种群密度差异造成的,并且是非个体大小依赖的。这些变化导致大果虫实形态特征及器官生物量之间在生长过程中权衡资源分配,形成不同的株型系统以顺利完成生活史。
