白曲酸性蛋白酶在无孢黑曲SH-2中的克隆表达及酶学性质分析
2018-07-11董文超王斌潘力
董文超,王斌,潘力
(华南理工大学生物科学与工程学院,广东广州 510006)
白曲作为一种重要的麸曲,酸性蛋白酶对粮食原料颗粒的溶解作用,促进了糖化作用,提高了淀粉的利用率,在一定程度上提高了出酒率,在酸性条件下,将植物蛋白分解为各类氨基酸,为堆积过程中的美拉德反应提供前提物质[1]。并且白曲酸性蛋白酶影响着芝麻香酒中杂环化合物的数量和种类,对芝麻香酒的风味有极其重要的影响。提高白曲酸性蛋白酶含量,对提高芝麻香酒的生产效率及稳定和提高质量都有重要的意义[2]。因此构造一株能高效表达酸性蛋白酶,而不引入其他无关蛋白的菌株对白酒生产工艺的改进具有很好的应用价值。
目前国内生产的酸性蛋白酶基因来源主要是黑曲霉、宇佐美曲霉和米曲霉。费笛波[3]等对黑曲霉变异菌株6042进行液体深层发酵,经4批次2300 L发酵罐扩大,使得酸性蛋白酶酶活达到了2535 U/mg,方春玉等[4]采用固体发酵技术,以黑曲霉为菌种,全面优化发酵条件,使发酵产物酸性蛋白酶酶活达 24350 U/g,平均酶活提高64.6%,杨琥等[5]以毕赤酵母为宿主,表达宇佐美曲霉酸性蛋白酶基因(GenBankTM accession number XM_001401056),产物经纯化后比活性为4127.3 U/mg。胡稳奇等[6]从土壤中分离出一株米曲霉菌株,在实验室最适产酶条件下酶活达 6789.6 U/g。
运用诱变、基因重组及发酵优化手段来提高黑曲霉、宇佐美曲霉和米曲霉源的酸性蛋白酶的报道,比较常见,但白曲霉源的酸性蛋白酶相关的报道很少见。其中,李杰等[7]构建了黑曲霉表达载体pSZHG-pepB,通过农杆菌介导法转化黑曲霉 CICC2462,其发酵产物酸性蛋白酶酶活达5543 U/mL。杨猛等[8]白曲是以麸皮为原料,通过单因素试验和正交试验确定白曲的最佳配方,用此配方培养白曲36 h,酸性蛋白酶活力为l173 U/g。
本研究采用丝状真菌表达系统,以经基因工程改造的低内源蛋白背景的无孢黑曲霉 SH-2为宿主,表达白曲霉酸性蛋白酶基因pepB,并且研究了表达产物的酶学性质及酶活随发酵pH的变化情况,为在酿酒工业中提高白曲酸性蛋白酶活力找到一个具有参考意义的方法。
1 材料与方法
1.1 材料
1.1.1 菌株与质粒
黑曲酶SH-2和pMD18表达载体由本实验室改造并保存,pepB目的片段由广州艾基生物技术有限公司合成。
1.1.2 工具酶、试剂盒和试剂
PCR反应预混酶 Prime STAR HS(premix)购自TaKaRa公司;抗生素Amp购自北京普博欣生物科技有限公司;质粒小量提取试剂盒和质粒大量提取试剂盒购自广州捷倍斯生物科技有限公司;胶回收试剂盒购自美国Omega公司;福林酚试剂购自北京普博生物科技有限公司。
1.1.3 培养基
CD培养基(g/L):葡萄糖20,NaNO33,KCl2,MgSO4·7H2O 0.5,KH2PO40.01,pH 5.5(固体含 1.7%琼脂,液体含0.05%琼脂)。
蔗糖高渗培养基(g/L):蔗糖 400,NaNO33,KCl2,MgSO4·7H2O 0.5,KH2PO40.01,pH 5.5(软琼脂含0.5%琼脂,固体含1.7%琼脂)。
淀粉发酵培养基(g/L):玉米淀粉50,玉米浆30,豆粕粉20。
淀粉培养基(g/L):牛肉膏3,蛋白胨10,酵母粉2,NaCl 2,可溶性淀粉5,pH 5.5。
1.1.4 引物
实验所用的引物如表1所示。

表1 引物Table 1 Primer
1.2 方法
1.2.1 pMD18-pepB表达载体构建
表达载体图谱如图 1。以黑曲霉基因组为模板PCR扩增糖化酶启动子PglaA,以广州艾基生物技术有限公司合成的pepB质粒为模板扩增 pepB片段。PCR反应条件:98 ℃预变性10 min;98 ℃变性30 s,68 ℃退火30 s,72 ℃延伸90 s,共30个循环;第30个循环72 ℃延伸7 min。PCR产物经1%凝胶电泳鉴定,将鉴定正确的PCR产物凝胶回收。PglaA、pepB、线性化后带有表达元件的pMD18-T三片段在Infusion酶作用下进行连接,55 ℃反应,15 min后得到连接液。用热激法将连接液转入大肠杆菌感受态Match1 T1,涂板于含有Amp抗生素的LB平板上,过夜培养后将阳性转化子液体LB培养12 h。然后进行菌液PCR鉴定,挑取条带正确的转化子,送到广州艾基生物技术有限公司测序。
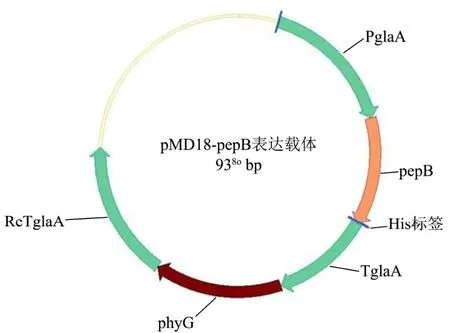
图1 pMD18-pepB表达载体图谱Fig.1 Map of pMD18-pepB expression vector
1.2.2 PEG介导转化
黑曲霉常用的遗传转化方法包括孢子电转化法、农杆菌介导转化法、原生质体介导的转化法等[9]。Meyer[10]对这些转化方法的利弊进行了系统的分析,认为孢子电转化法操作简单,但转化效率低。农杆菌介导转化虽具有转化受体形式多样、转化效率较高、转化子稳定、可转移大片段、单拷贝比例高等优点,但其操作复杂,周期长,转化过程受多种因素影响。Michielse等[11]明确指出了农杆菌介导转化方法在黑曲霉中的转化效率较低。原生质体PEG介导转化具有操作简单,转化效率高的特点,对黑曲霉菌株而言是较为有效的遗传操作方法。李秀鹏[18]、许晓红[12]用PEG介导转化法在无孢黑曲SH-2中都得到了成功实践。
原生质体的制备及转化步骤详见参考文献[9,13]。转化实验设置如下:实验组:将制得的原生质体、pMD18-pepB质粒和PEG缓冲液的转化液加入到CD高渗软琼脂培养基中,混合均匀后,铺在CD高渗培养基上,30 ℃培养观察。再生组:同实验组,唯一不同的是将CD高渗软琼脂培养基中预先加入尿嘧啶核苷,该实验组是检验原生质体的制备效果,30 ℃培养观察。阴性组:同实验组,唯一不同的是将上述pMD18-pepB质粒用无菌水代替,30 ℃培养观察,记录生长情况。
1.2.3 转化子鉴定
随机实验组CD高渗培养基上至普通固体CD板进行扩大培养,5 d后挑取菌落,提取基因组,进行PCR鉴定。鉴定引物设置三对:gyrG-F/gyrG-R、pepA-F/pepA-R、PglaA-F/PglaA-R之后将定位都正确的转化子保种。
1.2.4 转化子发酵
发酵罐投料(30 L):玉米浆3%、豆饼粉2%、玉米淀粉2%、麦芽糖浆(分消)3%、消泡剂 30 mL。除玉米浆和分消外,将原料投入发酵罐,调pH 6.0~6.2,加中温淀粉酶和高温淀粉酶,70 ℃保温 20min,90 ℃保温30 min,加入玉米浆,121 ℃~123 ℃,灭菌35 min。
发酵罐发酵相关参数:罐压:0.05~0.06 MPa;罐温:初始 34 ℃;转速:200~800 r/min(根据溶氧可调);溶氧:前期控制50%以上,中后期按最大转速、风量运转;pH:初始pH 5.5,当pH降至4.5时,补氨控制在4.8。在发酵48 h、72 h、96 h、120 h、144 h、168 h、192 h、216 h、240 h发酵上清液5 mL,-20 ℃保存。
1.2.5 发酵初始pH对产酶的影响
淀粉发酵培养基100 mL,分装于500 mL锥形瓶中,用磷酸缓冲液调节pH分别至4.5、5.5、6.5,115 ℃灭菌。分别接种液体CD培养的重组菌株菌液1 mL于淀粉发酵培养基中,于30 ℃、250 r/min条件下培养,分别在摇瓶24 h、48 h、72 h、96 h、120 h、144 h、168 h取发酵上清液5 mL,-20 ℃保存。
1.2.6 酶活测定与SDS-PAGE分析
采用Folin法测定蛋白酶酶活,以酪蛋白为底物,将其用乳酸缓冲液溶解,终浓度为10.0 g/L。反应体系为4 mL,将1 mL酪蛋白溶液加入1 mL酶液中42 ℃条件下反应10 min,加2 mL三氯乙酸终止反应,对照组先向酶液中加入三氯乙酸以使酶失活。取过滤液1 mL加碳酸钠溶液5.0 mL,再加福林试剂1 mL,42 ℃显色20 min,于680 nm波长测定吸光度,根据标准曲线计算酶活。酶活定义:温度42 ℃条件下,1 min水解酪素,产生1 μmol酪氨酸所需要的酶量为一个酶活力单位。
重组转化子发酵上清液清液40 μL,加入10 μL SDS loading,加热煮沸5 min,取10 μL点样,进行4~20% SDS-PAGE电泳,考马斯亮蓝R 250染色,脱色后检测目的蛋白条带,分析重组蛋白分子量。SDS-PAGE电泳方法参考《分子克隆手册》。
2 结果与分析
2.1 pMD18-pepB表达质粒构建
分别以黑曲霉基因组和含有pepB的质粒为模板,PCR得到PglaA片段和pepB片段。PCR产物凝胶电泳如图2所示,分别在1500 bp附近和1000 bp~1200 bp处扩增出特异性条带,与预期的PglaA片段(1500 bp)、pepB片段(1185 bp)大小相符。PglaA片段和pepB片段进行重叠PCR,产物片段回收。PglaA、pepB和pMD18连接后转入大肠感受态细胞,挑取阳性转化子,液体LB培养后,进行菌液PCR,电泳结果如图3,2号泳道菌液在10000 bp附近出现特异性条带,与pMD18-pepB质粒(9380 bp)大小相符。将该菌液送广州艾基生物技术有限公司测序,比对测序结果无基因突变,表明pMD18-pepB表达质粒构建成功,可以用于转化曲霉。

图2 PglaA片段和pepB片段PCR扩增Fig.2 PCR amplification of PglaA fragment and pepB fragment

图3 菌液电泳Fig.3 Escherichia coli electrophoresis
2.2 转化子鉴定
在丝状真菌遗传操作中,pyrG是常用的选择性标记,其编码的乳清苷-5-磷酸脱羧酶(OMP)尿嘧啶核苷酸合成途径中的关键酶,pyrG缺失菌株不具备尿嘧啶核苷酸合成能力。宿主无孢黑曲SH-2是gyrG缺陷菌株,pMD18-pepB表达载体带有pyrG,因此阳性转化子可以在不含尿嘧啶核苷的普通CD板上生长。转化后培养10 d,从实验组转化板上随机挑阳性转化子,至普通 CD板。提取基因组,用三对引物pepB-F/pepB-R、PglaA-F/PglaA-R、gyrG-F/gyrG-R进行PCR鉴定。PCR产物凝胶电泳结果如图4,对应的pepB、PglaA、gyrG泳道分别在1000 bp~1200 bp、1400 bp~1600 bp、1000 bp~1200 bp处扩增出了明亮的特异性条带,这与预期的 pepB(1185 bp)、PglaA(1500 bp)gyrG(1100 bp)条带大小相符,说明目的基因表达框整合到了黑曲霉 SH-2基因组上。鉴定正确的转化子用于发酵培养,以检测目的蛋白。

图4 转化子PCR鉴定Fig.4 PCR identification of transformant
2.3 表达产物检测
将定位正确的阳性转化子在 30 L发酵罐中进行发酵,取48 h、72 h、96 h、120 h、144 h、168 h、192 h、216 h、240 h发酵上清液,稀释 10倍后,进行SDS-PAGE检测及酶活测定。如图5,SDS-PAGE分析,47 ku附近处均有条带,与已知报道的酸性蛋白酶条带相符[14,15],这说明白曲霉酸性蛋白酶与黑曲霉和宇佐美曲霉来源的酸性蛋白酶在分子量上没有明显差别。随着发酵时间的增加,酸性蛋白酶条带逐渐加深,并且酶活逐渐增加。在48 h~96 h内,酶活增加较缓慢,96 h后酶活呈直线趋势增加,在发酵240 h后,发酵粗酶液酶活达9972 U/mL。这与杨猛等[8]对白曲原始菌株进行优化培养所得酸性蛋白酶酶活数据相比,酶活显著提高,是原始菌株的 8.5倍。李杰等[7]用基因工程技术,使白曲酸性蛋白酶基因在黑曲CICC2462中得到了表达,重组菌株酶活为 5543 U/mL,与之相比,本实验所得的重组菌株酶活提高了79.9%,与文献报道[5,6,16,17]的其他来源的酸性蛋白酶酶活数据相比,也存在一定的优势,这主要与本研究中采用的宿主有关。黑曲酶具有很强的糖化酶分泌能力,使得其内源表达自身的糖化酶过程中,消耗掉很多中间代谢物和能量,同时过量表达的糖化酶会对内质网和高尔基体等蛋白分泌途径产生一定的压力,若不能正确及时加工修饰成成熟蛋白分泌到胞外,将严重影响到其他蛋白的生成和分泌,限制外源蛋白的表达[18]。本研究中采用的宿主菌无孢黑曲SH-2,已敲除多拷贝糖化酶基因,为外源蛋白的表达提供了有力条件。同时,从重组菌株发酵液SDS-PAGE图谱可以看出,其他分泌蛋白表达量很少,这为其作为添加菌用于酿酒工业提高白曲酸性蛋白酶酶活提供了可能。

图5 白曲酸性蛋白酶SDS-PAGE分析及酶活测定Fig.5 Analysis of SDS-PAGE and enzymatic activity determination of acid protease
2.4 酶学性质分析
2.4.1 最适温度

图6 温度对酶活的影响Fig.6 Effect of temperature on enzyme activity
水浴锅设置 25 ℃、30 ℃、35 ℃、40 ℃、45 ℃、50 ℃、55 ℃ 7个温度梯度。在pH恒定条件下,测定在不同催化温度下的酶活。结果如图6所示,最适反应温度为35 ℃,在25 ℃~55 ℃范围内,酶活都保持在60%以上。酸性蛋白酶的活性一般在50 ℃以下可以保持稳定,但也随产酶的微生物菌源不同而有所差异[19]。黑曲霉SL22-111所产酸性蛋白酶最佳反应温度为 50 ℃。低于 45 ℃或高于 60 ℃,酶活急剧下降,70 ℃时酶活性完全丧失[14],米曲霉MTCC5341所产酸性蛋白酶最适温度为55 ℃,在30 ℃~60 ℃时酶活力较稳定[20],宇佐美曲霉L336酸性蛋白酶最适温度较高,在 60 ℃以上[21]。因此白曲酸性蛋白酶与其他来源酸性蛋白酶相比,最适反应温度偏低。

图7 pH对酶活的影响Fig.7 Effect of pH on enzyme activity
2.4.2 最适pH
用K2HPO4和KH2PO4不同配比得到pH为2.5、3.0、3.5、4.0、4.5、5.0、5.5的磷酸缓冲液,然后用其稀释酶液,并配成不同pH的1%酪蛋白底物,在恒定温度下,测定酶活。结果如图7,可以看出当pH为4时,酶的活性最高,pH低于或高于4,酶活都会降低。pH值对白曲酸性蛋白酶活力影响非常显著,pH值小幅下降会引起酶活迅速下降,当pH值为2.5时,酶基本失活,这均与酶结构改变有关。与丛毛红曲霉M.pilosus胞外酸性蛋白酶规律相似[22],pH值改变导致酶蛋白空间构象改变,甚至引起酶变性失活。在最适pH值时,酶的解离状态往往是最有利于该酶与底物结合并发生催化反应,使酶活力达到最大。袁康培等[23]对黑曲霉HU53酸性蛋白酶进行研究,得出其最适反应pH为3.0、曾黎明等[24]报道的米曲霉酸性蛋白酶最适pH为6.0,杨琥[5]对宇佐美曲霉酸性蛋白酶酶学性质研究,表明其最适反应pH为2.5。不同来源的酸性蛋白酶最适反应pH不同,可能与其酶活性中心含有的羧基有关[25]。
2.4.3 金属离子的影响
探究 Mn2+、Zn2+、Mg2+、Cd2+、Cu2+对酶活的影响,使以上各离子分别溶解于酶液中,终离子浓度为5 mmol/L。如图 8,CK为空白对照组,可以看出,Mn2+、Cu2+对酶活有显著的激活作用,酶活分别是对照组的3倍和2.5倍,Zn2+、Mg2+对酶活的影响作用不明显,而Cd2+则对酶有明显的抑制作用,使酶活低于对照组的 50%。戚淑威[15]研究的结果表明,Mn2+对该酶有明显的激活作用,可使原始酶活提高2倍以上,张秀江[17]研究表明,在 5.0 mmol/L的浓度下,Mn2+对黑曲霉所产酸性蛋白酶有强烈的激活作用,达到对照酶活力的 150%。可见,作为能显著提高酸性蛋白酶酶活的金属离子,Mn2+具有很大的应用潜力。

图8 金属离子对酶活的影响Fig.8 Effect of ion types on enzyme activity
2.4.4 发酵初始pH对产酶的影响
用磷酸缓冲液调节淀粉发酵培养基的pH,来研究pH对产酶的影响。


图9 不同发酵初始pH下的酶活测定Fig.9 Determination of enzyme activity under different initial pH of fermentation
设置pH 4.5、pH 5.5、pH 6.5三个梯度,取24 h、48 h、72 h、96 h、120 h、144 h、168 h发酵上清液样品。SDS-PAGE分析,结果如图9,a、b、c对应的发酵初始pH条件分别为pH 4.5、pH 5.5、pH 6.5,d相应的酶活测定。可以看出,在时间维度上,随着发酵时间的增加,酸性蛋白酶表达量及酶活性逐渐增加。在初始pH 4.5~6.5范围内,提高发酵液初始pH,表达量及酶活性会有所增加。与方春玉等[4]、袁康培等[23]的报道相比偏高,这可能与宿主菌的生长最适条件有关。本研究采用的宿主无孢黑曲SH-2最适生长pH为5.5[18],在其发酵过程中酸度会积累,从而解释了发酵初始pH为6.5酶活会更高的情况。在酸性范围内,适当地增加重组菌株发酵液初始pH有利于酶活的提高。
3 结论
3.1 有关白曲酸性蛋白酶表达研究的报道比较少,本研究利用基因工程技术,使白曲霉酸性蛋白酶基因,在无孢黑曲SH-2中得到了表达。经30 L发酵罐发酵240 h后,重组菌株发酵粗酶液酶活为9972 U/mL。酶活性质研究表明,该酶的最适温度为35 ℃,最适pH为 4,Mn2+、Cu2+对酶活有显著的激活作用。在 pH 4.5~6.5范围内,适当增大pH可以提高重组菌株在摇瓶发酵条件下酸性蛋白酶酶活。
3.2 本研究所用的宿主无孢黑曲SH-2经过了糖化酶基因多拷贝的敲出,具有高表达、背景干净的特点。重组菌株粗发酵液酶活是出发菌株酶活的8.5倍,与李杰等报道相比,酶活提高了 79.9%,与报道的其他来源的酸性蛋白酶表达也存在一定的优势,并且重组菌株背景单一,其他分泌蛋白表达量较少。因此,本研究所得的重组菌株有望用于白酒生产中调节糖化力和酸性蛋白酶活力的比例,来提高芝麻香酒的风味。
