蝗虫对蓝波谱不同偏光属性刺激效应的视偏响应效应测定
2018-07-05刘启航孔晓红付素芳杜家熙
刘启航 孔晓红 付素芳 杜家熙 周 强
(1河南科技学院机电学院, 新乡453003; 2中国农业大学工学院, 北京 100083)
0 引言
研究蝗虫灾害的光电波谱诱导捕集技术是实现现代物理农业的组成部分,该技术不仅能够实现农药使用量降低,还可以为其他农林牧害虫的物理防治提供有意义的借鉴,蝗虫视敏有效激发及视响应敏感的光波物理信息是解决问题的关键,但未能获得突破。目前,昆虫依据自然界偏振光场属性的定位导航功能研究,为蝗虫视觉诱导信息提供了重要借鉴,但昆虫偏光研究还限于偏振敏感神经探测控制机制及行为定向,因此,研究蝗虫偏光视励性光电诱导增益效应,有利于昆虫偏光响应机制及其在农业植保中的应用。
文献[1-4]研究表明,蝗虫等昆虫依赖复眼将光学能量转化为神经冲动电位,作为感受外界刺激的起点,并以膜外电位ERG来反映外界刺激强度,而蝗虫敏感波谱光能强度引起的视觉生理调控机制,在满足视觉功能的前提下产生趋光视觉行为,并引发蝗虫对光刺激信号的感应辨识和光照环境中的自身空间定位,但光谱识别力和条纹定向是2个明显不同的视觉行为,且对水平光栅目标的响应更强。同时,蝗虫偏振视觉具有E-矢量定向匹配功能,并能够监控响应天空的偏振类型,易造成生理神经的激发和改变活动行为,所以蝗虫等昆虫能够感知天空线偏光,并具有利用偏光进行定向的能力[5-7]。偏振光照对蝗虫偏振神经具有兴奋抑制、引起和增效作用,所以蝗虫对偏振光的响应取决于蓝光敏感光子接受器,并当外界环境亮度降低时,偏振光对蝗虫定向起着决定作用,且蝗虫对蓝光旋转交变偏振光场的响应强度比绿、蓝线偏振光场分别高25%和28%[8-11]。因此,研究蓝光偏振光场特征对蝗虫的光电诱导效应,获得蝗虫偏光响应中的视励性视敏增益机制及作用方式,能降低能量投入和提高诱导效果。
本文利用蓝光波谱光照、蓝光激光条纹光照及其不同特征的偏振光场,测试蝗虫视效性视敏响应效果,以揭示蝗虫视响应与视觉激励的相关关联、优化蝗虫视觉响应良好的激励增益机制及光场效应,以获得蝗虫趋光趋偏响应所需的光能物理特征和光波信息因素,并探讨蝗虫趋光趋偏响应机理,以期为蝗虫偏光响应及光响应的研究提供理论基础和方法参考。
1 材料和方法
试虫为河北邯郸棚内饲养羽化7 d内的东亚飞蝗健壮成虫。购买后试虫户外纱窗箱内以新鲜芦苇叶及竹叶饲养。室内进行试验,室温为27~30℃,试验采集时间为20:00—22:00。
试验装置如图1示,其中,视励光源共4组,黑光板制成的行为响应通道呈U型。
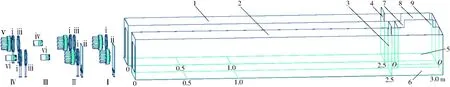
图1 蝗虫视觉响应不同光场特性的试验装置Fig.1 Experimental equipment of locust responding to different blue light propertiesⅠ.线偏蓝波谱光源组:i.线偏片,ii.线偏玻璃片 Ⅱ.退偏蓝波谱矢量光:i.固定偏振片,iii.退偏偏振片,ii.线偏玻璃片 Ⅲ.蓝波谱激光光源组:iv.横条纹光源,vi.竖条纹光源 Ⅳ.蓝波谱光源组:v.蓝光源,vi.激光蓝波谱竖条纹光照,i.固定偏振片,iii.退偏偏振片 1.行为响应通道1 2.行为响应通道2 3.闸门1 4.闸门2 5.蝗虫反应室1 6.蝗虫反应室2 7.反应室通道 8.闸门3 9.闸门4
Φ55 mm圆形LED蓝光源2个分别前置线偏片(透光率50%、起偏率95%)和线偏玻璃片(长×宽:200 mm×150 mm,透光率85%、起偏率25%)(Ⅰ组,形成起偏程度不同的线偏光和玻璃线偏光),Φ55 mm圆形LED蓝光源2个分别前置2个线偏片(固定线偏片i和矢量可调退偏偏振片iii)和线偏片与线偏玻璃片(固定线偏片i和线偏玻璃偏片ii)(Ⅱ组,形成退偏程度不同的偏光和玻璃偏光,且固定线偏片2′的起偏矢量相同),测试起偏程度不同的线偏光和退偏程度不同的退偏光的偏光属性视励效果;1.8 W蓝激光光源2个(Ⅲ组,调整聚光头分别形成蓝波谱横条纹光iv及竖条纹光vi,其中,条纹间距及条纹宽度在0.5 m内为5 mm),测试激光条纹光的蝗虫视励性趋光效果;Φ55 mm圆形LED蓝光源和1.8 W蓝激光竖条纹光源前端分别前置2个偏振片(固定线偏片i和矢量可调退偏偏振片iii)(Ⅳ组,形成光质属性不同的偏光),验证及讨论蝗虫偏光响应的矢励性矢敏效应。
Ⅰ~Ⅳ视励光源组分别置于行为响应通道1、2前端,调整相互间距,保证光照效果,465 nm蓝光光源及激光条纹光源供电电压分别为12 V和5 V。
为确定蝗虫的视响应效应及其视行为的光质调控效应,U型通道中,行为响应通道1与蝗虫反应室1、行为响应通道2与蝗虫反应室2分别制成直通道(总长:3.0 m,宽×高:0.5 m×0.4 m),由闸门在2.5 m处分隔形成视响应通道及蝗虫反应室,通道划分成如图1所示区段。为确定反应室1和2内的光照对蝗虫视响应的影响,反应室1、2由中间O-O标示的反应室通道(长×宽×高:0.3 m×0.4 m×0.4 m)相连。
为对比确定蝗虫对蓝光不同光质的视响应效应,针对每一光源组,30只试虫为一组并备2组。
首先,利用Ⅰ型光源组,2组试虫分别置于反应室1和反应室2内进行测试,测2次后2组试虫互换反应室再测2次(光源组中每一光质均共测4次,以避免不可比性),以确定线偏光质对蝗虫的视励性视敏效应。然后,在Ⅱ型光源组中,相对于固定偏振片i,利用矢量分度架调整偏振片ii形成矢量不同的试验用退偏光(试验中,选取矢量角度为0°、30°、90°、120°、180°、210°、270°、330°[12]),并利用偏振片i与线偏玻璃片形成的玻璃退偏光,针对每一矢量退偏光和玻璃退偏光,利用上述方法进行测试,以确定退偏光及玻璃退偏光对蝗虫的视励效应(Ⅰ和Ⅱ中光源光照度为100、1 000、10 000 lx)。
其次,针对Ⅲ型光源组(光照度为100 lx)中的横、竖激光条纹光照,利用上述相同方法进行测试,以优选蝗虫视敏良好的视励信息,在此基础上,利用Ⅳ型光源组形成的蓝波谱竖条纹和蓝波谱退偏矢量光照(二者光源对应的光照度分别为100、10 lx,退偏矢量同上),利用上述相同方法进行测试,以验证和探讨偏光不同光质对蝗虫视偏响应效应的视励效果和蝗虫偏振视觉的视敏光质属性。
为测试蝗虫对不同光质属性的视响应效应,试验前,试虫群体置于蝗虫反应室内适应。试验时,开启光源10 min及闸门1~4。试验中,两次试验间隔为20 min,以确保蝗虫视觉恢复,而且,两组试虫在不同测试次数中对相同光属性的视响应效应结果在0.01水平上差异不显著(F检验)。通过观察,反应室1和2内蝗虫的避光效应不显著,且相互光照作用下,避光蝗虫过O-O界线的虫数不影响试验结果。试验后,关闭光源及闸门,针对光源组中每一光质特性对应的响应通道,分别记录各区段内虫数。依据每次记录的试验数据,计算各通道0~0.5 m、0~1.0 m、0~2.5 m内总次数的虫数均值数与30只虫数的百分比,分别确定视趋强度、视聚集效应、视响应程度,用这3个指标来反映蝗虫对不同光质属性的视敏性视偏响应效应。
试验数据采用Excel软件和SPSS 16.0数据处理系统进行数据统计分析,不同光源处理间差异显著性采用F检验,多重分析采用LSD测验。试验结果均值标准误差为±(2.5%~5.0%),不同次试验处理间差异不显著(F检验,P>0.05)。
2 结果与讨论
2.1 试验结果
2.1.1蝗虫对蓝光线偏不同光质光照的视偏响应效应
蝗虫对不同线偏光质的视响应结果如图2所示。
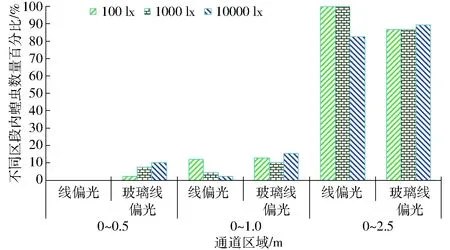
图2 不同光照度的线偏光质视偏响应效应Fig.2 Response effect of locust for different linear polarized lights
由图2可知,线偏光中,光照度递增性刺激效应未能引起蝗虫趋向于0~0.5 m区段内,而光照度递增,蝗虫视聚集效应呈递减性,且光源光照度100 lx与1 000 lx相比,蝗虫视响应程度无差异而10 000 lx对应的视响应程度显著低于100 lx及1 000 lx;玻璃线偏光中,光源光照度递增,蝗虫的视趋强度增强,而1 000 lx的视聚集效果最差,10 000 lx的视聚集效果最优,且光照度递增,蝗虫视响应程度的变化不显著。经对比,蝗虫对玻璃线偏光的视趋强度及视聚集效果优于线偏光,而100 lx及1 000 lx时蝗虫对线偏光的视响应程度显著高于玻璃线偏光,而10 000 lx时低于玻璃线偏光。
由于偏振片线偏光的偏振效应较高,玻璃线偏光的蓝光波谱光照效应较强,因而,在蝗虫的视偏响应效应中,蓝光线偏光的起偏效应决定蝗虫视响应程度的视敏性、线偏光照度制约蝗虫的视趋强度及视聚集效果,且较弱光照中单偏片线偏光照特性对蝗虫偏振视觉的视励性较优。
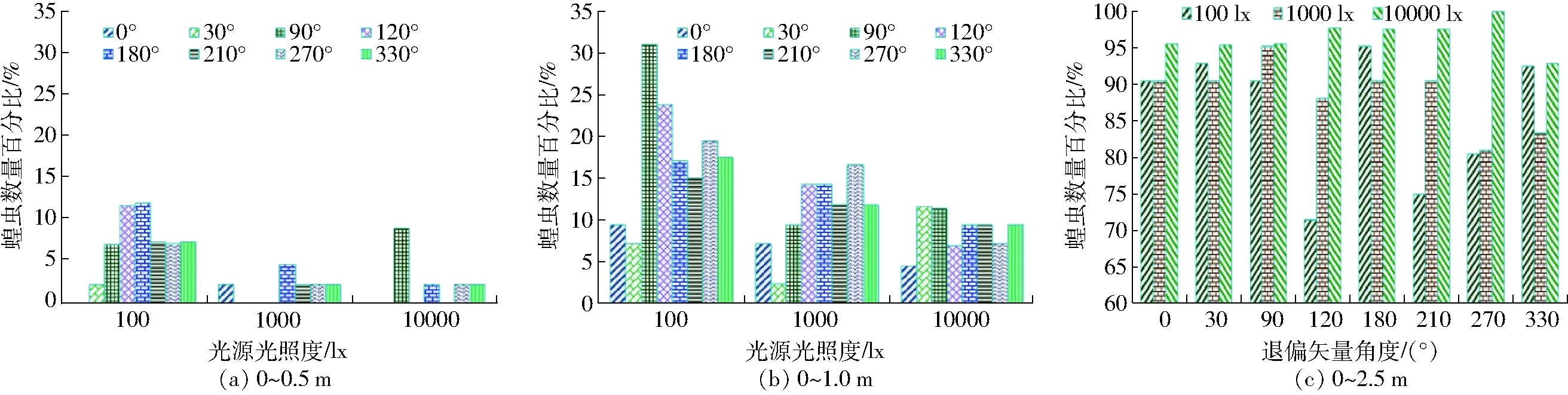
图3 不同退偏矢量角度的视偏响应效应结果Fig.3 Results of locusts response to different light illumination of polarized light
2.1.2蝗虫对蓝光退偏光不同光质光照的视响应效应
蝗虫对单偏振片与线偏玻璃形成的玻璃退偏光、退偏矢量不同退偏光的视偏响应效应结果分别如表1和图3所示。
由表1可知,100 lx时蝗虫对玻璃退偏光的视趋强度及视聚集效果显著最优,且1 000、10 000 lx时玻璃退偏光未引起蝗虫产生视趋效应,随光照度递增,10 000 lx与1 000 lx相比,10 000 lx对应的视聚集效果较优,10 000 lx对应的视响应程度最高,但与100 lx相比,差异性不显著(P>0.025)。经比较,光源光照度为1 000 lx时蝗虫的视偏响应效应最差,100 lx时视偏响应效应最优。
由图3可知,100 lx及10 000 lx时,退偏0°矢量光照未引起蝗虫产生视趋效应,且退偏90°矢量光照中10 000 lx对应的视趋强度高100 lx 2个百分点,而其余相同矢量光照中,100 lx对应的视趋强度较优,且1 000 lx与10 000 lx相比,二者对应的视趋强度差异性不显著(P>0.025),经比较,100 lx时,蝗虫对退偏120°及180°矢量光照的视趋强度最优。退偏30°矢量光照中10 000 lx对应的视聚集效应较优,其余相同矢量光照中,100 lx对应的视聚集效应较优,且90°矢量光照中10 000 lx次优而其余相同矢量中1 000 lx次优,经比较,100 lx时,蝗虫对90°和120°的视聚集效应分别最优和次优。但10 000 lx时蝗虫对相同矢量退偏光照的视响应程度较优并以270°矢量最优,而蝗虫对120°、180°、210°的视响应程度无差异且次优。
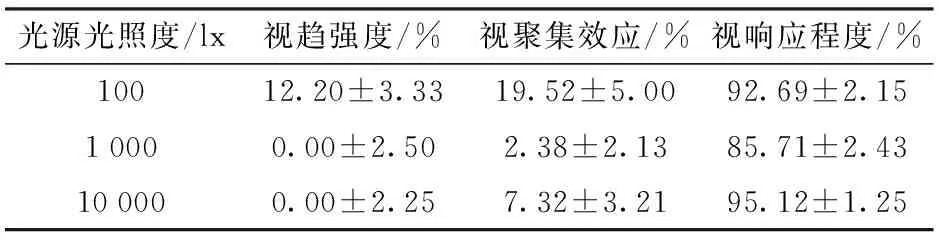
表1 蝗虫对单偏振片与线偏玻璃耦合性玻璃退偏光的视响应效应结果Tab.1 Results of locust response to polarized light formed by single polarizer and glass
注:表中数据为平均值±标准误差,不同处理间差异不显著(P>0.05)(F检验)。
玻璃退偏光中,100 lx对应的视趋强度及视聚集效应较优、10 000 lx对应的视响应程度较强(表1)。经测定和计算,退偏180°矢量光照的偏振度和光照度低于玻璃退偏光,而100 lx时蝗虫对玻璃退偏光和退偏180°矢量光的视趋强度相同。100 lx时退偏0°矢量光照的光照度与其180°相同,但蝗虫对0°矢量光照的视趋强度最低。100 lx时退偏120°矢量光照的光照度低于180°矢量光照度,但两者的视趋强度差异性不显著(P>0.01)。100 lx时,玻璃退偏光的光照度及偏振度均高于90°、270°退偏光,但蝗虫对90°及270°退偏光的视聚集效应优于玻璃退偏光,且退偏矢量光照中,蝗虫对90°退偏光的视聚集效应、对270°退偏光的视响应程度最优,并均以120°次优。
因而,偏光偏振度及E-矢量光照度影响退偏光质属性对偏振视觉视激性敏感效应的视励强度,并呈现强化或抑制性作用效应。经对比,100 lx时,蝗虫对180°退偏光的视趋强度、90°退偏光的视聚集效应、270°退偏光的视响应程度最优,且视偏响应效应均以120°次优。
2.1.3蝗虫对蓝波谱激光条纹及其退偏矢量光的视偏响应效应
利用蝗虫视偏响应效应较优光照度(100 lx),蓝波谱条纹光照中,蝗虫对竖条纹光照的视响应效应显著优于横条纹(图4),且蝗虫对竖条纹蓝波谱偏光的视偏响应结果如图5所示。
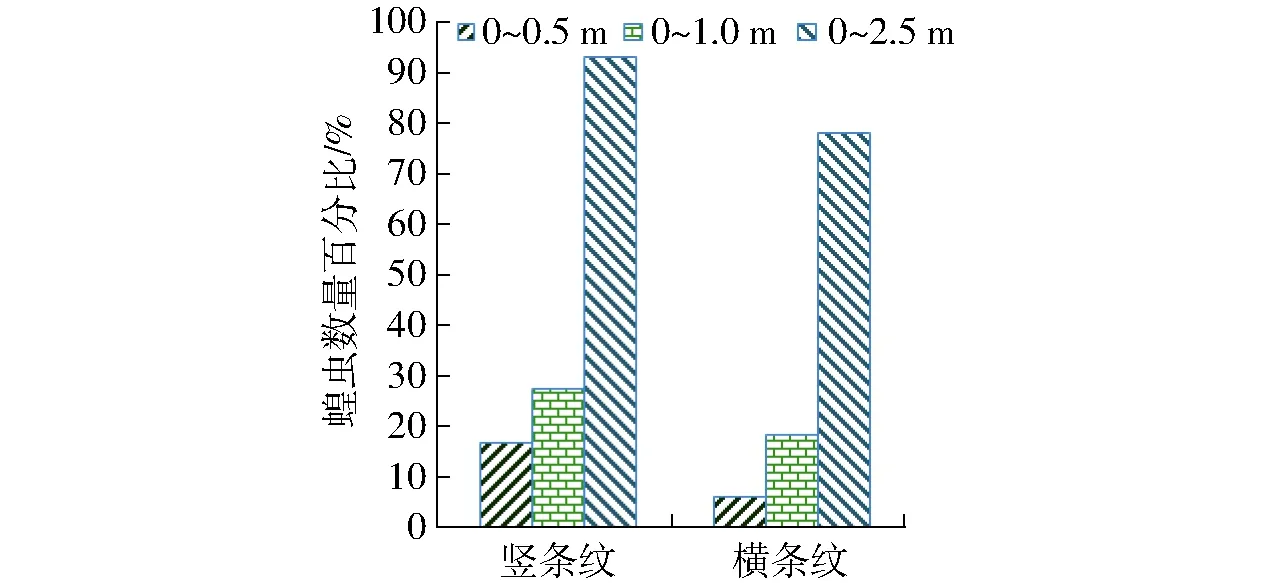
图4 蝗虫对横竖条纹光照的视响应结果Fig.4 Results of locust response to stripes light
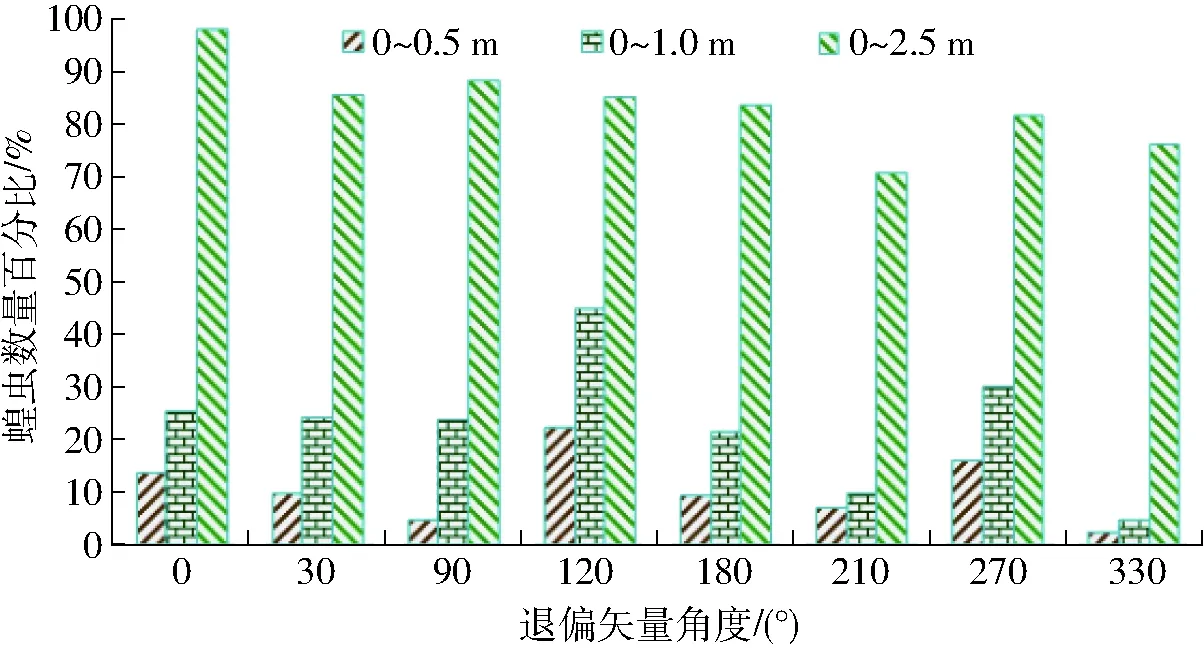
图5 蝗虫对竖条纹退偏矢量光照的视偏响应效应结果Fig.5 Results of locust response to vertical stripe of polarized light
竖条纹退偏矢量光照与竖条纹光照(图4和图5)相比:蝗虫对竖条纹120°退偏矢量光照的视趋强度优于竖条纹光照,而对其余矢量光照的视趋强度均低于竖条纹光照,且竖条纹退偏矢量光照中蝗虫对270°的视趋强度次优而330°较差;蝗虫对竖条纹120°及270°退偏矢量光照的视聚集效应优于竖条纹光照,而对其余矢量光照的视趋强度均低于竖条纹光照,且蝗虫对120°矢量光照的视聚集效应较优而330°较差;蝗虫对竖条纹0°退偏矢量光照的视响应程度较优,而对其余矢量光照的视响应程度均低于竖条纹光照且210°较差。经对比,蝗虫对竖条纹120°退偏矢量光照的视趋强度及视聚集效应最优而270°均次优,且蝗虫对0°的视趋强度及视聚集效应比270°约低2.5%,但蝗虫对竖条纹0°退偏矢量光的视响应程度最优。
鉴于竖条纹蓝偏光照度低于条纹光照并具有退偏效应,以及蝗虫偏振视觉视敏属性与光质激活视偏神经的匹配敏感性[13],因而,蝗虫对竖条纹120°及270°退偏矢量光照的视聚集效应、对0°的视响应程度优于条纹光,源于蓝波谱光照的激发性、偏光矢量的激励性、蝗虫辨识竖条纹光照的视敏性,三者的叠加调控效应对蝗虫视偏响应的耦合强化作用,但蝗虫对其余矢量光照的视偏响应效应均低于条纹光照,表明蝗虫偏振视觉的视励性与蝗虫趋光视觉的视激性呈现波谱光照激发操纵性视敏差异。
2.2 讨论
研究表明,蝗虫偏振视觉包含有对偏光方向极度敏感的感光器,并通过大量的小眼阵列和神经元对整个偏光模式进行接收而响应,且视响应敏感性取决于蓝光光子接受器[14-15],结果表明,蓝波谱线偏光质、蓝波谱退偏光质的刺激效应均引起蝗虫产生了视偏响应,但蓝波谱偏光光质光照特性不同,蝗虫视偏响应效应的视敏性不同,且在视响应程度中,100、1 000 lx时线偏光和10 000 lx时退偏270°矢量光诱发的视敏性较优,而10 000 lx时退偏330°矢量光诱发的视敏性较弱。MAPPES[16]指出,蝗虫的偏光响应强度与光梯度无关,当试验中光照度增强,线偏光梯度显著抑制而玻璃线偏光增效蝗虫的视趋强度,且线偏光光照度增强,抑制蝗虫视聚集效应的视敏性,而光照度相同时,蝗虫对玻璃线偏光的视敏性优于线偏光。
线偏光透过玻璃偏振片、偏振片,都会使退偏效应增强,而100 lx时蝗虫对退偏光的视趋强度及视聚集效应较强,1 000、10 000 lx时较弱,且相对于100 lx,1 000、10 000 lx时玻璃退偏光显著抑制视趋强度而退偏光对视趋强度的抑制性呈现光矢敏感调控性差异,该结果与较弱光环境中偏振光对蝗虫定向响应起决定作用结果相符,但100 lx时蝗虫的视趋强度未与E-矢量偏振模式的选择响应敏感性相匹配[17-18]。蝗虫对180°退偏光的视趋强度与玻璃退偏光无差异,而其余矢量的视励性视趋强度均低于玻璃退偏光,其可能与蝗虫中央复合体内神经元轴突在L5和R4有很多分枝对偏光偏振效应的敏感差异性相关[19]。
100 lx时蝗虫对E-矢量退偏光质光照的视聚集效应呈现弦函交变式视响应特性,而退偏光光质属性不同,蝗虫的视聚集效应不同,其源于蝗虫偏振视觉对偏光不同光质的偏振类型敏感识别差异及对偏光刺激属性的生物调控效应差异,并导致100 lx时,蝗虫对90°、120°退偏光质的视聚集效应优于玻璃退偏光。而退偏光光照强度增强,其对蝗虫偏振视觉的刺激强化效应,致使10 000 lx时蝗虫对退偏矢量光的视响应程度优于玻璃退偏光(330°除外)。
蓝波谱偏光光质中,偏光偏振效应及偏光光质属性决定而偏光光照强度制约蝗虫视励性视趋强度及视敏性视聚集效应,且弱光照中蝗虫偏振视觉的视励性视趋强度较强并呈现光矢敏感差异性,而蝗虫视聚集效应与偏光偏振度、偏光光照度分别呈正、负相关性,但100 lx时线偏光偏振效应增效蝗虫视响应的敏感活性,且蝗虫对退偏光视偏响应的视敏性与偏光能量强度视励效应显著相关。同时,100 lx时,利用退偏矢量光照对蝗虫趋偏视觉的偏振视励效应和竖条纹光照对蝗虫趋光视觉的视识激发性,耦合形成的视励视激效应,使蝗虫对竖条纹退偏120°矢量光的视趋强度高于退偏最优矢量光照(180°)且其视聚集效应高于退偏最优矢量光照(90°),而其视响应程度低于100 lx时的线偏光,且10 000 lx时蝗虫对退偏270°矢量光的视响应程度最优。
蝗虫偏振视觉对弱光照退偏蓝光具有良好的视偏响应效应,利用10 lx蓝光光源光照度对蝗虫偏光视响应进行了研究,结果如图6所示。
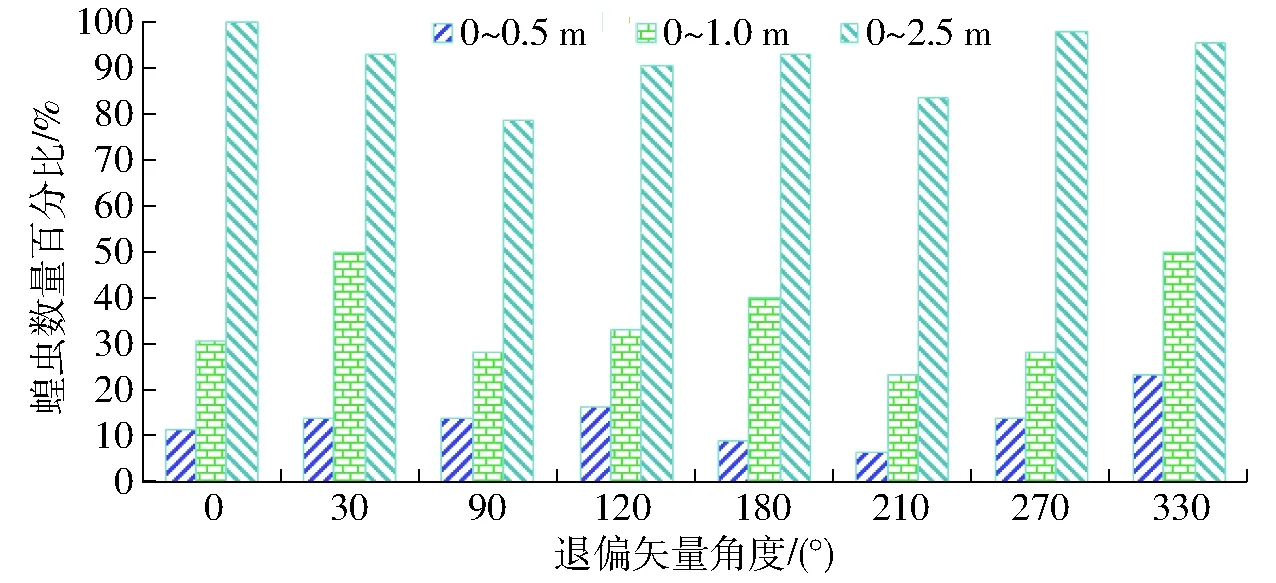
图6 蝗虫视觉响应10 lx偏光光照的结果Fig.6 Results of locust response to 10 lx polarized light
由图5、6可知,10 lx时退偏光中蝗虫视趋强度最优矢量(330°)、次优矢量(120°)的视励性分别优于100 lx时竖条纹退偏最优矢量(120°)、次优矢量(270°),并分别优于100 lx退偏光最优矢量(180°)、次优矢量(120°),且10 lx时蝗虫对330°、120°矢量的视趋强度分别最优、次优。10 lx时退偏光中蝗虫视聚集效应最优矢量(30°、330°)及次优矢量(180°)的视敏性优于100 lx时退偏光中视敏性
最优矢量(90°),并分别优于竖条纹偏光中视敏性最优(120°)、次优(270°)矢量,且10 lx时蝗虫的视聚集效应高于100 lx时相同矢量的退偏光及竖条纹退偏光,并以10 lx时退偏光30°及330°矢量最优而100 lx时竖条纹退偏光120°次优。10 lx时退偏光中0°矢量及100 lx时退偏光中270°矢量对蝗虫视响应程度的激发性最优。
图6结果表明,蝗虫对10 lx偏光的视偏响应效应呈现矢量变化性弦函响应特征,其验证了100 lx时退偏光中视聚集响应的E-矢量弦函响应特性的正确性,且与偏光诱发的蝗虫偏航力矩变化规律相符[20-21],因而,在弱光照蓝退偏光中蝗虫偏振视觉的E-矢量偏振模式的响应敏感性较显著,并具有视偏神经最大激活的矢量敏感函数,但蝗虫对其余偏光波谱光质特性的偏振敏感效应是否与蓝偏光相同,尚需探讨。
3 结论
(1)线偏蓝光偏振效应提高,其光强递增性刺激效应抑制蝗虫视趋强度及视聚集效应,而线偏弱光中蝗虫视响应程度的敏感性较优。退偏光光照效应中,蝗虫对弱光照偏振效应的视偏响应敏感性较强,且较强光照抑制视偏响应效应的敏感性,并呈现偏光E-矢量调控性视励视激性视敏差异。
(2)100 lx光源光照条件下,竖条纹蓝光照与偏光矢量耦合刺激强化措施,致使其最优(120°)及次优(270°)矢量对蝗虫视趋强度的视励性相对于退偏光最优矢量(180°)增强,且其最优(120°)矢量对蝗虫视聚集效应的视敏性增强相对于退偏光最优矢量(90°)增强。蝗虫视趋强度及视聚集效果以10 lx时330°偏光最优,而蝗虫视响应程度以100 lx时退偏270°矢量和10 lx 时退偏0°矢量最优。
(3)蝗虫对10 lx偏光的视偏响应效应呈现矢变性弦函视响应变化规律,表明弱光照蓝退偏光中蝗虫偏振视觉的E-矢量偏振模式的敏感匹配性较显著且视偏神经最大激活的矢量敏感函数得以显现,而且,蝗虫视偏聚集及视趋效应与偏光偏振度及视励信息、偏光光照度分别呈正负相关性,且蝗虫偏振视觉对弱光照退偏蓝光具有良好的视响应程度。
1 魏国树, 张青文, 吴卫国,等. 棉铃虫蛾复眼光反应特性[J]. 昆虫学报, 2002, 45(3):323-328.
WEI Guoshu, ZHANG Qingwen, WU Weiguo, et al. Characteristic response of the compound eyes ofHelicoverpaarmigerato light[J]. Acta Entomologica Scinica, 2002, 45(3):323-328.(in Chinese)
2 WILLIAM T C.The effect of target orientation on the visual acuity and the spatial frequency response of the locust eye[J]. Journal of Insect Physiology, 1999,45:191-200.
3 刘启航,周强.东亚飞蝗对波谱光及其偏光对比选择性视励效应的测定[J/OL]. 农业机械学报, 2016,47(4):233-238. http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx? flag=1&file_no=201604031& journal_id=jcsam. DOI:10.6041/j.issn.1000-1298.2016.04.031.
LIU Qihang, ZHOU Qiang. Comparative investigation of locusts visual bio-selection response effect induced by incentive effect of polarized light and spectral light[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2016,47(4):233-238.(in Chinese)
4 LIU Q H, XIN Z, ZHOU Q. Visual reaction effects induced and stimulated by different lights on phototactic bio-behaviors inLocustamigratoriamanilensis[J]. Journal of Agricultural and Biological Engineering, 2017, 10(4): 173-181.
5 SCHMELING F, WAKAKUWA M, TEGTMEIAR J, et al. Opsin expression, physiological characterization and identification of photoreceptor cells in the dorsal rim area and main retina of the desert locust,Schistocercagregaria[J]. Journal of Experimental Biology, 2015, 217:3557-3568.
6 BECH M, HOMBERG U, PFEIFFER K. Receptive fields of locust brain neurons are matched to polarization patterns of the sky[J]. Current Biology, 2014, 24(18):2124-2129.
7 HEINZE S, HOMBERG U. Maplike representation of celestial E-vector orientations in the brain of an insect[J]. Science, 2007,315: 995-997.
8 刘启航,蒋月丽,周强.东亚飞蝗对LED光信号视觉响应的波谱视敏效应测定[J/OL].农业机械学报, 2016,47(2): 338-344. http:∥www.j-csam.org/jcsam/ch/reader/view_abstract.aspx?flag=1&file_no=201606041&journal_id=jcsam.DOI:doi:10.6041/j.issn.1000-1298.2016.06.041.
LIU Qihang, JIANG Yueli, ZHOU Qiang. Spectral vision acuity reaction detection of phototactic response ofLocustamigratoriato LED light signal[J/OL]. Transactions of the Chinese Society for Agricultural Machinery, 2016,47(2): 338-344.(in Chinese)
9 SCHMELING F, TEGTMEIAR J, KINOSHITA M, et al. Photoreceptor projections and receptive fields in the dorsal rim area and main retina of the locust eye[J]. Journal of Comparative Physiology A, 2015, 202:585-599.
10 刘启航,周强.蝗虫对蓝光偏振光和非偏振光定向响应的对比研究[J]. 农业机械学报,2011,42(12):135-139.
LIU Qihang, ZHOU Qiang. The study on locusts orienting and responsing to blue-polarized and unpolarized light[J]. Transactions of the Chinese Society for Agricultural Machinery, 2011,42(12):135-139.(in Chinese)
11 ANDREAS B, MARCEL S, ROWELL C. The perception of the visual flow field by flying locusts: a behavioral and neuronal analysis [J]. Journal of Experimental Biology, 1992,165: 137-160
12 HOMBERG U, HEINZE S,PFEIFFER K,et al.Central neural coding of sky polarization in insects[J].Philosophical Transactions of the Royal Society B, 2011, 366: 680-687.
13 KRAPP H G. Polarization vision: how insects find their way by watching the sky[J]. Current Biology, 2007, 17(14):557-560.
14 YANG Hongzhang, WEN Lizhang, YI Qian, et al. Effects of light on the phototaxis of several important agricultural pests[J]. Chinese Agricultural Science Bulletin, 2014,30(25):279-285.
15 HEINZE S, GOTTHARDTS S, HOMBERG U. Transformation of polarized light information in the central complex of the locust[J].The Journal of Neuroscience, 2009, 29(38): 11783-11793.
16 MAPPES M. Behavioral analysis of polarization vision in tethered flying locusts[J].Journal of Comparative Physiology A,2004,190: 61-68.
17 KERAM P, MICHIYO K, HOMBERG U. Polarization-sensitive and light-sensitive neurons in two parallel pathways passing through the anterior optic tubercle in the locust brain [J]. Journal of Neurophysiology, 2005, 94: 3903-3915.
18 HORVATH G, VARJU D. Polarized light in animal vision: polarization patterns in nature [M]. Berlin: Springer, 2004.
19 KRAPP H G. Sensory integration: neuronal filters for polarized light patterns[J]. Current Biology, 2014,24(18):R840-R845.
20 JUNDI B, HOMBERG U. Receptive field properties and intensity response functions of polarization-sensitive neurons of the optic tubercle in gregarious and solitarious locusts[J]. Journal of Neurophysiology, 2012, 108:1695-1710.
21 HOMBERG U, AGNES P. Ultrastructure and orientation of ommatidia in the dorsal rim area of the locust compound eye[J].Arthropod Structure & Development,2002,30(4):271-280.
