rs2231142位点多态性与川东北地区人群原发性痛风的关系
2018-07-02蒋红贾艾敏王丽恒邱亚青玉凤蔡燕杨明辉
蒋红,贾艾敏,王丽恒,邱亚,青玉凤,蔡燕,杨明辉,
(川北医学院附属医院,1.风湿免疫研究所;2.医学研究中心;3.检验科;4.风湿免疫科,四川 南充 637000)
痛风性关节炎是由于嘌呤代谢紊乱,导致尿酸生成过多而诱发关节炎的炎症性疾病,由先天性嘌呤代谢障碍和尿酸排泄减少所致的痛风称为原发性痛风。尽管痛风的发病机制仍然不清楚,但越来越多的证据显示原发性痛风60%与遗传因素有关,且约90%原发性痛风和高尿酸血症是由于尿酸排泄减少所致[1-2]。三磷酸腺苷结合转运蛋白G超家族成员2(ATP-binding cassette,sub-family G,member 2,ABCG2)是一个高容量的尿酸分泌型转运蛋白,主要在近曲小管的管腔膜侧表达,必须形成同型二聚体才能发挥转运尿酸的活性,研究表明ABCG2 蛋白主要参与肾脏尿酸的分泌[3]。其单核苷酸多态性(SNP)rs2231142位点变异引起ABCG2蛋白的141位的谷氨酸(Q)变成赖氨酸(K),尿酸转运活性较野生型明显下降,进而引起血尿酸明显升高,导致痛风发生[3]。
研究表明,ABCG2蛋白rs2231142(A/C)基因多态性与痛风的发生密切相关,且rs2231142多态性的分布具有明显的地域性和人群差异性特征[3-6]。本实验对川东北地区男性痛风患者ABCG2基因rs2231142多态性与痛风的相关性进行研究,为该地区原发性痛风的发病机制研究提供线索。
1 材料与方法
1.1 研究对象
本实验选取2013年1月至2015年12月在川北医学院附属医院风湿血液科诊断为原发性痛风的175例男性患者作为病例组,年龄18~85岁,平均年龄(47.8 ±13.4)岁。纳入标准:原发性痛风诊断标准依据1977年美国风湿病学会诊断标准[7];排除标准:排除近期服用可能导致尿酸升高药物的患者,排除任何可能导致尿酸升高的疾病,排除因嘌呤代谢酶导致的尿酸代谢缺陷[8-9]。同时,选取无高尿酸血症及痛风既往史和家族史,无其它自身免疫性疾病,与病例组年龄、性别、职业特点无明显差异的277例健康体检者作为对照组,年龄 23~78岁,平均年龄(46.5±12.8)岁。
1.2 外周血DNA提取
用EDTA抗凝管抽取175例痛风患者和277例健康对照人群的静脉血3 mL,室温反应30 min,分离血浆,将剩余血液成分按每管500 uL进行分装,做好标记,于-80 ℃保存备用。使用前将冻存的血液于37 ℃水浴溶解,用天根的血液DNA提取试剂盒(离心柱法)提取外周血DNA,按试剂盒说明书进行外周血DNA提取;用Pultton P200+超微量分光光度计检测DNA浓度和质量,OD260/OD280=1.8~2.0表示DNA提取效果较好,可用于Taqman-MGB探针法SNP分型。
1.3 Taqman-MGB探针法检测rs2231142位点多态性
从ABI公司购买rs2231142位点对应的Taqman-MGB探针,实时荧光定量PCR(qRT-PCR)Taqman-MGB探针法检测样本SNP位点基因型及等位基因频率:只检测到VIC信号,表明所测样本基因型为CC型;只检测到FAM信号,表明所测样本基因型为AA型;同时检测到VIC信号和FAM信号,表明基因型为CA型。反应体系5 μL:TaqMan Universal PCR Master Mix (2×)2.50 μL,40×SNP Genotyping Assay 0.25 μL,模板DNA 1 μL;反应条件为:95 ℃ 10 min,92 ℃ 15 s,60 ℃ 1 min,40个循环;扩增在ABI QuantStudio 12K荧光定量PCR仪上进行。
1.4 统计学分析
按照Taqman-MGB探针说明书统计实验结果,hardy-weinberg平衡检测对照组的群体代表性,P>0.5,表明所选样本具有群体代表性;P<0.5,表明所选样本不具有群体代表性。用χ2检验分析病例组与对照组的基因型和等位基因型差异,分析结果用OR值和95%的置信区间(95% CI)表示;P<0.05,表示差异具有显著性;P<0.01,表明差异极显著。SHEsis在线分析软件用于结果统计。
2 结果
2.1 DNA提取质量检测
OD260/OD280=1.8~2.0表示DNA提取质量良好,且用于Taqman-MGB探针法SNP分型效果好。对于OD260/OD280<1.8,说明样品中酶和蛋白含量过高,应将样品用酚、氯仿、异戊醇抽提,再用乙醇沉淀DNA;OD260/OD280>2.0,说明样本中RNA含量过高,可用RNase消化后,用酚、氯仿、异戊醇抽提,再用乙醇沉淀DNA。或者用分装的血液标本进行二次抽提,保证DNA提取质量满足分型要求。
2.2 基因分型检测结果
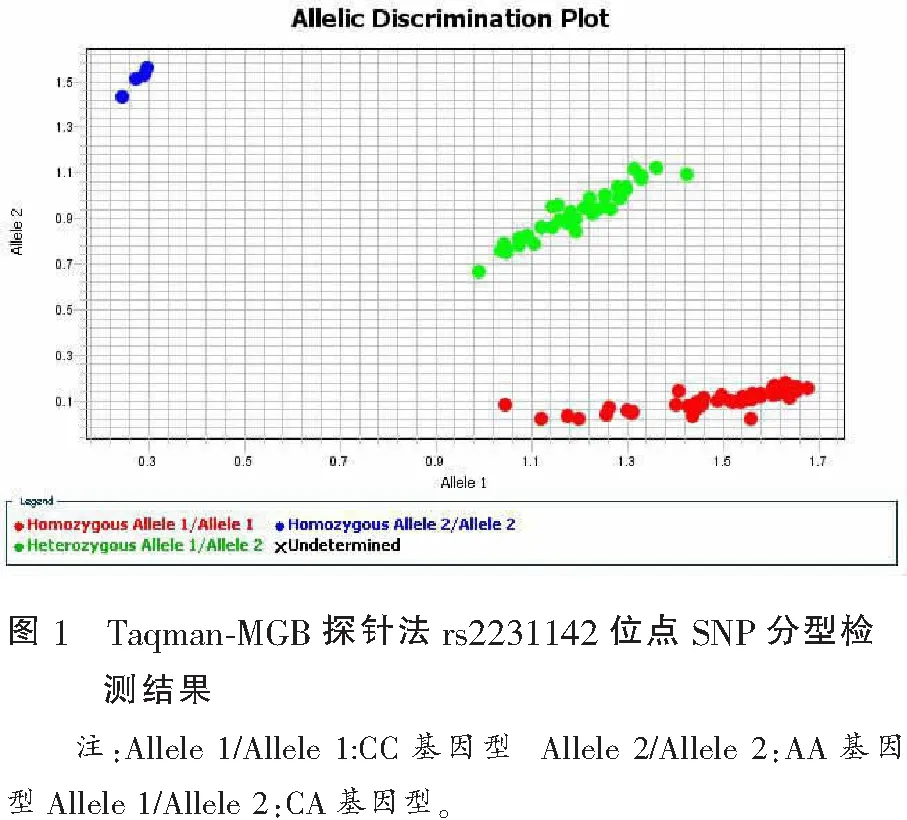
Taqman-MGB探针法检测ABCG2基因rs2231142位点SNP分型,结果如图1所示:红点表示只检测到荧光信号VIC,所测样本基因型为CC型;蓝点表示只检测到荧光型号FAM,所测样本基因型为AA型;绿点表示同时检测到荧光信号VIC和FAM,所测样本基因型为CA型。根据荧光信号分布情况,初步判断突变型AA基因型和A等位基因在原发性痛风患者中的分布频率更高。
2.3 两组基因型和等位基因型频率分布的分析
对175例痛风患者和277例健康人进行SNP分型分析,所得结果进行hardy-weinberg平衡检测,检测结果显示对照组P=0.92,符合hardy-weinberg遗传平衡,具有群体代表性。进一步分析显示(表1),rs2231142位点CC基因型,在痛风组中分布频率为35.4%,对照组中分布频率为46.9%;CA杂合型在痛风组中分布频率为41.7%,对照组为43.0%;AA型在痛风组中分布频率为22.9%,对照组为10.1%;各基因型分布频率在痛风患者和正常人群之间差异极显著(P<0.01)。rs2231142位点等位基因A在痛风患者中的携带频率显著高于正常人群,分别为43.7%和31.6%(P<0.01),OR值为1.682(1.275~2.218)。实验结果提示:rs2231142多态性与川东北地区男性原发性痛风的发病相关,A等位基因和AA基因型为痛风发病的易感因素,C等位基因和CC型为痛风发病的保护机制。

表1 rs2231142位点基因型和等位基因型频率分布比较[n(%)]
3 讨论
2017我国痛风患者已经超过8 000万,且呈现发病率增高,发病年轻化的趋势,痛风已成为我国仅次于糖尿病的第二类代谢性疾病。痛风以男性发病为特征,男性患者占80%以上,发病年龄一般在45岁左右。痛风的发病与患者不健康的饮食习惯(高热量、高脂肪、高嘌呤食物摄入过多)和生活习惯(熬夜、不运动、不节制饮酒等)相关[10],此外,遗传因素是原发性痛风发病的主要原因。痛风不仅引发关节损伤,痛风性关节炎反复发作可形成痛风结节和转变为慢性痛风性关节炎,并导致关节功能障碍和造成骨破坏,晚期还可能导致痛风性肾病及肾功能衰竭,最终引起死亡。痛风不仅累及关节和肾脏,而且可诱发和加重现代流行病如糖尿病、冠心病、高血压及脑血管疾病的的发生和发展,严重影响了患者的生活质量,给家庭和社会带来巨大的精神和经济负担[9-10]。探索痛风的发病机制,发现早期诊断标志和治疗靶点,对痛风的治疗至关重要。
原发性痛风与嘌呤代谢异常和尿酸排泄减少相关 ,肾脏尿酸排泄减少的相关基因包括有机阴离子转运体1(OAT1)基因、尿酸盐阴离子交换器(hURATI)基因、人葡萄糖转运蛋白9(GLUT9)基因和三磷酸腺苷结合转运蛋白G超家族成员2(ABCG2)基因等[11-13]。研究表明,ABCG2主要参与肾脏尿酸的分泌,体外实验证实SNP rs2231142位点变异引起ABCG2蛋白的141位的谷氨酸(Q)变成赖氨酸(K)(Q141K),使ABCG2蛋白多肽链的氨基酸种类和序列发生改变,引起尿酸升高,导致痛风的发生[3]。本实验以川东北地区男性原发性痛风患者为研究对象,通过Taqman-MGB探针法检测ABCG2基因 rs2231142位点多态性与痛风的关系。实验结果表明,痛风组AA基因型和A等位基因的携带率显著高于对照组,A等位基因携带者发病风险是C等位基因携带者的1.682倍,A等位基因是原发性痛风发病的危险因素。据报告,rs2231142位点A等位基因型在不同人群中的分布不同[4,14-17],对川东北地区男性痛风患者的rs2231142位点进行SNP分型分析,对揭示该地区原发性痛风的发病机制具有重要意义,为将来拟进行的靶向药物治疗痛风筛选位点。
[1] 熊安,姚麒,余晶波.高尿酸血症和痛风易感基因单核苷酸多态性研究新进展及应用[J].宁波大学学报(理工版),2016,29(2):116-120.
[2] Dehghan A,Kottgen A,Yang Q,etal.Association of three genetic loci with uric acid concentration and risk of gout: a genome-wide association study[J].Lancet,2008,372(9654):1953-1961.
[3] Woodward OM,Kottgen A,Coresh J,etal.Identification of a urate transporter,ABCG2,with a common functional polymorphism causing gout[J].PNS,2009,106(25):10338-10341.
[4] Wang B,Miao Z,Liu S,etal.Genetic analysis of ABCG2 gene C421A polymorphism with gout disease in Chinese Han male population[J].Hum Genet,2010,127(2):245-246.
[5] 李发贵,楚轶,孟冬梅,等.ABCG2基因第5外显子C421A多态与中国汉族男性原发性痛风的相关性研究[J].中华遗传医学杂志,2011,28(6):683-685.
[6] 邱亚,柳华,周京国,等.三磷酸腺苷结合盒亚家族G超家族第二个成员(ABCG2)基因rs2231142多态性与痛风发病风险的Meta分析[J].中华风湿病学杂志,2015,19(6):384-388.
[7] 中华医学会风湿病学分会.原发性痛风诊断和治疗指南[J].中华风湿病学杂志,2011,15(6):410-413.
[8] 余家会,何霞,张蓓,等.新疆地区维吾尔族和汉族人群ABCG2基因rs2231142位点多态性与高尿酸血症的相关性[J].临床检验杂志,2015,33(2):142-146.
[9] 王婧宇,常宝成.高尿酸血症/痛风流行病学特点及危险因素[J].国际内分泌代谢杂志,2016,36(2):78-81,88.
[10] 关宝生,白雪,王艳秋,等.痛风/高尿酸血症患者生活习惯的危险因素[J].中国老年学杂志,2014,34(2):455-457.
[11] 董鹏,宋慧.痛风发病机制研究进展[J].基础医学与临床,2015,35(12):1695-1699.
[12] Kawamura Y,Matsuo H,Chiba T,etal.Pathogenic GLUT9 mutations causing renal hypouricemia type 2 (RHUC2)[J].Nucleosides Nucleotides Nucleic Acids,2011,30(12):1105-1111.
[13] Anthony,Reginato,David B,etal.The genetics of hyperuricaemia and gout[J].Nat Rev Rheumatol,2012,8(10):610-621.
[14] 王金丹,余玲玲,黄德益,等.ABCG2基因rs2231142位点多态性与浙南地区人群原发性痛风的关系[J].基础医学与临床,2016,36(4):503-507.
[15] Matsuo H,Takada T,Ichida K,etal.Common defect of ABCG2,a high capacity urate exporter cause gout: a function based genetic analysis in a Japanese population[J].Sci Transl Med,2009,1(5):5ra11.
[16] Chang HY,Pan WH,Yeh WT,etal.Hyperuricemia and gout in Taiwan:results from the Nutritional and Health Survey in Taiwan (1993-96)[J].J Rheumatol,2001,28(28):1640-1646.
[17] Miao Z,Li C,Chen Y,etal.Dietary and lifestyle changes associated with high prevalence of hyperuricemia and gout in the Shandong coastal cities of Eastern China[J].J Rheumatol,2008,35(9):1859-1864.
