扎龙湿地不同生境芦苇种群根茎数量特征及动态
2018-06-23焦德志姜秋旭闫秋月杨允菲
焦德志,姜秋旭,曹 瑞,闫秋月,杨允菲
1 齐齐哈尔大学生命科学与农林学院,齐齐哈尔 161006 2 东北师范大学草地科学研究所,植被生态科学教育部重点实验室,长春 130024
植物的根茎不仅是营养物质贮存器官,也是重要的繁殖器官[1]。根茎不仅具有营养繁殖和扩展种群空间的功能[2],同时,也是无性系植物不同构件间进行生理整合的通道[3]。
根茎宿存于土壤中,其繁殖和生活均要消耗大量养分,不同植物根茎的寿命取决于遗传特性[4],有的植物根茎是短寿型的,如光稃茅香[5]和牛鞭草[6]的根茎只能活1—2年,有的植物根茎是长寿型的,如羊草[7]的根茎能活3年以上,芦苇[8]的根茎甚至可存活10年以上。但是,即使是长寿型根茎,也不是每条根茎都同等长寿,单个根茎的寿命不仅与养分的输出消耗有关,也与养分能否再输入贮藏密切相关[9-10]。
根茎型无性系植物能够通过地下根茎的变长或变短来完成种群的迁徙和对生境的选择,将分株安置在远离母株的生境中,避免空间和养分的种内竞争[11];无性系构件的生长具有觅养特性和种群的扩展行为[12]。在根茎的数量特征中,根茎芽是种群的营养繁殖潜在种群,根茎的长度蕴含着生长和空间扩展等信息,根茎的生物量蕴含着营养物质的储存状况[1]。因此,开展异质生境根茎的数量特征及时空动态研究,具有重要的理论与实践意义。通过对扎龙湿地保护区内4个异质生境芦苇种群5个生育期的调查测定,比较根茎长度、生物量和干物质贮量的生境间差异,以及随生育期进程各数量特征的变化规律,籍以揭示芦苇种群根茎生长及根茎物质消耗和贮藏的生物学特性和生态学意义,为芦苇种群水平趋异适应机理的深入研究提供科学依据,也对扎龙湿地保护区的恢复和保护以及科学管理提供理论参考。
1 研究地区和研究方法
1.1 研究区概况
本项研究是在松嫩平原北部,扎龙国家级自然保护区(齐齐哈尔东南23 km)。其位于46°52′—47°32′N,123°47′—124°37′E,该地区属于中温带大陆性季风气候,冬寒漫长,春干风大,夏热多雨,秋凉霜早,年平均气温为3.2 ℃,最冷月平均气温-19.4 ℃,最热月平均气温22.9℃,年日照时数2700—3000 h,年辐射总量是2052—2140 s/cm2,积温2600—3000 ℃,平均地温4.9 ℃,无霜期128 d[13-14]。
1.2 样地设置
在扎龙国家级自然保护区核心区,以土壤水分条件,从旱生、湿生、水生及盐碱化土壤,共设置4个生境样地:
1)旱生生境(H1):土壤为草甸土,全年无积水或7—8月份雨季短时积水,经常形成羊草+芦苇群落,微地形中心也可形成面积不等的单优种芦苇群落斑块[15]。
2)湿生生境(H2):土壤为沼泽土,平时无积水或少量积水,7—8月雨季来临以后逐渐积水,积水深度一般情况低于30 cm[16]。
3)水生生境(H3):土壤为腐殖沼泽土,形成湿地内面积最大的芦苇群落,常年积水,水深变化为10—100 cm[16]。
4)盐碱生境(H4):土壤为盐碱土,全年无积水,土壤表层已完全丧失,具有明显盐碱斑,有的碱斑可形成单优势种芦苇居群[15]。
样地多年来一直冬季收割,造纸或民用。另外,4个生境间距离均在5 km内,光照和温度等环境条件可视为一致。
1.3 研究方法

图1 不同生长期异质生境芦苇种群根茎长度比较 Fig.1 Comparison of rhizome length in different growing period of Phragmites australis populations in heterogeneous habitats H1:旱生生境 Xeric habitats; H2:湿生生境 Wet habitats; H3:水生生境 Aquatic habitats; H4:盐碱生境 Saline-alkali habitats;不同小写字母表示生境间差异显著(P<0.05)
2013年6—10月份,在芦苇的营养生长初期(6月15日)、营养生长旺盛期(7月10日)、生殖生长初期(8月15日)、生殖生长旺盛期(9月12日)、休眠期(10月15日)取样。地上分株采取单位面积(1 m×1 m)刈割取样,地下根茎采取单位体积(1 m×1 m×1 m)挖土取样,3次重复。芦苇根茎集中分布在1 m土层以内,在4个样地内分别挖取长宽深为1 m的土体,捡取土体中的所有根茎,分装在不同的塑料袋里,带回实验室。逐样方测量每条根茎的长度,计算单位土体根茎的累积长度后,80 ℃下烘至恒量,称其生物量。用土钻对每一样点土壤取样,深度为1 m,3次重复,把相同样点的土壤样品充分混匀后过筛备用。烘干法测土壤含水量;电位法测pH值;重铬酸钾加热法测有机质含量;碱解扩散法测速效氮含量;钼锑抗比色法测速效磷含量[17-18]。
1.4 数据处理
根茎长度和生物量以实际观测统计值表示。根茎干物质贮量以每100 cm单位长度根茎生物量表示。用M 反映样本的平均数量,SD 反应样本的绝对变异程度。用Excel 2007绘制图表,SPSS 19.0 软件对数据进行分析处理。对不同月份和不同生境间差异进行单因素方差分析(One-way ANOVA,ɑ=0.05)。对根茎长度、生物量、干物质贮量与返青后的实际生长时间(以5月10日返青计)分别用线性、指数、二次函数进行回归,选取相关性最高的拟合方程作为定量刻画模型,方程的拟合优度以R2检验,方程显著性采用F检验。重复取样获得的根茎数量特征数据和土壤因子间,分株密度与根茎长度间分别进行Pearson相关分析,其中土壤含水量相关分析中均不包括水生生境(H3),用相关系数反映土壤因子对根茎数量特征的影响及根茎长度与分株密度间关系,正值为正向驱动,负值为负向驱动。
2 结果分析
2.1 不同生境芦苇种群根茎长度动态
根茎的长度是根茎生长和空间扩展的重要标志。在扎龙湿地保护区,芦苇一般5月10左右返青,4个生境芦苇种群根茎长度动态如图1和图2所示。各月份根茎长度均以H2最大,为13754—17446 cm/m3之间,显著(P<0.05)高于其他3个生境,依次H1为11639—14330 cm/m3之间,H3为9676—13027 cm/m3之间,H4最小,为7898—9877 cm/m3之间,显著低于其他3个生境,生境之间的差异均达到了显著水平(图1)。根茎长度从6月份开始逐渐增加,其中6—8月份缓慢增加,8—10月份显著增加,后期是前期的3.5—10.3倍(图2)。由此反映了各生境芦苇种群根茎长度的差异以及差异序位基本稳定,在生长季中后期均有一个持续时间较长的新根茎补充和生长时期,以此实现种群的空间扩展,也为种群的营养繁殖储备了大量的繁殖芽。
经回归分析,4个生境芦苇种群根茎长度与返青后的实际生长时间(以5月10日返青计)之间均较好地符合直线函数关系(图2),方程的拟合优度(R2)在0.804—0.920之间,并均达到了P<0.05的显著水平。

图2 异质生境芦苇种群根茎长度季节动态Fig.2 Seasonal changes of rhizome length of Phragmites australis populations in heterogeneous habitats
2.2 不同生境芦苇种群根茎生物量动态

图3 不同生长期异质生境芦苇种群根茎生物量比较 Fig.3 Comparison of rhizome biomass in different growing period of Phragmites australis populations in heterogeneous habitats
根茎生物量是根茎养分贮量的主要表征,也是无性系植物进行营养繁殖的物质基础。4个生境芦苇种群根茎生物量动态如图3和图4所示。各月份根茎生物量均以H2最大,为1063.6—1379.6 g/m3之间,依次是H1为939.3—1247.0 g/m3之间,H3为624.6—952.2 g/m3之间,H4最小,为587.0—787.3 g/m3之间,其中6—8月份H3和H4间,7—9月份的H1和H2间差异均没达到显著水平(P>0.05)(图3)。根茎生物量6—8月份逐渐减少,8—10月份逐渐增加,生长季末期的10月份最大,并均显著地高于其他月份(图4)。由此反映出各生境芦苇种群根茎生物量生境间差异序位均相对稳定,但生长季后期差异更为明显,在生长季前中期均有一个持续时间较长的根茎养分消耗过程,中后期均又有一个持续时间较长的根茎养分储藏过程。无性系植物在生长季前中期,将根茎储藏的物质用于新根茎和芽的形成与生长,而中后期将光合产物转移分配给地下储藏与营养繁殖器官,为翌年根茎芽的萌发做好物质储备。
经回归分析,4个生境芦苇种群根茎生物量与返青后的实际生长时间(以5月10日返青计)之间均较好地符合二次曲线函数关系(图4),方程的拟合优度(R2)在0.984—0.997之间,并均达到了P<0.05的显著水平。
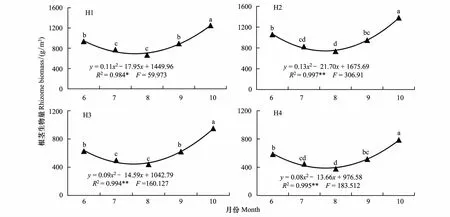
图4 异质生境芦苇种群根茎生物量季节动态Fig.4 Seasonal changes of rhizome biomass of Phragmites australis populations in heterogeneous habitats
2.3 不同生境芦苇种群根茎干物质贮量动态
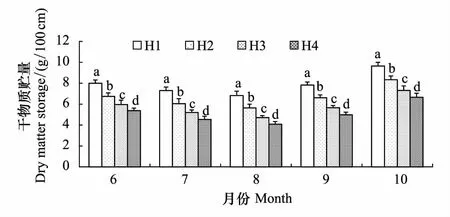
图5 不同生长期异质生境芦苇种群根茎干物质贮量比较 Fig.5 Comparison of rhizome dry matter storage in different growing period of Phragmites australis populations in heterogeneous habitats
根茎干物质贮量(单位长度根茎生物量)可以衡量根茎的养分贮藏和消耗,也是决定根茎寿命长短的直接证据。4个生境芦苇种群根茎干物质贮量动态如图5和图6所示。各月份根茎干物质贮量均以H1最大,为8.0—9.6 g/100 cm之间,依次是H2为6.7—8.3 g/100 cm之间,H3为5.9—7.3 g/100 cm之间,H4最小,为5.4—6.6 g/100 cm之间,4个生境之间的差异均达到显著水平(P<0.05)(图5)。根茎干物质贮量6—8月份逐渐减少,8—10月份逐渐增加,生长季末期的10月份最大,并均显著地高于其他月份(图6)。由此反映出4个异质生境芦苇种群干物质贮量在整个生长期内差异性及其差异序位均相对稳定,在生长季中后期对根茎养分的贮藏均超过生长季前期的消耗,一方面反映了从生长季末期到6月份取样时,芦苇根茎贮藏的养分发生了消耗,另一方面从物质补偿的活力看,芦苇种群对地下根茎存在明显的养分“超补偿性”贮藏现象,也是芦苇根茎维持长寿的最重要证据。
经回归分析,4个生境芦苇种群根茎干物质贮量与返青后的实际生长时间(5月10日返青计)之间均较好地符合二次曲线函数关系(图6),方程的拟合优度(R2)在0.990—0.993之间,并均达到了P<0.05的显著水平。

图6 异质生境芦苇种群根茎干物质贮量季节动态Fig.6 Seasonal changes of rhizome dry matter storage of Phragmites australis populations in heterogeneous habitats
2.4 芦苇种群根茎长度与分株密度的关系
在扎龙湿地保护区,芦苇种群分株主要由根茎芽萌发形成,根茎生产具有重要的营养繁殖意义。对4个生境芦苇种群分株密度与根茎长度进行相关性分析,结果表明分株密度(x)与根茎长度(y)极显著(P<0.01)正相关,最优拟合方程为直线关系改成y= 5355.5+19.8x(R2=0.45,P=0.001,n=20)。拟合方程中参数b值为19.8,其生物学含义为分株密度每增加1株/m2,根茎需要至少延长19.8 cm/m3。由此可见,单位土体芦苇种群根茎越长,单位面积分株数量可能会越大。
2.5 芦苇种群根茎数量特征与土壤因子的关系
对4个生境芦苇群落的立地土壤同步取样并进行理化性质测定。经统计分析和多重比较,4个生境5个土壤因子存在较大差异(表1)。H3因常年积水,土壤含水量处于饱和状态,其他3个生境各月份土壤含水量均以H2最高,均以盐碱生境H4最低,各生境间差异均达到显著水平(P<0.05),H1、H2和H4土壤含水量因季节性降雨波动较大,均以6月份最低,9月份最高;各月份土壤pH值均以H4最高,均显著地高于其他3个生境,以H3最低,其中和H2差异均没达到显著水平(P>0.05),各生境均以9月份最低,10月份最高;各月份土壤有机质、速效氮含量均以H2最高, H4最低,各生境间的差异均达到显著水平;各月份土壤速效磷含量均以H4最高,H1最低,各生境间的差异均达到显著水平,生长期内土壤有机质、速效氮、速效磷含量呈逐渐降低的趋势。因此,4个生境芦苇群落的5个土壤因子既存在比较稳定的空间差异,同时,也表现出明显的季节性波动。

表1 不同生境土壤理化特征
H1:旱生生境 Xeric habitats; H2:湿生生境 Wet habitats; H3:水生生境 Aquatic habitats; H4:盐碱生境 Saline-alkali habitats;表示生境间差异显著(P<0.05)
对4个生境土壤因子与芦苇种群根茎数量特征进行相关性分析(表2)。结果表明, 5个土壤因子对芦苇种群根茎数量特征的驱动作用不同,土壤含水量、pH、有机质和速效氮是芦苇种群根茎长度、生物量的主要影响因子,土壤pH、速效磷是根茎干物质贮量的主要影响因子。土壤含水量、有机质、速效氮为正向驱动,pH、速效磷为负向驱动,土壤含水量、pH的驱动作用更突出。

表2 芦苇种群根茎数量特征与土壤因子之间的相关系数(n=12)
*P<0.05; **P<0.01
3 讨论
3.1 根茎生长和物质生产的生物学与适应意义
根茎型无性系植物在不同生境中表现出特有的适应方式,包括形态可塑性、觅食行为、生理整合以及营养繁殖适合度调节等等,根茎型无性系植物的研究不断得到广泛的关注[19]。研究表明,植物死亡有至少95%是发生在种子阶段[20],但植物的根茎具有极强的繁殖能力和存活率[21-22],由根茎芽萌发形成的分蘖苗比种子萌发形成的实生苗具有更强的生存能力[23],在天然群落的种群更新中远远超过种子更新[24],在此种意义上,根茎生长具有种群更新与适应意义。根茎不仅是无性系植物的主要营养繁殖构件,而且是养分的主要贮藏器官,单个根茎的寿命不仅与养分的输出消耗有关,也与养分能否再输入贮藏密切相关[9-10],在此种意义上,根茎生产具有养分消耗和储存等生物学和生态学意义[4]。
研究发现,在扎龙湿地保护区,4个生境芦苇种群根茎在土体中呈纵横交错分布。整个生长期内,根茎长度均呈逐渐增加的趋势,新根茎形成与伸长生长的同时,老根茎也存在陆续死亡的现象。因此,新根茎的产生不仅可以补充因衰老而死亡的老根茎对潜在种群造成的损失,其水平方向延伸,还可以使新的分株避免与亲代根茎形成的分株对生存空间和资源的直接竞争,更有利于种群占据或扩展水平方向的生态位空间[25]。种群根茎生物量和干物质贮量在整个生育期内呈先减少后增加的“V”形趋势,从营养生长初期的6月份逐渐减少,到生殖生长初期的8月份最小,然后又逐渐增加,生长季末期的10月份最大,各生境芦苇种群在生长季前期均有一个持续时间较长的根茎养分消耗过程,后期又均有一个持续时间较长的根茎养分储藏过程,后期对根茎养分的贮藏均超过前期的消耗,是芦苇根茎能够维持长寿的物质基础,也是其根茎具有较强活力的直接证据[2,26]。
三江平原芦苇种群生产力在整个生长季内,其地下生物量增长和下降趋势非常显著[27]。长江口淡水潮湖芦苇种群地下部分根茎的生物量呈单峰变化[28],最小值出现在5月份,最大值出现在9月份,其地上和地下生物量可分为此消彼长、共同增长和共同降低3个阶段。由于受研究地区不同气候条件的影响,芦苇种群的生育期在时间(月份)上会有所提前或滞后,地下生物量峰值出现的时间自然也表现出早晚差异,但各研究区域内根茎生物量的动态变化却具有高度一致的生育期节律,这种节律性同样应该是由遗传因素所决定的。在松嫩平原大多数禾草的根茎一般最多存活2—4年,硬拂子茅和假苇拂子茅根茎最多可存活3年[29- 30],羊草根茎最多可存活4年[6],而芦苇根茎一般可存活6年,个别可存活7—9年,乃至更长时间[7]。对上述根茎存活2—4年的禾草的研究表明,随着生育期的进程,根茎干物质贮量均呈下降的趋势,表明整个生育期根茎养分一直处于消耗状态,其养分未给予再输入,也是根茎生活力衰退的重要标志。当然,干物质贮量只是为理解根茎营养物质的消耗和补偿提供了宏观证据,对于如何解释芦苇根茎的长寿性和其他禾草根茎的短寿性现象,需要进一步开展不同种群的生存与发展策略及其生理生化调节机理的研究[7],另外,对于芦苇长寿的分子机制也有待进一步研究。
3.2 土壤因子对芦苇种群根茎数量特征的驱动影响
植物生长繁殖的必需资源及植物所处的环境条件在很小的尺度上也存在着异质性[10],环境异质性作为一种选择压力,使得植物可以形成有效获取必需资源的生态适应对策[31-32]。无性系植物的根状茎具有较大的储藏功能[33-34],克隆植物能够通过克隆器官适时存储与释放资源,从而缓解资源异质性所带来的压力[11]。根茎处于微生境的土壤中,既具有根的空间位置又承担着茎的生理功能,成为有机体与无机环境之间信息整合的纽带和桥梁[35]。特殊的结构和功能使根茎成为深入研究植物与生境之间协同关系的最佳模式之一,研究根茎与环境之间的相互作用,更有助于了解植物的进化历史和未来趋势[35]。
研究表明,4个生境芦苇种群的根茎长度和根茎生物量均以湿生生境最大,依次为旱生生境和水生生境,盐碱生境最小,根茎干物质贮量以旱生生境最大,依次为湿生生境和水生生境,盐碱生境最小,在整个生长期内生境间差异以及差异序位均相对稳定,表现出明显的环境效应,生境间的差异是多种土壤因子综合协同作用的结果,并非某种因子单独作用引起的,其中土壤含水量和pH的驱动作用更明显。
以往对根茎生长和生物量的研究也证实这一点。羊草根茎在58.43%的水分处理下直径最高,在20.87%中度水分胁迫下有利于节间距增长,重度和轻度水分胁迫下茎节间距增长缓慢[36],当盐碱胁迫上升至pH=9.84,电导率0.88 s/cm,羊草根茎长度、节密度和节间距明显减小(P<0.05)[37]。假苇拂子茅通过增加种群的地下部分生长,增强整个种群的水分获取与储存能力,提高整个分株种群应对水分异质性的能力[37]。同样,多年生湿地植物的根茎由于环境条件的变化也表现出可塑性,在松嫩平原水淹后的单优芦苇群落根茎长度和生物量均高于很少积水或积水时间短的其他样地[1],深水中的芦苇根茎较短,在基质中埋深也较浅,可能是为了缩短氧气的运输距离[38]。而对香蒲(TyphaorientalisPresl.)的研究结果则表明,在稳定水体中香蒲的根茎长度随水位升高呈增加趋势[39]。可见,同一环境因子对不同的物种也可能会产生不同的影响。
参考文献(References):
[1] 杨允菲, 郎惠卿. 不同生态条件下芦苇无性系种群调节分析. 草业学报, 1998, 7(2): 1- 9.
[2] Asaeda T, Manatunge J, Roberts J, Hai D N. Seasonal dynamics of resource translocation between the aboveground organs and age-specific rhizome segments ofPhragmitesaustralis. Environmental and Experimental Botany, 2006, 57(1/2): 9- 18.
[4] 杨允菲, 张宝田, 田尚衣. 松嫩平原旱地生境芦苇种群不同龄级根茎的干物质贮藏及水溶糖含量. 应用生态学报, 2008, 19(9): 1905- 1910.
[5] 杨允菲, 李建东, 郑慧莹. 松嫩平原光稃茅香无性系种群的营养繁殖特征. 应用生态学报, 1997, 8(6): 571- 574.
[6] 杨允菲, 李建东, 郑慧莹. 松嫩平原牛鞭草无性系种群的营养繁殖策略. 草业学报1997, 6(2): 36- 40.
[7] 李海燕, 杨允菲. 松嫩平原水淹恢复演替过程中羊草无性系种群构件的年龄结构. 生态学报, 2004, 24(10): 2171- 2177.
[8] 杨允菲, 魏春雁, 张宝田, 刘宝. 松嫩平原碱化草甸旱地生境芦苇种群的芽流和芽库动态. 应用生态学报, 2005, 16(5): 854- 858.
[9] Hong M G, Kim J G. Role and effects of winter buds and rhizome morphology on the survival and growth of common reed(Phragmitesaustralis). Paddy and Water Environment, 2014, 12(1): 203- 209.
[10] 轩弋淳, 包国章, 王鑫. 松嫩平原羊草根茎分枝格局对水分梯度的响应. 科学技术与工程, 2014, 14(17): 174- 177.
[11] 董鸣. 资源异质性环境中的植物克隆生长: 觅食行为. 植物学报, 1996, 38(10): 828- 835.
[12] 董鸣. 克隆植物生态学. 北京: 科学出版社, 2011.
[13] 中华人民共和国林业部. 扎龙国家级自然保护区管理计划. 北京: 中国林业出版社, 1997: 1- 5.
[14] 赵魁义. 中国沼泽志. 北京: 科学出版社, 1999: 231- 239.
[15] 焦德志, 么璐, 黄曌月, 杨允菲. 东北草地异质生境芦苇芽种群动态. 应用生态学报, 2015, 26(2): 404- 410.
[16] 焦德志, 黄曌月, 周婵, 杨允菲. 扎龙湿地异质生境芦苇种群根茎动态及年龄结构. 生态学杂志, 2016, 35(4): 888- 895.
[17] 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000.
[18] 骆东奇, 白洁, 谢德体. 论土壤肥力评价指标和方法. 土壤与环境, 2002, 11(2): 202- 205.
[19] 汤俊兵, 肖燕, 安树青. 根茎克隆植物生态学研究进展. 生态学报, 2010, 30(11): 3028- 3036.
[20] 班勇. 植物生活史对策的进化. 生态学杂志, 1995, 14(3): 33- 39.
[21] De Kroon H, Knops J. Habitat exploration through morphological plasticity in two chalk grassland perennials. Oikos, 1990, 59(1): 39- 49.
[22] Dong M, de Kroon H. Plasticity in morphology and biomass allocation inCynodondactylon, a grass species forming stolons and rhizomes. Oikos, 1994, 70(1): 90- 106.
[23] Harris D, Davy A J. Regenerative potential ofElymusfarctusfrom rhizome fragments and seed. Journal of Ecology, 1986, 74(4): 1057- 1067.
[24] 刘凤红, 叶学华, 于飞海, 董鸣. 毛乌素沙地游击型克隆半灌木羊柴对局部沙埋的反应. 植物生态学报, 2006, 30(2): 278- 285.
[25] 梁士楚, 李久林, 程仕泽. 贵州青岩油杉种群年龄结构和动态的研究. 应用生态学报, 2002, 13(1): 21- 26.
[26] Karunaratne S, Asaeda T, Yutani K. Growth performance ofPhragmitesaustralisin Japan: influence of geographic gradient. Environmental and Experimental Botany, 2003, 50(1): 51- 66.
[27] 张友民, 杨允菲, 王立军. 三江平原沼泽湿地芦苇种群生产与分配的季节动态. 中国草地学报, 2006, 28(4): 1- 5.
[28] 张佳蕊, 张海燕, 陆健健. 长江口淡水潮滩芦苇地上与地下部分月生物量变化比较研究. 湿地科学, 2013, 11(1): 7- 12.
[29] 杨允菲, 张宝田, 李建东. 松嫩平原硬拂子茅无性系种群营养繁殖的数量特征. 草业学报, 1998, 7(4): 7- 12.
[30] 杨允菲, 郑慧莹. 松嫩平原假苇拂子茅无性系种群的年龄结构. 草业学报, 2000, 9(3): 8- 13.
[31] 董鸣. 异质性生境中的植物克隆生长: 风险分摊. 植物生态学报, 1996, 20(6): 543- 548.
[32] Dong M, During H J, Werger M J A. Root and shoot plasticity of the stoloniferous herbAjugareptansL. planted in a heterogeneous environment. Flora-Morphology, Distribution, Functional Ecology of Plants, 2002, 197(1): 37- 46.
[33] Dong B C, Yu G L, Guo W, Zhang M X, Dong M, Yu F H. How internode length, position and presence of leaves affect survival and growth ofAlternantheraphiloxeroidesafter fragmentation. Evolutionary Ecology, 2010, 24(6): 1447- 1461.
[34] Dong B C, Zhang M X, Alpert P, Lei G C, Yu F H. Effects of orientation on survival and growth of small fragments of the invasive, clonal plantAlternantheraphiloxeroides. PLoS One, 2012, 5(10): e13631.
[35] 邹元春, 吕宪国, 姜明. 湿地克隆植物根茎对变境适应的表型可塑性. 湿地科学, 2007, 5(4): 305- 310.
[36] 刘滨硕, 康春莉, 李忠民, 蒋丽群, 周杭, 包国章. 不同盐碱梯度生境下羊草根茎生长的研究. 东北师大学报: 自然科学版, 2013, 45(3): 110- 114.
[37] 叶学华, 胡宇坤, 刘志兰, 高树琴, 董鸣. 水分异质性影响两种根茎型克隆植物赖草和假苇拂子茅的水分存储能力. 植物生态学报, 2013, 37(5): 427- 435.
[38] Weisner S E B, Strand J A. Rhizome architecture inPhragmitesaustralisin relation to water depth: implications for within-plant oxygen transport distances. Folia Geobotanica, 1996, 31(1): 91- 97.
[39] White S D, Ganf G G. The influence of convective flow on rhizome length inTyphadomingensisover a water depth gradient. Aquatic Botany, 1998, 62(1): 57- 70.
