小兴安岭森林地表鞘翅目成虫对不同干扰梯度的响应
2018-06-22程赛赛倪娟平高梅香李景科
程赛赛,倪娟平,高梅香,*,李景科
1 哈尔滨师范大学地理科学学院,哈尔滨 150025 2 黑龙江省普通高等学校地理环境遥感监测重点实验室,哈尔滨 150025
人类活动通过作用于生物生存环境而影响物种动态,从而影响着生物多样性的变化[1-2]。随着人为活动和自然干扰的增多,对不同强度干扰下生物多样性的研究更具有现实意义。干扰理论是生态学的重要组成部分,目前研究较多的则是“中度干扰假说”[3- 4],该假说认为中等程度的干扰能够维持高的生物多样性,虽然这一假说得到许多学者的验证[5-7],但对于森林生态系统中地表土壤动物群落是否支持这一假说有待探究。国内学者对于土壤动物对人为干扰强度响应机制的研究开展较少,且只见于草地、耕地和湿地生态系统[8- 10]。张智英等[11]在云南石林喀斯特景区对不同生境大型土壤动物的多样性进行研究,表明受人为干扰较严重的灌木丛和草地,无论土壤动物的类群数、密度、生物量还是多样性都较低。吴东辉、陈鹏等[12]对长春市不同土地利用方式下大型土壤动物进行研究表明,土壤动物多样性随着土地利用方式和人为干扰程度的不同而存在差异,受农业生产活动和旅游干扰较轻的土地利用方式下土壤动物个体数、类群数和多样性相对较高。在森林生态系统中,地表土壤动物对不同干扰强度响应特征的研究却鲜有报道。
凉水自然保护区具有原始林-次生林-人工林完整的干扰梯度类型,为探索森林生态系统经人为干扰或经营后土壤动物多样性和时空变异特征提供了良好的研究平台[13]。本实验在小兴安岭凉水国家级自然保护区,选择代表不同干扰梯度的6种林型(轻度、中度和重度),于2015年7、8、10月采用陷阱法调查地表鞘翅目成虫群落,尝试性回答以下问题:(1)6个林型地表鞘翅目成虫个体数量、物种组成和群落多样性是否存在显著差异和季节变化;(2)鞘翅目成虫个体数量、物种组成是否随干扰强度变化表现为梯度性规律;(3)凉水地区鞘翅目成虫群落多样性是否符合中度干扰假说。通过本实验的分析揭示凉水森林生态系统地表鞘翅目成虫对不同干扰梯度的响应特征,为地表生物多样性保护和森林生态系统管理提供数据支撑。
1 研究区概况与研究方法
1.1 自然概况
研究区位于黑龙江省伊春市带岭区凉水自然保护区内(47°10′50″N,128 °53′20″E),地处小兴安岭山脉南坡达里带岭支脉的东坡,海拔高度在280—707m之间,为典型的低山丘陵地貌。该区地处欧亚大陆东缘,属温带大陆性夏雨季风气候,春季迟缓,多大风,降水少;夏季短,多雨水,降雨占全年60%以上;秋季降温快,多出现早霜;冬季长且寒冷干燥。地带性土壤为暗棕壤,非地带性土壤为草甸土、沼泽土和泥炭土。该区具有古老的区系和群落发生的历史,保存了较完整而典型的地带性顶极群落类型及自然景观,境内森林覆盖率达96%[13-14]。
区内植被群落复杂多样,地带性植被是以红松为主的温带针阔混交林,具有原始林-次生林-人工林完整干扰梯度类型的森林生态系统。选取原始阔叶红松林(以KY表示)、谷地云冷杉林(YL)、阔叶红松择伐林(ZF)、次生白桦林(BH)、落叶松人工林(RL)和红松人工林(RHS)6种林型作为实验样地,其中KY和YL为轻度干扰林型,是受干扰程度较轻的原始林,林龄均在300a以上;ZF和BH为中度干扰林型,是经过人为干扰后经营恢复良好的次生林,ZF林型历史约200a,BH林龄为62a;RL和RHS为重度干扰林型,是由1954年阔叶红松林皆伐后人工造林而成,成林时间均为63a。关于实验样地立地状况和树种组成特征,详见参考文献[13]。
1.2 样地设置与样品采集
在KY、YL、ZF、BH、RL和RHS内分别设置3个20m×30m的重复样方,3个样方随机分布且彼此间隔50m以上。采用陷阱法采集地表土壤动物,在每个样方的4个顶点中随机选择3个顶点(以白色PVC管作为地标),在以地标为中心的30cm半径圆内选取一个采样点。在每个采样点用土钻(内径7cm)挖取一个深15cm的柱状土坑,将诱捕杯(内径7cm,高度14cm)置于土坑内使杯口与地面齐平,内置约占诱捕杯容积2/3的饱和NaCl溶液,杯口上方约10cm处支起一个一次性餐盘防止凋落物和雨水等杂物进入,陷阱放置野外7天7夜后取回。分别于2015年7月、8月和10月开展调查,共采集样品162个(3个采样点×3个样方×6种林型×3次调查)。手捡法分拣地表鞘翅目成虫,并置于95%医用酒精中保存,鞘翅目成虫在体视显微镜(Motic SMZ168)下观察,参照《原色中国东北土壤甲虫图鉴——步行虫类》[15]、《原色中国东北土壤甲虫图鉴——隐翅虫类拟步甲类》[16]、《中国东北的葬甲科研究》[17]、《中国土壤动物检索图鉴》[18]将其鉴定到种。
1.3 数据处理
1.3.1 多度等级和Raunkiaer频度指数的划分
个体数占总数10.0%以上者为优势物种,个体数占总数1.0%—10.0%者为常见物种,个体数占总数1.0%以下者为稀有物种。依据每个物种出现的样点数与总样点数的比值计算Raunkiaer频度指数[19],划分标准为:0—20%为A级,>20%—40%为B级,>40%—60%为C级,>60%—80%为D级,>80%—100%为E级。
1.3.2 多样性指数
Shannon-wiener指数:
(1)
Pielou均匀度指数:
J=H′/lnS
(2)
Simpson指数:
(3)
式中,S为样地中的物种数,Pi为样地中第i物种个体数占总个体数的比例。
密度-类群指数:
(4)
式中,D为第i物种个体数,Dimax为各群落中第i物种的最大个体数,g为群落中的物种数,G为各群落所包含的总物种数,Ci/C为相对次数,即在C个群落中第i物种出现的比率。
1.3.3 群落相似性
选用Jaccard相似性系数q和Gower系数Sg度量群落间的相似程度。
Jaccard相似性系数:
(5)
式中,a为A样地全部物种数,b为B样地全部物种数,c为A、B两样地共有的物种数,q值在0.75—1.00为极相似;0.50—0.74为中等相似;0.25—0.49为中等不相似;0.00—0.24为极不相似。
Gower系数:
(6)
式中,n为两个群落相比较的物种数,Xi为第i物种的个体数,j和k代表两个不同的群落,Ri是第i物种在j和k群落的个体总数。系数最大值为1最小值为0。
1.3.4 方差分析和聚类分析
采用Q-Q Plot对每次调查的数据进行正态性检验,不符合正态分布的数据进行平方根转换使之符合或近似符合正态分布;采用三因素方差分析对干扰梯度、林型和月份进行主效应及交互效应检验,并用LSD多重比较对比不同干扰梯度之间、不同林型之间和不同月份之间的个体数、物种数及多样性指数差异显著性;采用单因素方差分析对比相同月份不同林型之间的个体数、物种数及多样性的差异性和对比同一林型不同月份间个体数、物种数及多样性的差异性,采用LSD法进行组间多重比较;依据个体数和物种数运用平方Euclidean聚类分析对6种林型进行聚类,揭示6种林型之间的相似程度。
数据处理工作均在软件SPSS 21.0、Origin8.6和Excel 2007上完成。
2 结果与分析
2.1 地表鞘翅目成虫群落物种组成及数量特征
所有调查共捕获地表鞘翅目成虫879只,分属9科44物种。总体看,步甲科(Carabidae)、葬甲科(Silphidae)和隐翅虫科(Staphylinidae)为优势科(个体数分别占总数的39.81%、35.16%和23.54%),其他6科均为稀有科(个体数占总数的1.94%)。优势种共有3种,分别为葬甲科的Phosphugaatrata、Nicrophorustenuipes和步甲科的Aulonocarabuscanaliculatus(个体数分别占总数的21.96%、10.69%和14.56%);常见种共有12种,个体数占总数的42.43%;优势种和常见种的个体数占总捕获量的80.65%,是该区地表鞘翅目成虫的主要组成部分;其余29种均为稀有物种,物种数高达总物种数的65.91%,但个体数仅占总捕获量的10.35%。
就空间分布特征来看,优势种和常见种分布广泛,3种优势种在不同林型或干扰梯度中均有分布。常见种的分布差异较大,但其中50%的常见种在6个林型中都有分布。稀有种不仅个体数量少,且分布的比较狭窄,几乎仅分布于1—3个群落中,群落中多数物种空间分布的广狭与个体数量的多寡具有明显的一致性。所有调查中,属于Raunkiaer频度A级的物种数最多,表明绝大多数物种在样地中出现的频度较低;7月和8月Raunkiaer频度基本表现为A>B>C>D>E,10月份物种仅有A级14种和B级1种,符合Raunkiaer频度定律,即群落中低频度物种数目高于频度较高物种的数目[19](表1)。
所有调查中,6种林型地表鞘翅目成虫数量从大到小依次为:RL(232只)>KY(150只)>ZF(144只)>BH(132只)>RHS(120只)>YL(101只),RL林型个体数量与其他5种林型的差异均达到显著水平(P<0.05),其他5种林型个体数量无显著差异;物种数从大到小依次为:BH(23种)>RL(22种)>KY(21种)>ZF(20种)=YL(20种)>RHS(17种),6个林型的物种数均无显著差异。轻度、中度和重度干扰生境分别捕获成虫251只(6科29种)、276只(6科27种)和352只(6科29种);所有干扰梯度生境中均以步甲科、葬甲科和隐翅虫科为优势科,3种干扰梯度间成虫个体数和物种数均无显著差异(表2)。基于三因素方差分析的主体间效应检验表明林型(F=6.088,P<0.05)和月份(F=31.441,P<0.05)对鞘翅目成虫群落个体数量影响显著,干扰梯度对群落个体数量无显著影响,三者之间的交互作用对群落个体数量均无显著影响;同时,主效应检验表明月份(F=60.454,P<0.05)对鞘翅目成虫群落物种数影响显著,干扰梯度、林型及三者之间交互效应对群落物种数无显著影响。
7月共捕获地表鞘翅目成虫441只(7科25种),其中以步甲科、葬甲科和隐翅虫科为优势科;8月共捕获成虫323只(5科28种),以步甲科、葬甲科和隐翅虫科为优势科;10月共捕获成虫115只(5科15种),其中以隐翅虫科为优势科。10月的个体数和物种数分别与7月、8月具有显著差异(P<0.05)。
2.2 各林型地表鞘翅目成虫水平分布及季节动态
7月,RL林型鞘翅目成虫个体数高于其他5个林型,YL个体数为6种林型中最少,RL个体数除了与KY差异不显著外与其他林型均具有显著差异(P<0.05),且与YL差异极显著(P<0.01),其他林型之间个体数量无显著差异。8月,RL林型鞘翅目成虫个体数与RHS和YL分别具有显著差异(P<0.05),RHS与ZF、ZF与YL具有显著差异(P<0.05),其他林型之间个体数量无显著差异,RHS和YL林型个体数低于其他4个林型。10月份各样地捕获个体数量都较少,RL个体数与KY和BH分别具有显著差异(P<0.05),其他样地间个体数量无显著差异。7月和8月,步甲科和葬甲科在数量上占据优势,而在10月份则以隐翅虫科占据明显优势。
7月,RL林型物种数分别与YL、ZF具有显著差异(P<0.05);8月和10月,6个林型之间物种数均无显著差异。对比分析相同林型不同月份的差异,6个林型的地表鞘翅目成虫物种数从7月到8月存在不同程度下降,而10月物种数相较7月和8月下降幅度更大,具体表现为:KY林型物种数7月和8月差异显著(P<0.05),RL林型物种数7月和8月具有显著差异(P<0.05),其他林型7月的物种数和8月的物种数无显著差异,各样地10月份的物种数分别与7月、8月具有显著差异(P<0.05)(图1)。
表2不同林型、干扰梯度及月份间地表鞘翅目成虫群落个体数和物种数差异比较
Table2DifferencesforindividualsandspeciesnumbersofgroundColeopteraadultsamongdifferentforesthabitats,disturbancegradientsandmonths
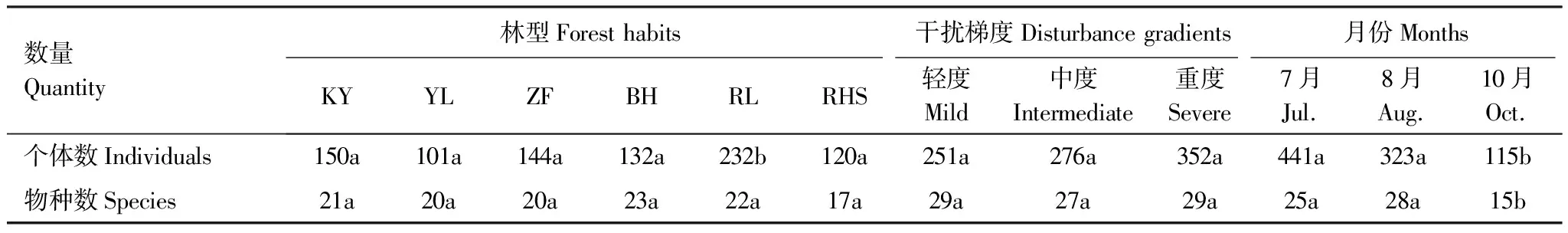
数量Quantity林型Forest habits干扰梯度Disturbance gradients月份MonthsKYYLZFBHRLRHS轻度Mild中度Intermediate重度Severe7月Jul.8月Aug.10月Oct.个体数Individuals150a101a144a132a232b120a251a276a352a441a323a115b物种数Species21a20a20a23a22a17a29a27a29a25a28a15b
同一行字母相同表示差异不显著,字母不同表示差异显著(P<0.05)

图1 6种林型不同月份地表鞘翅目成虫个体数和物种数Fig.1 Individuals and species numbers for ground Coleoptera adults in different months of six forest habitats字母A、B、C表示同一月份不同林型间差异性,a、b、c表示同一林型不同月份间差异性,有相同字母表示无显著差异,字母不同表示有显著差异; KY:原始阔叶红松林virgin mixed broadleaved-Korean pine forest; YL:谷地云冷杉林valley spruce-fir forest; ZF:阔叶红松择伐林selectively cut mixed broadleaved-Korean pine forest; BH:次生白桦林secondary birch forest; RL:落叶松人工林Dahurian larch plantation; RHS:红松人工林Korean pine plantation
2.3 地表鞘翅目成虫群落多样性
7月,RL林型DG指数分别与YL、ZF、BH具有显著差异(P<0.05),其他林型间无显著差异,8月或10月各林型间DG指数均无显著差异;三次调查,7、8、10月所有群落的DG指数均呈现不同程度降低,尤其以10月DG指数降低幅度更大。7月RHS林型多样性指数H′分别与BH、ZF差异显著(P<0.05),其他林型之间无显著差异;8月或10月6个林型之间的H′均无显著差异。总体看,均匀度指数J和优势度指数C基本表现为各月份6种林型之间无显著差异。
对比多样性指数在相同林型不同月份间的差异性,RL林型3个月间DG指数均有显著差异(P<0.05),ZF林型在8月和10月间DG指数具有显著差异(P<0.05)。6种林型的H′指数基本表现为7月与8月差异不显著,10月分别与7月、8月具有显著差异(P<0.05);J指数表现为ZF、BH在7月和10月间具有显著差异(P<0.05),其他林型3个月份之间J指数均无显著差异;C指数表现为YL、ZF、RHS林型10月分别与7月和8月差异显著(P<0.05),其他林型3个月份间均无显著差异(表3)。
相同调查月份中不同干扰梯度之间多样性指数基本表现为差异不显著,而相同干扰梯度的不同月份之间多样性指数存在差异,表现为10月分别与7月或8月差异显著(P<0.05)(表3)。主体间效应检验表明月份对多样性指数DG(F=26.372,P<0.05)、H′(F=48.123,P<0.05)、C(F=25.714,P<0.05)和J(F=3.502,P<0.05)均具有显著影响,林型和干扰梯度则对多样性指数影响不显著;同时,三者之间交互作用对群落多样性指数也无显著影响。
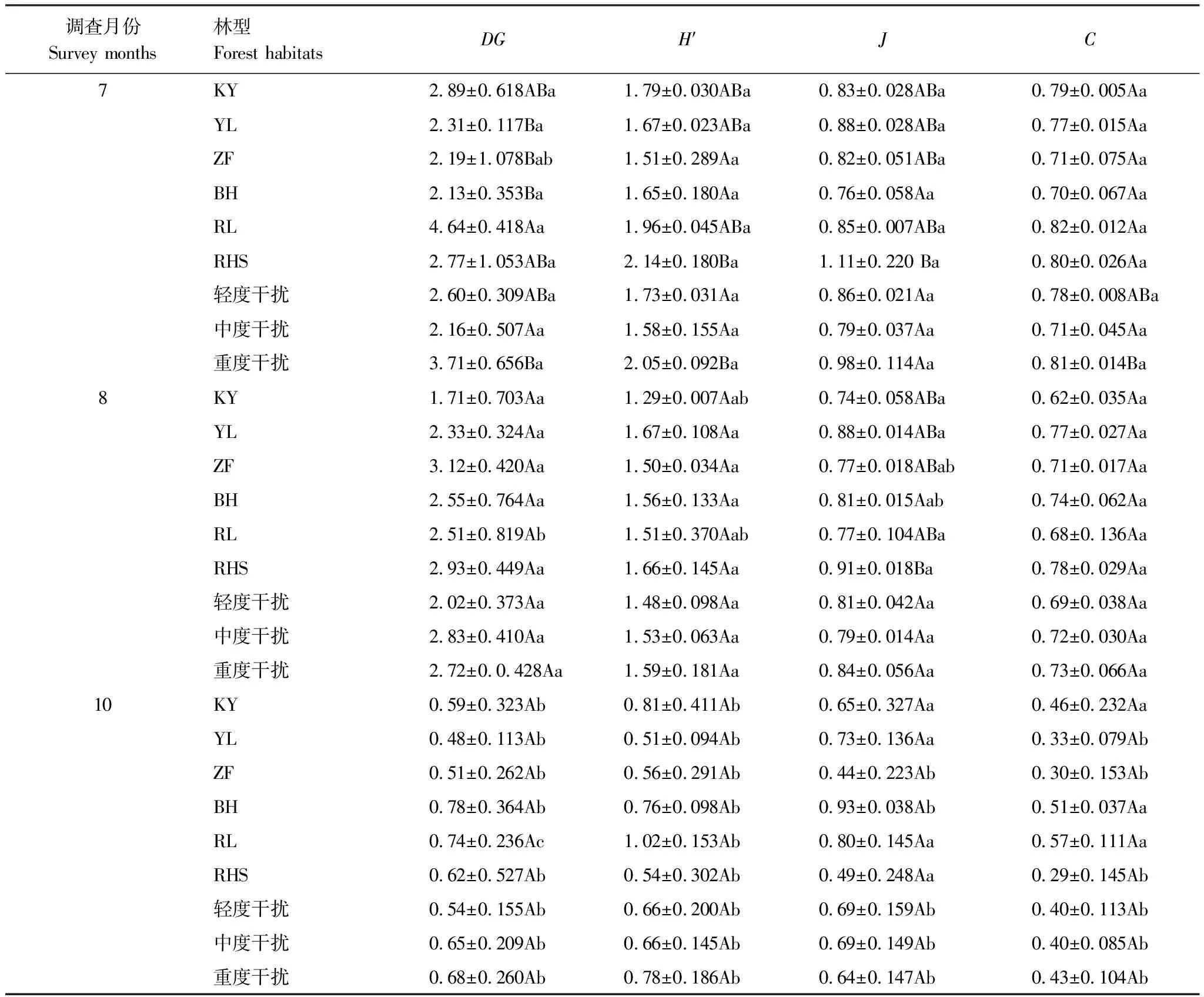
表3 不同月份、林型和干扰梯度的地表鞘翅目成虫群落多样性指数
同一列字母A、B、C表示相同月份不同林型/不同干扰梯度间差异,a、b、c表示相同林型/相同干扰梯度的不同月份间差异,有相同字母则表示无显著差异,字母不同则表示有显著差异(P<0.05);DG:密度—类群指数Density-Group index,H′:Shannon指数Shannon-wiener index,J:均匀度指数Pielou index,C:优势度指数Simpson index
2.4 群落相似性和聚类分析
Jaccard系数表明若只考虑群落物种组成上的相似性,7月份,RHS、YL分别与RL为中等不相似,RHS、KY、YL分别与BH为中等不相似,其他林型之间均为中等相似,其中以RL和KY相似程度最高(0.688);8月份,仅RL与YL为中等相似(0.500),其他几个林型之间均为中等不相似;10月份,YL、BH分别与ZF为中等相似,RHS、YL分别与BH为中等相似,其他林型之间为中等不相似或极不相似,其中KY与BH的相似性最低(0.125)。
若加入个体数量因素,Gower系数Sg表明各林型间的相似性明显比Jaccard系数q低。7月份以ZF与KY间的相似性最高(0.453),以RL与YL间的相似性最低(0.240);8月份和10月份,各林型之间的相似性都较低;可以看出,无论7月、8月或10月,以Gower系数Sg表示的不同林型间相似性都较低(Sg<0.5)(表4)。
聚类分析结果表明RHS与YL最为相似,ZF相似度次之,BH、RL、KY与其他样地相似度较低;其中7月以ZF和YL最为相似,KY次之,RHS、RL和BH与其他林型相似程度较低;8月则表现为YL和BH最为相似,ZF次之,RL、RHS和KY与其他林型差异较大;10月以YL和BH最为相似,RHS次之,ZF、KY和RL与其他林型相似度较低(图2)。

表4 不同林型地表鞘翅目成虫群落Jaccard系数与Gower系数
无括号的数字为Jaccard系数q,括号中的数字为Gower系数Sg

图2 各月份地表鞘翅目成虫群落聚类图(平方Euclidean距离)Fig.2 Clustering figures (based on square Euclidean distance) for ground Coleoptera adults communities in different months
3 讨论
3次调查共捕获鞘翅目成虫879只,分属9科44物种,依据各林型鞘翅目成虫个体数量多少排序RL>KY>ZF>BH>RHS>YL,RL林型个体数量高于其他林型,YL林型个体数量为6种林型中最低。轻度干扰生境共捕获成虫251只(6科29种),属于轻度干扰生境的KY和YL个体数量差异不显著;中度干扰生境捕获成虫276只(6科27种),属于中度干扰生境的ZF和BH个体数量较接近,差异不显著;重度干扰生境共捕获成虫352只(6科29种),同属重度干扰生境的RHS与RL个体数量差异显著。三因素方差分析表明林型和月份对鞘翅目成虫群落结构有显著影响,而干扰梯度对鞘翅目成虫群落结构无显著性影响,并且进一步的LSD多重比较表明不同干扰梯度处理中鞘翅目群落无显著差异,不同林型或不同月份之间鞘翅目群落个体数、物种数及多样性存在显著差异性,说明干扰并不是影响地表鞘翅目成虫群落组成的重要因素,林型和季节动态才是影响鞘翅目成虫群落结构的重要因素。
进一步对多样性指标进行分析,受人为干扰程度最轻的原始林(KY和YL)生态系统结构复杂,植被多样性较高,生物多样性理论上应该更高[20],但实际3次调查中KY和YL林型地表鞘翅目成虫的多样性指数H′和DG指数并不是6个林型中最高,尤其YL的DG指数较低;结合3个月份的Jaccard系数、Gower系数以及聚类分析结果来看,具有相同干扰梯度的KY和YL林型间相似程度并不高。BH是经过中度干扰后天然更新恢复良好的次生林生态系统,其7月份和8月份鞘翅目物种较为丰富,但10月份是较寒冷的调查季节,多样性指数明显下降,同为中度干扰梯度的ZF林型在8、10月份的多样性指数与BH具有很大差异性,相似性指数和聚类分析结果表明,具有相似干扰梯度的次生林ZF和BH在7月、8月或10月中没有表现出较高的相似性;ZF和BH属于中度干扰模式下的植被群落,但二者鞘翅目成虫多样性相对其他干扰模式下的群落并无显著优势。人工林代表着完全人为干扰模式下的生态系统,这样的生态系统树种单一、结构简单,通常生物多样性会较低[21],但对于凉水地区落叶松人工林(RL)并不如此,在所有调查中RL的鞘翅目成虫个体数量和多样性指数高于其他几个林型;RHS是人工林生态系统,但其地表鞘翅目成虫多样性与RL存在较大差异,并且属于同一干扰梯度的RHS和RL在3个月份中所表现出的相似性都较低。另外,具有相似林龄的BH、RL和RHS(62a—63a)地表鞘翅目成虫群落并不相似,具有相同林龄的KY和YL(300a以上)地表鞘翅目成虫群落结构也不相似,表明各群落间的相似程度与林龄没有直接对应的关系。3次调查中,各林型鞘翅目成虫多样性没有表现出随干扰梯度升高或降低的相关规律,相似性分析和聚类分析结果不能将相同干扰梯度的鞘翅目成虫群落归为一类,表明干扰对群落多样性并无显著影响。
具有相同干扰梯度的森林生态系统地表鞘翅目群落结构没有表现出高度相似性,不同干扰梯度生境鞘翅目群落组成差异不明显,且干扰对群落物种个体数量、物种数量和多样性均没有显著影响,表明干扰梯度不是造成各林型间地表鞘翅目成虫群落结构差异的关键因素,即凉水地区鞘翅目成虫群落多样性不符合中度干扰假说。据此归纳原因:谷地云冷杉林(YL)虽受干扰较轻,但云冷杉林多高大乔木,林木郁闭度较高,且该生境处于地势相对低洼的谷地,尤其7、8月份多雨,样地会不同程度被水淹没,土壤动物生存环境质量低下,严重影响了地表大型土壤动物多样性[22- 23],使得该林型鞘翅目成虫群落多样性较低;原始阔叶红松林(KY)内有游客临时踩踏形成的山路,相对较多的人员出入导致部分地表植被不同程度的破坏,可能使得鞘翅目成虫群落多样性下降。中度干扰梯度的次生白桦林(BH)植被覆盖度较高,其7月和8月物种较为丰富,但10月份是较寒冷的调查季节,食物资源减少、地表温湿度等适宜性明显降低,个体数和物种数明显下降;次生白桦林和红松择伐林(ZF)虽属于同一种干扰模式的次生林,但实际二者样地内具体的植被状况、地形、土壤温度及含水量差异较大[13],这可能是造成白桦林与红松择伐林鞘翅目成虫多样性存在差异的主要原因。落叶松人工林(RL)位于小于25°的向阳斜坡上,林地内虽树种单一,但地表草本和灌木丰富,其土壤疏松,孔隙度大,土壤排水状况良好,位于阳坡,地表温度适宜,且落叶松林凋落物也较为丰富,有利于地表大型土壤动物的栖息与繁衍,其地表大型土壤动物群落的个体数和多样性都较高。这与刘继亮等[24]的研究结果类似,刘继亮等对秦岭西部山地次生林和人工林大型土壤动物群落特征的研究发现,油松林和日本落叶松人工林土壤动物密度显著高于次生林的土壤动物密度,日本落叶松林虽是人工林,但其凋落物厚度、土壤养分含量和土壤孔隙度均较高,为土壤动物生存提供有利条件;红松人工林(RHS)地表鞘翅目成虫群落多样性低于落叶松人工林,原因可能在于红松人工林相比落叶松人工林地表植被覆盖度低,样地内地表裸露面积较大,土壤相对瘠薄[13]。
本实验还表明:10月份步甲科和葬甲科的个体数量及物种数较之7、8月份大幅下降,温度降低可能是造成10月份步甲科和葬甲科物种及个体数量减少的决定因素;而在10月份6个群落中隐翅虫科都是唯一的优势科,数量较7、8月份有所增加,说明在调查的低温环境条件下,隐翅虫科比步甲科、葬甲科等其他物种具有更宽的生态位。就不同分析指标来看6种林型样地共有物种数较多,但有些共有物种的相应个体数相差较大,因此,用于反映群落物种上相似性的Jaccard相似性系数q明显比反映群落物种个体数量上的相似性的Gower系数Sg要高,表明不同林型间地表鞘翅目成虫个体数的异质性明显比物种数的高。Jaccard相似性系数q的变化范围为0.1250—0.6875,Gower相似性系数Sg的变化范围为0.0522—0.4525,说明该区6个林型地表鞘翅目成虫群落异质性明显。6个地表鞘翅目成虫群落中稀有物种数多于优势物种数和常见物种数,稀有物种数共29种(占总物种数65.91%),这证实了群落的异质性愈明显,大型土壤动物类群中稀有类群数就愈多这一观点[25- 28]。
4 结论
本实验对凉水自然保护区6种林型的地表鞘翅目成虫群落组成进行研究,三次调查表明地表鞘翅目成虫群落个体数量、物种数和群落多样性不具有随干扰强度变化而变化的梯度性规律,说明地表鞘翅目成虫群落多样性并不支持中度干扰假说。人为干扰梯度不是直接影响该区地表鞘翅目成虫群落结构的主要因素,而林型和时间(月份)则是影响地表鞘翅目成虫群落组成的重要因素。
致谢:朱纪元、闫龙、曹阳、胡媛媛、张超、刘俊同学帮助实验,凉水保护区生态站提供实验便利,特此致谢。
参考文献(References):
[1] 杨利民, 周广胜, 王国宏, 王玉辉. 人类活动对榆树疏林土壤环境和植物多样性的影响. 应用生态学报, 2003, 14(3): 321- 325.
[2] 刘会玉, 林振山, 张明阳. 人类周期性活动对物种多样性的影响及其预测. 生态学报, 2005, 25(7): 1635- 1641.
[3] Fox J F, Connell J H. Intermediate-disturbance hypothesis. Science, 1979, 204(4399): 1344- 1345.
[4] Sven E J, Brian F. Encyclopedia of Ecology. USA: Elsevier Science, 2008: 1986-1994.
[5] Roxburgh S H, Shea K, Wilson J B. The intermediate disturbance hypothesis.: Patch dynamics and mechanisms of species coexistence. Ecology, 2004, 85(2):359- 371.
[6] Townsend C R, Scarsbrook M R, Dolédec S. The intermediate disturbance hypothesis, refugia, and biodiversity in streams. Limnology and Oceanography, 1997, 42(5):938- 949.
[7] Molino J F, Sabatier D. Tree diversity in tropical rain forests: a validation of the intermediate disturbance hypothesis. Science, 2001, 294(5547): 1702- 1704.
[8] 武崎, 吴鹏飞, 王群, 文勇立, 高艳美, 张荣芝, 龙伟. 放牧强度对高寒草地不同类群土壤动物的群落结构和多样性的影响. 中国农业科学, 2016, 49(9): 1826- 1834.
[9] 王广力, 王勇, 韩立亮, 张美文, 李波. 洞庭湖区不同土地利用方式下的土壤动物群落结构. 生态学报, 2005, 25(10): 2629- 2636.
[10] 吴玉红, 蔡青年, 林超文, 赵欣, 程序. 四川紫色土丘陵区不同土地利用方式下中型土壤动物群落结构. 生态学杂志, 2009, 28(2): 277- 282.
[11] 张智英, 张亮, 李玉辉, 何晓岚. 云南石林喀斯特景区不同生境大型土壤动物多样性研究. 林业科学研究, 2005, 18(6): 701- 705.
[12] 吴东辉, 张柏, 陈鹏. 长春市不同土地利用条件下大型土壤动物群落结构与组成. 动物学报, 2006, 52(2):279- 287.
[13] 史宝库, 金光泽, 汪兆洋. 小兴安岭5种林型土壤呼吸时空变异. 生态学报, 2012, 32(17): 5416- 5428.
[14] 马建章, 刘传照, 张鹏. 凉水自然保护区研究: 第一集. 哈尔滨: 东北林业大学出版社, 1993.
[15] 李井科, 林琳, 张雪萍. 原色中国东北土壤甲虫图鉴——步行虫类. 哈尔滨: 哈尔滨地图出版社, 2015.
[16] 李井科, 张利敏, 张雪萍. 原色中国东北土壤甲虫图鉴——隐翅虫类拟步甲类. 哈尔滨: 哈尔滨地图出版社, 2015.
[17] 李景科, 张雪萍, 林琳, Flutsch G, 于强. 中国东北的葬甲科研究. 哈尔滨: 哈尔滨地图出版社, 2011.
[18] 尹文英, 胡圣豪, 沈韫芬, 宁应之, 孙希达, 吴纪华, 诸葛燕, 张云美, 王敏, 陈建英, 徐成刚, 梁彦龄, 王洪铸, 杨潼, 陈德牛, 张国庆, 宋大祥, 陈军, 梁来荣, 胡成业, 王慧芙, 张崇洲, 匡溥人, 陈国孝, 赵立军, 谢荣栋, 张骏, 刘宪伟, 韩美贞, 毕道英, 肖宁年, 杨大荣. 中国土壤动物检索图鉴. 北京: 科学出版社, 1998.
[19] Raunkiaer C. The life forms of plants and statistical plant geography. Oxford:Oxford University Press,1934.
[20] 樊正球, 陈鹭真, 李振基. 人为干扰对生物多样性的影响. 中国生态农业学报, 2001, 9(2): 31- 34.
[21] 方升佐, 田野. 人工林生态系统生物多样性与生产力的关系. 南京林业大学学报自然科学版, 2012, 36(4):1- 6.
[22] 王玉, 高光彩, 付必谦, 吴专. 北京野鸭湖湿地地表甲虫群落组成与空间分布格局. 生物多样性, 2009, 17(1): 30- 42.
[23] 于晓东, 罗天宏, 周红章, 杨建. 边缘效应对卧龙自然保护区森林-草地群落交错带地表甲虫多样性的影响. 昆虫学报, 2006, 49(2): 277- 286.
[24] 刘继亮, 曹靖, 李世杰, 潘春林, 潘成臣. 秦岭西部山地次生林和人工林大型土壤动物群落结构特征. 应用生态学报, 2012, 23(9): 2459- 2466.
[25] 路有成, 王宗英. 九华山土坡动物的垂直分布. 地理研究, 1994, 13(2): 74- 81.
[26] 王宗英, 陈发扬, 路有成, 聂献忠. 九华山森林土壤蜘蛛群落的初步研究. 生态学报, 1997, 17(l): 71- 77.
[27] 傅必谦, 陈卫, 董晓辉, 邢忠民, 高武. 北京松山四种大型土壤动物群落组成和结构. 生态学报, 2002, 22(2): 215- 223.
[28] 张雪萍, 黄丽荣, 姜丽秋. 大兴安岭北部森林生态系统大型土壤动物群落特征. 地理研究, 2008, 27(3): 509- 518.
