逆境条件下镉对臭牡丹幼苗生理变化的影响
2018-06-15张婷婷崔乐怡龙昌莲陈建华
张婷婷,崔乐怡,龙昌莲,张 航,张 琪,苟 秋,陈建华,刘 鹏
(1.浙江师范大学植物学实验室,浙江 金华 321004;2.浙江省野生动物生物技术与保护利用重点实验室,浙江 金华 321004)
镉(Cd)作为生物毒性最强的重金属元素之一,在土壤中具有化学活性强、移动性差、毒性持久等特征。目前,我国重金属镉污染问题十分严重,污染耕地面积超过1.33万hm2,镉含量超标的农产品年产量达14.6亿kg,用镉污染水灌溉的农田面积达330多万hm2,浙江省镉超标土壤占10.69%[1]。重金属镉在土壤中的环境容量小且易被植物吸收,这不仅严重影响农作物的产量与品质,还随着食物链最终进入人体,对人体健康造成了极大危害[2]。
臭牡丹(Clerodendron bungeiSteud.)是马鞭草科(Verbenaceae)落叶小灌木,是一种兼顾观赏性和药用价值的植物,在中医药方面具有活血化瘀、清热利湿、祛风解毒等多种功效,现代医学研究指出,其还具备镇静催眠、抗肿瘤、抗氧化和治疗肾炎及关节炎等功能[3]。此外,臭牡丹喜温暖潮湿、抗逆性强且对水肥的要求不高,在城市环境美化及水土保持方面均发挥着重要作用。近年来,许多学者对臭牡丹提取物进行药理作用分析,如余娜等[4]探讨臭牡丹总黄酮对人肺癌A549细胞增殖、迁移和侵袭作用与相关机制,刘建新等[5]研究指出臭牡丹根正丁醇提取物具有显著的抑菌作用。目前,国内外对臭牡丹的研究多集中于化学成分和药用功效方面,对臭牡丹抗逆性的研究较少。张建新等[6]对遮光条件下臭牡丹生理变化进行探讨,但主要集中在部分生长和光合性能方面,针对重金属镉逆境对臭牡丹生理变化影响的研究至今未见报道。因此,本试验以臭牡丹幼苗为试材,测定镉胁迫下植株叶绿素荧光参数、丙二醛含量、脯氨酸含量、可溶性糖含量及可溶性蛋白含量等生理指标,以期为进一步研究植物抗逆性机理奠定基础,并为臭牡丹的实际生产与推广应用提供可靠的理论依据。
1 材料与方法
1.1 试验材料
供试臭牡丹来自江苏省海门市三厂镇,扦插繁殖培育1年后选取健康且大小长势相似的幼苗栽植于塑料花盆中,每盆2株。
1.2 试验方法
试验于2017年6~7月在浙江师范大学植物学实验室进行。对臭牡丹幼苗进行镉胁迫处理,设0、0.1、0.3、0.6、0.9 mmol/L 5个镉胁迫浓度处理,3次重复,各重金属溶液均用去离子水配制。盆栽试验期间每隔2 d浇灌1次镉溶液,每次每盆40 mL。胁迫处理12、24、36 d分别对臭牡丹幼苗叶绿素荧光参数、脯氨酸含量、丙二醛(MDA)含量、可溶性糖含量、可溶性蛋白含量等生理指标进行测定。
1.3 测定项目及方法
叶绿素荧光参数采用便携式叶绿素荧光仪MINI-PAM进行测定:测定前对植物活体叶片进行暗处理10 min,在叶片上选取中部区域进行测定,本试验主要探讨PSⅡ潜在活性(Fv/Fo)、光化学淬灭系数(qP)和非光化学淬灭系数(qN)。MDA含量采用硫代巴比妥酸比色法[7]测定;脯氨酸含量参照磺基水杨酸比色法测定;可溶性糖含量采用蒽酮比色法测定;可溶性蛋白含量采用考马斯亮蓝比色法[8]测定。
1.4 数据处理
试验数据使用Excel 2013进行整理,并运用SPSS 20.0统计软件进行分析。通过单因素方差分析法(one-way ANOVA)和Duncan法计算平均值和标准误,分析差异显著性,同时用Origin 8.5软件作图。
2 结果与分析
2.1 镉胁迫对臭牡丹叶绿素荧光参数的影响
在荧光诱导动力学参数中,Fv/Fo是叶片PSⅡ量子效率,反映植物PSⅡ系统的潜在活力[9]。光化学猝灭系数(qP)是对PSⅡ反应中心原初电子受体氧化态的一种度量[10],而非光化学猝灭系数(qN)代表PSⅡ热耗散能力[11]。由图1可知,随着镉浓度的升高,臭牡丹叶片Fv/Fo和qP总体上均呈现出下降趋势,在0.9 mmol/L镉浓度处理下数值最低。其中,不同胁迫周期臭牡丹叶片Fv/Fo较对照的最大降幅分别为22.70%、25.61%和43.10%,而不同胁迫周期臭牡丹叶片qP较对照的最大降幅则分别为37.82%、36.99%和26.20%。叶片qN随镉浓度的增加而增加,相较于对照其不同周期增幅分别为34.76%、48.13%和43.10%。表明镉胁迫下臭牡丹光合电子传递速率下降,其潜在光合活性受到抑制,热耗散作用加强。

图1 不同浓度镉胁迫处理对臭牡丹叶片Fv/Fo、qP、qN的影响
2.2 镉胁迫对臭牡丹MDA含量的影响
MDA是细胞膜脂过氧化的最终产物,是植物膜系统完整性的重要指标[12]。MDA含量越大,植株膜脂过氧化现象越严重,膜系统受损程度越大。由表1可知,MDA含量随镉胁迫浓度的升高而有所增长,均在0.9 mmol/L浓度下达到最大值。但不同周期镉胁迫下,植株MDA含量增长趋势有明显差异,胁迫前期和中期臭牡丹叶片MDA含量上升趋势平缓,胁迫后期其增长趋势则较为显著。胁迫36 d时,0.1、0.3、0.6、0.9 mmol/L镉浓度处理臭牡丹MDA含量较对照上升9.09%、26.07%、38.52%和45.29%。可见,高浓度、长时间的镉污染致使臭牡丹产生膜脂过氧化现象,损伤植物膜系统完整性。
2.3 镉胁迫对臭牡丹脯氨酸含量的影响
游离脯氨酸是一种广泛存在于植物体内的偶极含氮化合物,具有保护细胞膜系统和维持胞内酶结构的功能,有利于植物抵御逆境胁迫[13]。从表1可以看出,在同一胁迫周期中,臭牡丹叶片脯氨酸含量随着镉胁迫浓度的升高而不断增加。随着胁迫时间的变化,臭牡丹叶片脯氨酸含量增长趋势更为显著。镉胁迫12 d时,臭牡丹脯氨酸含量于0.9 mmol/L处理浓度下达到最大值,较对照增长90.48%。而胁迫处理24 d和36 d时,植株脯氨酸含量变化趋势更加显著,较对照其最大增长率分别高达308.75%和331.49%。由此可见,镉胁迫刺激臭牡丹累积脯氨酸,调节细胞渗透势,从而在一定程度上抵御外界渗透胁迫。
2.4 镉胁迫对臭牡丹可溶性糖含量的影响
在重金属污染环境下,植物为缓解水分流失,细胞会主动累积一些可溶性物质来降低胞内渗透势[14]。其中,可溶性糖是植物的重要渗透调节物质,其累积有利于植物适应外界环境[15]。由表2可知,臭牡丹可溶性糖含量随着镉处理浓度的升高呈现出先升后降的趋势。植株可溶性糖含量均在0.6 mmol/L镉浓度下达到最大值,与对照相比,不同周期植株可溶性糖含量增长率分别为24.39%、39.01%和36.26%。当镉胁迫浓度大于0.6 mmol/L时,不同胁迫时间下臭牡丹可溶性糖含量均有所下降,但都高于对照,相较0.6 mmol/L胁迫处理其降幅分别为33.48%、33.29%和27.49%。说明一定浓度范围内的镉胁迫能够促使臭牡丹叶片可溶性糖含量升高,但过高浓度的镉则对其表现出抑制效应。
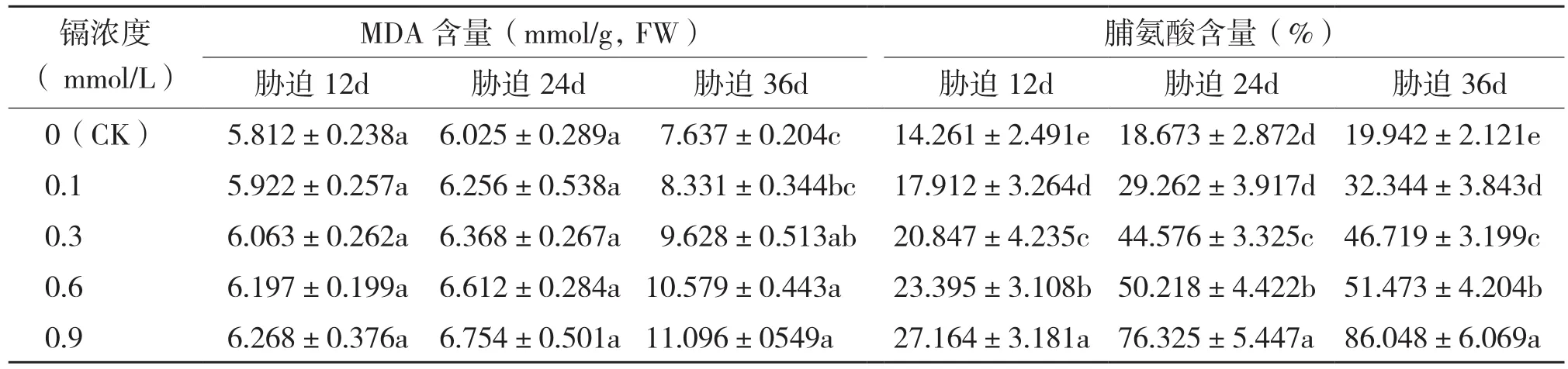
表1 不同浓度镉处理对臭牡丹叶片MDA含量及脯氨酸含量的影响
2.5 镉胁迫对臭牡丹可溶性蛋白含量的影响
可溶性蛋白具有较强亲水性,能够增强细胞持水能力,是一种植物体内重要的渗透调节物质[16]。从表2可以看出,随着镉胁迫浓度的升高,臭牡丹可溶性蛋白积累量先升后降。在不同处理周期中,可溶性蛋白含量均在0.6 mmol/L胁迫处理时达到最大值,与对照相比,其最大增长率分别为73.07%、48.25%和28.99%。当镉胁迫浓度超过0.6 mmol/L时,臭牡丹可溶性蛋白含量下降,但均高于对照,相较于0.6 mmol/L胁迫处理其降幅分别为7.68%、7.63%和10.35%。说明一定浓度范围内的镉逆境能够刺激臭牡丹积累可溶性蛋白缓解细胞水分散失,但高浓度镉则抑制臭牡丹可溶性蛋白的合成。
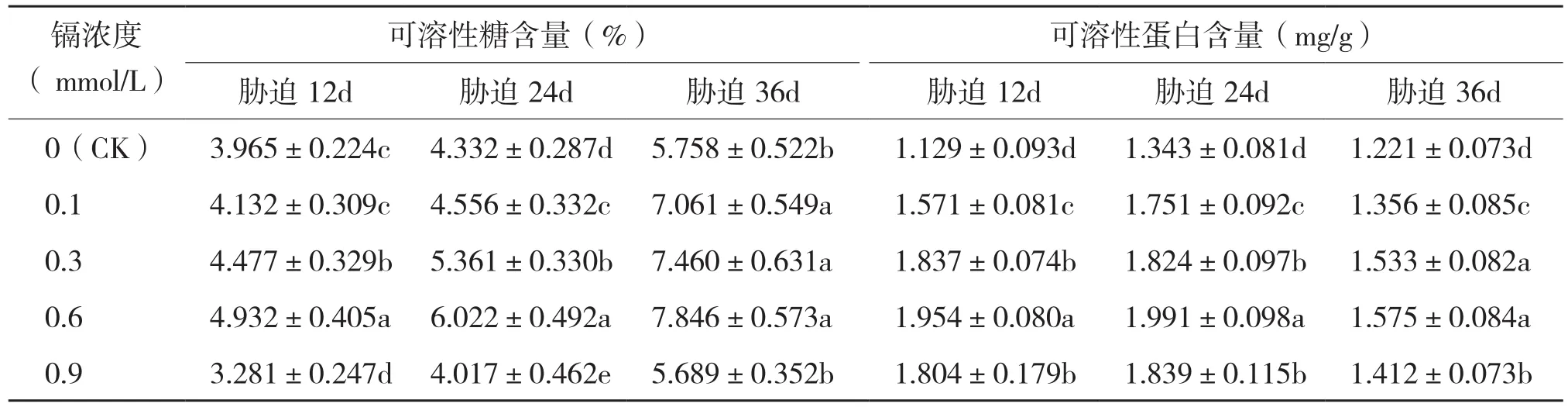
表2 不同浓度镉处理对臭牡丹叶片可溶性糖含量及可溶性蛋白含量的影响
3 结论与讨论
镉是植物生长过程中的一种非必需元素,易被植株根系吸收,过量的镉对植物生理状态和和新陈代谢具有较强毒害作用[17]。叶绿素荧光与植物光合作用各个过程紧密相关,是检测光合系统损伤程度的灵敏探针[18],其动力学参数能够快速地探测逆境对植物PSⅡ反应中心的影响。其中,Fv/Fo是植株光化学反应状况的一项重要参数,代表植物PSⅡ光化学系统的潜在活力。本试验结果显示,臭牡丹叶片Fv/Fo随着镉处理浓度升高而不断下降,说明镉污染使PSⅡ中心受到光抑制,其潜在活性减弱,电子传递活性下降。qP反映植物PSⅡ原初电子受体QA的还原状态及PSⅡ开放中心数目,其值越大则光合电子传递速度越快。而qN代表植物PSⅡ天线色素吸收后不能用于光合电子传递而以热形式消散的份额,其值升高有利于保护PSⅡ免受光氧化伤害。本试验结果表明重金属镉胁迫促使臭牡丹叶片qP持续下降,qN不断上升,这与梁开明等[19]的研究结果基本一致,也说明在镉胁迫下臭牡丹叶片PSⅡ反应中心开放程度减小,光合电子传递能力降低,同时其将过剩的光能转换成热能散失,从而在一定程度上提高自身对镉的耐受能力。关于镉胁迫对臭牡丹叶绿素荧光参数的影响,本试验取得了一定进展,今后可在此基础上结合植株气孔运动状态进行更深入的探讨。
丙二醛(MDA)是植物组织遭受氧化胁迫发生膜脂过氧化作用的产物,能够交联核酸、糖类及蛋白质等物质,从而损伤细胞膜系统的完整性。因此,MDA含量是反映细胞膜过氧化作用强弱和质膜破坏程度的重要指标[20]。在本试验中,臭牡丹叶片MDA含量在胁迫后期随镉浓度的增加而增加,这与在朱砂根(Ardisia crenataSims)上的研究结果[21]基本相同,也说明长期镉胁迫致使臭牡丹体内活性氧平衡被打破,产生的大量自由基对植物细胞质膜透性产生严重破坏,脂类物质被氧化分解,膜脂过氧化作用加剧。在重金属污染环境中,土壤渗透势较低,植物细胞易出现失水现象。为了抵御渗透胁迫,植物会主动累积一些渗透调节物质来降低胞内渗透势,从而维持细胞的正常生理功能。其中,可溶性糖、可溶性蛋白及脯氨酸都是重要的渗透调节物质。试验结果表明,较低浓度的镉胁迫刺激臭牡丹积累可溶性糖及可溶性蛋白,而高浓度镉则使臭牡丹可溶性糖和可溶性蛋白含量降低,说明镉胁迫刺激植物启动防御机制,可溶性糖、可溶性蛋白作为渗透调节物质保护细胞免受伤害,但高浓度镉对臭牡丹的伤害较大,抑制了可溶性糖和可溶性蛋白的合成。在不同浓度镉胁迫对植物脯氨酸累积的影响方面,何俊瑜等[22]在对龙葵(Solanum nigrumL.)的探究中发现,镉胁迫可促进龙葵积累脯氨酸,且高镉胁迫下其积累效应更加显著。而在本试验中,脯氨酸含量随镉胁迫浓度的增加不断升高,这与上述在龙葵上的研究趋势基本一致,说明在高镉胁迫下脯氨酸作为一种重要的渗透调节物质,发挥着缓解细胞渗透胁迫的重要作用,在一定程度上增强了植株对镉的抗逆性。
综上所述,重金属镉胁迫对臭牡丹的叶绿素荧光参数、膜脂过氧化程度、渗透调节物质累积都具有一定程度的影响。在镉逆境下,臭牡丹降低光合活性,将过量光能转化为热能,大量累积渗透调节物质,从而增强自身抗逆性。但长时间的高镉胁迫会导致植物光合系统受损、膜脂过氧化现象加剧、渗透调节物质合成受阻。在臭牡丹对镉胁迫的生理响应方面,本试验结果表明光合特性的调整与渗透调节物质的积累在植物抵御外界逆境胁迫方面具有重要作用,这是植物进行能量代谢及细胞抵御渗透胁迫的主要方式,也是植物耐镉机理中的重要生理反应。重金属镉胁迫下植物的耐受机制是一个复杂的生理体系,后续的试验应在现阶段的基础上综合植株气孔运动、内源激素调节、DNA损伤过程等方面作进一步研究。
[1]刘洋,张玉烛,方宝华,等.栽培模式对水稻镉积累差异及其与光合生理关系的研究[J].农业资源与环境学报,2014,31(5):450-455.
[2]Ingwersen J,Streck T.A regional-scale study on the crop uptake of cadmium from sandy soils:measurement and modeling[J].Journal of Environmental Quality,2005,34(3):1026-1035.
[3]陈思勤,朱克俭,李勇敏,等.臭牡丹总黄酮抑制小鼠Lewis肺癌实体瘤及其与p53、bcl-2、bax表达相关性研究[J].世界中医药,2016,11(6):946-949.
[4]余娜,朱克俭,马思静,等.臭牡丹总黄酮通过调控Wnt/β-catenin通路影响A549细胞上皮间质转化研究[J].中草药,2018,49(3):663-670.
[5]刘建新,李燕,连磊凡,等.臭牡丹根正丁醇提取物的体外抗菌实验的研究[J].时珍国医国药,2015,26(8):1849-1850.
[6]张建新,颜赟,方炎明.遮光对臭牡丹生长和光合特性的影响[J].植物资源与环境学报,2013,22(1):88-93.
[7]张志良.植物生理学实验指导[M].第4版.北京:高等教育出版社,2009.
[8]李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2000.
[9]赵海亮,赵文东,孙凌俊,等.不同架式巨峰葡萄光合特性与叶绿素荧光参数研究[J].西南农业学报,2015,28(6):2691-2695.
[10]徐焕文,刘宇,姜静,等.盐胁迫对白桦光合特性及叶绿素荧光参数的影响[J].西南林业大学学报,2015,35(4):21-26.
[11]吴刘萍,许衍才,廖飞雄,等.杜鹃红山茶叶片叶绿素荧光对干旱胁迫及复水处理的响应[J].广东农业科学,2017,44(10):25-31.
[12]张金燕,孙雪婷,龙光强,等.根系残留对三七植株生长的影响[J].广东农业科学,2017,44(6):27-38.
[13]艾吉尔·阿不拉,李丕军,等.4个造林树种缓苗期生理生化特性分析[J].广东农业科学,2017,44(3):80-86.
[14]韩淑梅,简丽,董旋,等.不同浓度及不同时间镉胁迫对超富集植物孔雀草渗透调节物质的影响[J].山地农业生物学报,2017,36(3):44-48.
[15]范志霞,李绍才,孙海龙.多效唑作用下紫穗槐对干旱胁迫的生理响应及抗旱性评价[J].草业学报,2017,26(3):132-141.
[16]姚佳,刘信宝,崔鑫,等.不同NaCl胁迫对苗期扁蓿豆渗透调节物质及光合生理的影响[J].草业学报,2015,24(5):91-99.
[17]田禹璐,朱宏.重金属镉对植物胁迫的研究进展[J].哈尔滨师范大学自然科学学报,2015,31(2):149-153.
[18]Kieffer P,Dommes J,Hoffmann L,et al.Quantitative changes in protein expression of cadmium-exposed poplar plants[J].Proteomics,2008,8(12):2514-2530.
[19]梁开明,傅玲,章家恩,等.水稻/再力花单、间作系统中作物对镉胁迫的叶绿素荧光响应及镉积累特征[J].华南农业大学学报,2014,35(4):35-41.
[20]卢颖林,陈迪文,江永,等.硅对铝胁迫下甘蔗幼苗的缓解效应研究[J].广东农业科学,2016,43(5):87-91.
[21]张建新,郦枫,马丽,等.镉胁迫下朱砂根和虎舌红生理响应及其镉抗性[J].水土保持学报,2017,31(5):321-327.
[22]何俊瑜,任艳芳,王阳阳,等.不同耐性水稻幼苗根系对镉胁迫的形态及生理响应[J].生态学报,2011,31(2):522-528.
