龙眼果实挂树成熟与退糖期间的解剖学结构比较
2018-06-15韩冬梅李建光李保建吴振先
韩冬梅,曾 婷,李建光,罗 焘,李保建,吴振先
(1.广东省农业科学院果树研究所/农业部南亚热带果树生物学与遗传资源利用重点实验室,广东 广州 510640;2.华南农业大学园艺学院/广东省果蔬保鲜重点实验室,广东 广州 510642;3.江门市农业技术服务中心,广东 江门 529000)
龙眼(Dimocarpus longanLour.)俗称桂圆,成熟于高温乃至酷暑季节,高糖多汁,成熟快,采收期短,适宜的采收成熟度是保证龙眼果实采后品质及贮运寿命的重要因素[1]。采收过早,果实尚未发育完全;采收过晚,果肉含糖量和甜度均明显下降,俗称“退糖”现象。龙眼果实具有完全成熟后很快退糖的特性,在生产上常常因为错过适宜的采收时间而导致果实糖度下降,品质降低[2],不耐贮运,经济效益受损。有关龙眼退糖特性和机理的研究鲜有报道,李建光等[3-4]、韩冬梅等[5]报道了石硖、古山二号和储良龙眼果实的成熟特性,认为龙眼果实完全成熟后可溶性固形物(TSS)、蔗糖和总糖含量逐渐降低,表现出过熟后退糖的特性,其中前两者成熟期较短、退糖快,而储良退糖最慢;同时还研究了石硖和古山二号龙眼果实适宜贮藏成熟度及其完熟、退糖后的果皮横切面结构差异,认为不同成熟度的果皮结构是其耐贮性差异的生物学基础之一[6]。林河通等[7]、屈红霞等[8]也对不同品种成熟时龙眼的果皮结构差异与耐贮性关系开展过研究,认为两者之间存在相关性,而关于成熟及退糖后龙眼果肉结构的变化与品种间差异迄今未见有详细报道。石硖和储良是广东、广西、海南三省两大主栽品种,一般认为石硖较早熟、成熟快、退糖迅速、采收期很短,而储良较晚熟、成熟期长、退糖也慢,因此选择这两个品种,对其果实在挂树成熟与退糖期间的内外果皮表征、果皮横切面、果肉显微组织结构的变化及品种差异进行研究分析,旨在丰富龙眼果实成熟生理发展的解剖学基础,为生产中准确把握适采成熟度和深入研究果实退糖机理提供参考依据。
1 材料与方法
1.1 试验材料
试验于2012—2013年进行两次,供试品种为石硖和储良,采自广东省农业科学院果树研究所龙眼资源圃。选择品种内花期一致、树势相近的单株各2棵,花后挂牌,追踪果实的生长动态(谢花期分别为石硖4月19日、储良4月20日)。当果实发育至50%~60%成熟时开始采果,每隔7 d采样1次,共采6次,每次采果约100个,品种内样品树混合采样,采收后常温下立即运回实验室。
1.2 试验方法
1.2.1 取样 剪取单果,用清水洗净后晾干,挑选大小均匀、成熟度一致、无病虫害及机械伤的果实。其中10个鲜果的果皮和果肉用于扫描电子显微镜观测样本的制定;30个鲜果于当天用作成熟度指标TSS含量的测定,以ATAGO PR-32α数显折光仪(日产)读取TSS含量。根据成熟期间果实TSS变化规律,选择完全成熟期和退糖后两个时期的样品进行电镜观察,即TSS含量最高和最后一次采果时。
1.2.2 测定项目及方法 成熟度按照韩冬梅等[6]方法确定,以比熟度表示,比熟度在95%~100%区段为果实的完全成熟阶段,之后进入退糖期,成熟前比熟度达95%时为始熟期,成熟后达95%时为始退期。计算比熟度和退糖速率:

1.2.3 电镜制样 参照黄旭明等[9]方法,用刀片将鲜果皮切成2 mm×2 mm大小片块,鲜果肉切成5 mm×3 mm大小块状,立即放入已预冷的4%戊二醛固定液(用pH 5.0柠檬酸-磷酸缓冲液配制、内含少量抗坏血酸)中,在2℃下固定1个月,每隔15 d更换一次固定液。制样时,用pH 5.0的磷酸缓冲液冲洗3次,每次15 min;经过乙醇梯度脱水(分别用50%、50%、60%、90%、100%乙醇溶液和100%加脱水剂的乙醇进行脱水,每个梯度脱水15 min);用醋酸异戊酯在通风橱内洗2次,每次洗15 min;临界点干燥,使用Eiko IB-5型离子溅射仪镀白金金膜,上台,在Philip XL30 ESEM环境扫描电子显微镜下观察拍照。
2 结果与分析
2.1 两个品种龙眼果实在成熟至退糖期间TSS含量和成熟度的变化
石硖和储良的采收日期、TSS含量、比熟度和退糖速率见表1。表1显示,石硖在7月25日、储良在8月1日分别进入成熟期,TSS含量分别为21.78%和19.88%,比熟度分别为96.39%和96.52%;两个品种的最高TSS含量分别为22.60%和20.60%,之后开始下降,当比熟度低于95%时,果实进入退糖期,开始衰老;石硖完全成熟期在7月25日至8月8日,储良在8月1~22日,分别历时14 d和21 d,退糖速率分别为每天0.60%和0.31%。可见,石硖较早熟、成熟期短、退糖速率高,储良较晚熟、成熟期长、退糖速率也慢。
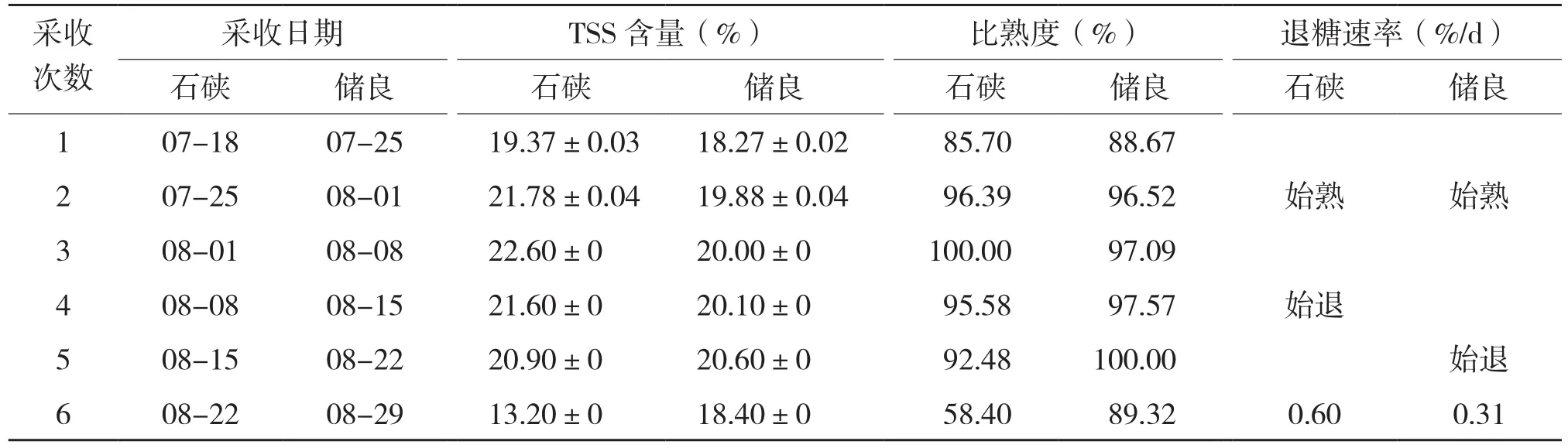
表1 两个品种龙眼果实在成熟至退糖期间TSS含量和成熟度的变化
2.2 龙眼果实在成熟至衰老期间的表观性状变化
挂树成熟期间,石硖和储良两个品种龙眼的果实内外表观性状均发生相似的规律性变化,具体表现为:成熟至完全成熟期间,果实有弹性,果形趋于固有形状;外果皮由青色转为青褐色或褐色,龟裂纹由尖锐粗糙变得平滑;内果皮呈白色或黄白色,表面光滑细腻,有光泽;果肉逐渐离核,种皮开始皱缩。完全成熟至退糖期间,果实弹性渐失,变得坚硬,果形逐渐不规则,外果皮趋向深褐色,内果皮出现褐斑且后期出现凸起的瘤状“果钉”;果核收缩明显,果蒂组织纤维化并突起、褐变。但不同品种间也存在差异:接近成熟后期,石硖果肉纤维化严重,口感变硬起“砂”,果实大小不一,形状也不规则;储良果肉虽然也出现少许纤维化,但内果皮“果钉”与褐变现象不如石硖严重,并仍保持光滑明亮。
2.3 成熟与退糖期间龙眼果皮显微结构变化
2.3.1 石硖龙眼果皮显微结构 成熟时,石硖果皮外表面为不规则平行细胞群,上覆不连续的较大块状蜡(图1A、C),带较多表皮毛;表面栓质层布满微裂缝并交错连接(图1C)。果皮分外果皮、中果皮和内果皮(图1B),外果皮的表皮细胞紧密平行排列,形状小,多呈长方形;中果皮主要由薄壁组织、维管束和石细胞组成[7],维管束细胞多位于接近内表皮的中下部位。中果皮接近外层的上部细胞为扁圆形,体积大,排列疏松,细胞间隙大,中部横向的维管束细胞具有较大的胞间隙和胞间腔;靠近内表皮的下部细胞较大,排列疏松不规则,为扁圆形或不规则四边形,呈海绵状结构(图1D)。内果皮由一层致密的薄壁细胞组成,内表面呈平行整齐的波浪状,附角质层和块状蜡,表面光滑,有光泽(图1E),即将起“钉”处有少数由细胞堆积形成的峰状突起,中间开裂(图1F)。

图1 成熟时石硖龙眼果皮的显微结构
退糖时,石硖果皮表皮毛脱落,蜡质层不明显,表面栓质层似风化状(图2A),有菌丝缠绕,沟壑变宽(图2B)。外果皮细胞明显木栓化,形态消失,胞间“裂缝”明显(图2C)。中果皮细胞腔变小,细胞壁变厚,也呈栓质化(图2D)。内果皮薄壁细胞整齐平行排列,有的细胞不断向内表面分生形成堆积状“果钉”(图2E),其与内果皮接合的底部细胞为平行排列的薄壁细胞,中部细胞形状和排列变得越不规则,个别细胞伸长呈管状,最顶端分生细胞形状再次变小,大小均匀(图2F),整个“果钉”顶部表面,均为平行排列、不断往上隆起的正方形或圆形细胞,有裂缝(图2G)。内果皮某些部位出现裂缝、孔洞,角质层破裂或缺失,表面粗糙,失去光泽(图2H)。

图2 退糖时石硖龙眼果皮的显微结构
2.3.2 储良龙眼果皮显微结构 成熟时,储良外果皮较平整,栓质组织碎裂成片状,分散在果表(图3A),蜡质层与表皮毛数目较少,有菌丝体缠绕(图3B)。外果皮薄壁细胞平行紧密排列(图3D)。中果皮细胞排列疏松,胞间隙较大,间以致密的块状栓质组织[10](图3C、F),其胞腔几乎消失,胞壁坚厚(图3E)。内果皮由一层长方形薄壁细胞紧密平行排列,其表面呈平行整齐的条带状(图3G),附致密的角质层和块状蜡,有光泽(图3H)。
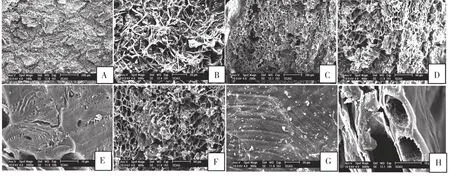
图3 成熟时储良龙眼果皮的显微结构
退糖时,储良果皮外表面疣状突起数目增多,蜡质脱落,微裂缝明显增多,表面更加粗糙(图4A),胞腔增大,呈空洞死细胞状,附薄层蜡状物(图4B、D)。中果皮细胞结构不清晰,胞间隙增大(图4C)。内果皮有“果钉”形成(图4E),由少量体积较大的薄壁细胞堆积形成峰状突起,无裂缝(图4F),底部为排列疏松无序的细胞群,胞腔较大(图4E);中上部细胞为纵向平行维管束结构(图4G);顶部为排列疏松的扁方形、无序堆叠的薄壁细胞(图4H)。

图4 退糖时储良龙眼果皮的显微结构
2.4 成熟与退糖期间龙眼果肉显微结构变化
2.4.1 石硖龙眼果肉显微结构 成熟时,石硖果肉表面波浪状,平滑(图5A),细胞呈长管状,饱满,平行排列,胞壁结构致密,较厚,胞腔大,切口平整,呈椭圆形或长椭圆形,刚性结构清晰(图5B)。退糖时,果肉外表面粗糙不平,皱褶状,无较大裂缝(图5C);细胞腔收缩,结构坍塌(图5D);胞壁分层,次生胞壁明显增厚(图5E),其表面出现裂缝(图5F)。

图5 成熟至退糖过程中石硖龙眼果肉的显微结构变化
2.4.2 储良龙眼果肉显微结构 成熟时,储良果肉表面乳状密集隆起,光滑(图6A),细胞丰满,呈正方形或长方形,整齐有序地平行排列(图6B);细胞腔比石硖小,狭长型,切口处胞壁向内弯曲,刚性较石硖弱(图6C)。退糖时,储良果肉表面细胞发生大块凹陷,呈鱼鳞状,现皱褶,多有大裂缝(图6D)。胞腔模糊,胞壁皱缩,结构坍塌,呈严重破坏状(图6E、F)。

图6 成熟至退糖过程中储良龙眼果肉的显微结构变化
3 结论与讨论
果皮是果实对抗病原物的第一道防线,与果实生理和抗病能力密切相关[10],果皮上的角质和蜡质层除了可以抑制水分蒸腾,更是阻止病原菌侵染的一道重要屏障[11-12],与果实成熟采收后的抗衰老能力密切相关。随着成熟进程的推进,果实衰老伴随而生,完全成熟和过熟后果实结构趋向分解,细胞结构趋向破坏,往往不耐贮运,因此选择合适的采收期非常重要[13-15]。退糖是龙眼果实挂树成熟至衰老期间的一大变化特征,对果实的适时采收和贮运性能影响很大,但品种之间也存在较大差异。帅良等[16]研究了35个品种龙眼果实成熟衰老期间的糖含量变化,发现TSS含量与总糖和甜度相关性极其显著,变化趋势基本一致,认为TSS含量的变化可以反映出果实退糖状况。本研究通过对石硖、储良两个品种的TSS含量变化分析,以比熟度划分成熟与退糖时间,认为石硖属于快退类型、储良属于极慢退类型,与帅良等[2]研究结果一致。
退糖快慢与两个品种的果实结构差异存在一定关系:(1)果皮结构差异主要在于中果皮,成熟时,石硖中果皮细胞排列紧密,细胞间隙小而少。退糖时,两个品种果皮结构差异不大。(2)果肉细胞结构差异较大,成熟时,石硖果肉细胞结构饱满,胞壁坚实,刚性较强;储良胞壁结构不及石硖挺实,切口胞壁向内卷曲。退糖时,石硖出现明显的细胞壁纤维化增厚现象,而储良不明显。(3)石硖退糖后内果皮着生“果钉”现象明显,数量也比储良多。可见,退糖后,两个品种果实结构的差异主要在于内果皮上“果钉”的发生和果肉的纤维化程度。“果钉”不断生长,插入果肉并汲取营养,并可能通过“果钉”中管状细胞,经由果蒂向果枝单方面输送,造成果实养分向树体的回流;果肉细胞的纤维化,可能也与糖分的转化有关,造成果肉中糖分迅速流失;而细胞器结构的破坏,除了导致果肉透水、透气和失重能力增加[6],其结构的破坏、质壁分离也会影响细胞壁代谢,如果胶物质代谢异常[17]、脂氧合酶活性增高[15]。另外,正常采收的果实在贮藏期间不会产生“果钉”,因此退糖后产生“果钉”也是挂树龙眼果皮衰老、果肉退糖的重要特征之一。石硖退糖时“果钉”数量较多,果肉纤维化明显,口感砂质化,TSS含量下降速率快,比储良表现出更明显的特异性衰老特征。
成熟时品种间果实结构差异影响其耐贮性,而退糖后的果实结构严重破坏,不利于长期贮藏。成熟时果皮组织结构致密,果肉细胞排列紧密、细胞壁厚实坚硬,有利于维持果实的贮藏性能[6],在同样栽培条件和同等采收成熟度的前提下,石硖的耐贮性比储良强[18-19]。退糖时的龙眼果实结构破坏严重,耐贮性大幅降低,不仅在于结构完整性遭到破坏,还在于退糖时随着糖含量尤其是双糖含量的大幅下降和单糖含量的微量上升[20],导致果实总糖含量显著降低,可能不足以为高呼吸水平的龙眼果实度过长期贮藏提供足够的能量底物,而退糖后的龙眼果实品种间耐贮性差异不大[6]。因此,根据品种特性,选择耐贮品种,及时、适时采收,有利于果实保持较好的品质和贮藏性能;过晚采收,既不利于保持商品价值,也不利于采后贮运,而研究有效的调控措施,延缓果实退糖速度,维持果实结构完整,从而延长果实挂树成熟期,既有利于延长采收期,又能够提升或保持果实的贮藏品质,这是龙眼生产中需要解决的难题之一,也是今后一个重要研究方向。
[1]Han D M,Li J G,Pan X W,et al.Recent advances in postharvest research on Longan fruit[J].Acta Hort,2014(1029):325-330.
[2]帅良,谷李桃,刘文浩,等.不同龙眼品种果实退糖特性分析[J].热带作物学报,2016,37(10):1900-1907.
[3]李建光,李荣,韩冬梅,等.古山二号龙眼采收成熟度研究[J].中国果树,2008(2):41-44.
[4]李建光,潘学文,李荣,等.广东储良龙眼果实的成熟特性研究[J].华南农业大学学报,2008,29(10):14-18.
[5]韩冬梅,李建光,潘学文,等.石硖龙眼果实成熟特性研究[J].广东农业科学,2007,34(11):69-72.
[6]韩冬梅,吴振先,李建光,等.龙眼果实采收成熟度与耐贮性关系研究[J].华南农业大学学报,2008,29(4):12-16.
[7]林河通,陈绍军,席屿芳,等.龙眼果皮微细结构的扫描电镜观察及其与果实耐贮性的关系[J].农业工程学报,2002,18(3):95-99.
[8]屈红霞,孙谷畴,蒋跃明.龙眼果皮显微结构与果实耐贮性关系探讨[J].武汉植物学研究,2001,19(1):83-86.
[9]黄旭明,袁炜群,王惠聪,等.抗裂性不同的荔枝品种果皮发育过程中钙的分布动态研究[J].园艺学报,2005,32(7):578-583.
[10]林河通,陈绍军,席屿芳.采收期对龙眼果实品质和耐贮性的影响[J].农业工程学报,2003,19(6):183-184.
[11]李雄彪,吴琦.植物细胞壁[M].北京:北京大学出版社,1993:126.
[12]胡宝忠,胡国宣.植物学[M].北京:中国农业出版社,2002:18.
[13]许娟.库尔勒香梨果实生长发育期及贮藏期生理生化特性研究[D].乌鲁木齐:新疆农业大学,2015.
[14]马玄,朱璇,赵亚婷,等.不同成熟度杏果实组织的结构变化与絮败关系[J].食品与发酵工业,2015,41(8):203-207.
[15]谢娟.红树莓果实生长发育规律及超显微结构研究[D].哈尔滨:东北农业大学,2011.
[16]帅良,钱盼红,刘文浩,等.不同龙眼品种果实成熟时糖含量及其特征研究[J].热带作物学报,2016,37(5):915-921.
[17]刘熙东.龙眼果实自溶及其与细胞壁代谢的关系[D].广州:华南农业大学,2005.
[18]韩冬梅,吴振先,陈维信,等.龙眼采后果肉生理生化变化研究[J].华南农业大学学报,2002,23(1):20-23.
[19]韩冬梅,杨武,牛佳佳,等.龙眼果实低温贮藏性能常规指标评价体系的构建[J].热带作物学报,2015,36(9):1685-1693.
[20]Shuai L,Li J,Niu J J,et al.Sucrosemetabolizing enzymes and their genes in the arils of two Dimocarpus longan cultivars[J].Biologia Plantarum,2016,60(4):741-748.
