高温胁迫对茄子果皮活性氧代谢、花青素及其主要合成酶的影响
2018-06-14吴雪霞张爱冬朱宗文查丁石
吴雪霞,张爱冬,朱宗文,姚 静,查丁石,李 贤*
(1.上海市农业科学院 园艺研究所,上海 201403;2.上海市设施园艺技术重点实验室,上海 201403;3.山东省临沂市农业局,山东 临沂 276001)
茄子(SolanummelongenaL.)是茄科重要蔬菜作物,在我国南北各地广泛种植。据统计,中国2014年茄子播种面积为80.6万hm2,总产量3068万t[1]。茄子属喜温作物,其生长发育的最适宜温度为22~30 ℃;当温度超过35 ℃时,茄子苗期植株的生长发育会受到抑制,坐果期落花落果,商品果果皮颜色变淡无光泽,产量和品质降低[2-4]。在高温胁迫下,植物体内的活性氧(ROS)清除系统被破坏,ROS大量产生,引起膜蛋白和膜内酯的变化,植物细胞受到破坏。植物体内启动抗氧化酶系统(SOD、POD、CAT和APX)以及渗透调节物质的变化,有效清除和抵御活性氧,以适应高温胁迫[5-6]。高温会导致花青素合成减少和分解增加。在花青素合成的上游途径中,温度主要对苯丙氨酸解氨酶(Phenylalanine ammonialyase, PAL)、查耳酮合成酶(Chalcone synthase, CHS)、查耳酮异构酶(CHI)等的活性进行调控;在下游途径中,主要通过对二氢黄酮还原酶(Dihydroflavonol 4-reductase, DFR)、花青素合成酶(Anthocyanidin synthase, ANS)和类黄酮3-葡糖基转移酶(Flavonoid glucosyltransferase, 3GT/UFGT)等的活性进行调控[7-8]。
目前高温胁迫对茄子的影响研究主要集中在茄子苗期耐热性鉴定[2,9]、生理指标变化等方面[4,10],而关于高温对茄子果实的影响较少报道。因此,本试验对茄子‘特旺达’的商品成熟果进行高温处理,测定研究了在果皮活性氧代谢过程中果皮花青素含量及花青素主要合成酶活性的变化规律,以期为明确高温对茄子果实、花青素含量的影响提供科学依据。
1 材料和方法
1.1 材料
供试茄子品种为‘特旺达’,由上海市农业科学院提供,在国内推广种植面积较大。
1.2 试验处理
选择大小、成熟度基本一致,无机械损伤的商业成熟的果实为研究对象。选出具有以上要求的商品成熟果的20株茄子植株,将它们随机分成2组,每组包含10株茄子,均置于植物生长气候室内。将其中一组作为对照,全天昼夜温度设置为(27±1)℃;另外一组做高温处理,全天温度设置为(45±1)℃;高温处理组与对照组除温度不同外,其他条件如光照和湿度等均一致。分别于处理后0、3、6、12、24和48 h取样,测定各项理化指标,重复3次。
1.3 测定指标与测定方法
1.3.1 茄子果皮活性氧及抗氧化酶活性的测定 丙二醛(MDA)含量、超氧阴离子(O2·-)产生速率,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)的活性,脯氨酸(Pro)和可溶性蛋白含量的测定方法均参照吴雪霞等[11]的方法;过氧化氢(H2O2)含量的测定参照郭欣欣等[12]的方法。
1.3.2 茄子果皮花青素含量及其主要合成酶活性的测定 称取0.2 g茄子果皮,加2 mL 25 mmol/L HEPES (pH 7.4)样品提取液,在冰浴下研磨成匀浆,在4 ℃下以12000 r/min离心20 min,上清液用于测定花青素含量,具体测定步骤参照苏州科铭生物技术有限公司(www.cominbio.com)的试剂盒说明书,每处理重复3次。
精确称取1 g茄子新鲜果皮,加入少量石英砂和等量PVPP,在液氮中将其研磨粉碎,分装到预冷的离心管中;再加入2 mL预冷的酶提取缓冲液,震荡漩涡混匀,在4 ℃下以12000 r/min离心20 min;将上清液(粗酶液)移至新离心管中,放置在冰浴中,用于酶活性测定;采用美国TSZ公司植物花青素酶(PAL、CHS、DFR、ANS、3GT)活性测定专用试剂盒进行检测。
1.4 统计分析
采用Origin软件绘图,用SPSS统计软件对平均数用Duncan’s新复极差法进行多重比较。
2 结果与分析
2.1 高温胁迫对茄子果皮MDA含量、O2·-产生速率和H2O2含量的影响
如图1所示,在对照条件下,茄子果皮MDA含量、O2·-产生速率和H2O2含量在0~48 h内基本保持不变;在高温胁迫下,茄子果皮MDA含量、O2·-产生速率和H2O2含量在处理后0~48 h内均呈上升趋势,在48 h时达到最大值,分别比对照增加了516%、128%和90%。

图1 高温胁迫对茄子果皮MDA含量、O2·-产生速率和H2O2含量的影响
2.2 高温胁迫对茄子果皮抗氧化酶活性的影响
图2表明:对照茄子果皮的SOD和APX活性在0~48 h内呈逐渐上升的趋势,POD和CAT活性在0~48 h内均未发生明显变化;在高温处理下,茄子果皮4种酶的活性随处理时间的延长均呈先上升后下降的趋势,其中SOD活性在处理12 h时达到最大值,比对照增加了70.05%,POD、CAT和APX活性在处理24 h时达到最大值,分别比对照增加了337.75%、279.38%和42.79%。
2.3 高温胁迫对茄子果皮脯氨酸和可溶性蛋白质含量的影响
从图3可以看出:茄子果皮脯氨酸和可溶性蛋白质含量的变化趋势一致,其中对照组在0~48 h内变化幅度均较小,高温处理组在0~48 h内均呈直线上升的趋势,在处理48 h时分别为对照的7.05和0.75倍。
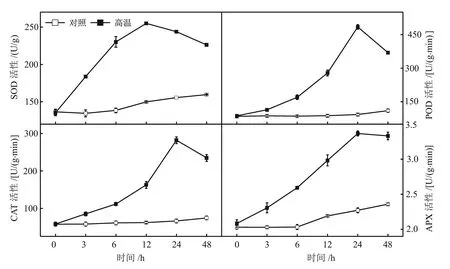
图2 高温胁迫对茄子果皮SOD、POD、CAT和APX活性的影响

图3 高温胁迫对茄子果皮脯氨酸和可溶性蛋白含量的影响
2.4 高温胁迫对茄子果皮花青素及其主要合成酶活性的影响
由图4可见:在对照下,茄子果皮花青素(ANTH)含量及其主要合成酶(CHS、DFR、ANS和3GT)的活性在0~48 h内基本保持恒定;在高温处理下,以上指标在0~48 h内均呈持续下降的趋势,在处理48 h时,ANTH、CHS、DFR、ANS和3GT分别比对照降低了31.60%、39.91%、35.10%、27.29%和30.35%;PAL的活性在高温处理下呈先上升后降低的趋势,在处理12 h时达到峰值,比对照增加了50.88%。
3 讨论
据金春燕等[13]报道,高温处理导致番茄体内MDA含量和O2·-产生速率均显著增加;张建光等[14]研究表明,高温和强光能促使苹果果实表皮组织内MDA含量和O2·-产生速率大幅增加,使细胞膜的结构和功能受损,从而引起高温和强光伤害。本试验结果表明,高温胁迫导致茄子果皮MDA含量、O2·-产生速率和H2O2含量增加,且在胁迫过程中均呈持续上升的趋势,说明高温胁迫造成了茄子果皮的过氧化伤害,且这种伤害随胁迫时间的延长而加重。
SOD、POD、CAT和APX是酶促防御系统的重要组成成分,SOD催化O2·-生成H2O2和O2,POD、CAT和APX负责将H2O2分解为H2O和O2,以提高植物对高温胁迫的适应性[15-16]。李建建等[17]研究认为,高温胁迫使黄瓜幼苗叶片的SOD活性呈下降趋势,而POD和CAT的活性持续升高。徐佳宁等[18]研究表明,在40 ℃高温处理下,番茄幼苗叶片中SOD、POD、CAT和APX等抗氧化酶的活性呈先上升后下降的趋势,但不同番茄品种的峰值出现时间不同。本研究结果表明,在高温胁迫过程中茄子果皮的SOD、POD、CAT和APX活性均呈先上升后下降的趋势。在高温胁迫前期抗氧化酶活性的升高可能是茄子果实对短时高温胁迫的一种应激和自我适应性反应,通过调节抗氧化酶活性试图建立ROS产生与清除的平衡关系;随着胁迫时间的延长,抗氧化酶受抑程度超出了茄子果皮自身的适应能力,因此,抗氧化酶活性降低,膜脂过氧化程度加重。

图4 高温胁迫对茄子果皮花青素及其主要合成酶活性的影响
脯氨酸和可溶性蛋白是植物抗逆系统中极为重要的渗透调节物质,具有调节渗透压及保护细胞膜结构稳定的作用。在本试验中,高温胁迫促进了茄子果皮脯氨酸和可溶性蛋白的积累,且随着高温胁迫时间的延长呈持续增加趋势。这与李建建等[17]的研究结果类似,可能是因为脯氨酸和可溶性蛋白的积累提高了植株的渗透调节能力,调节了细胞的渗透平衡,维持了细胞结构的稳定,以适应高温胁迫逆境。
高温处理使植物花青素合成减少和分解增加,甚至表现出褪色或较难着色[7-8]。高温可通过诱导植物体内花青素苷合成途径相关基因的表达来调控花青素苷的呈色反应[7,19]。魏海荣等[20]对甜樱桃花芽的研究结果表明,高温促进PAL活性上升,低温降低PAL活性。Caldwell等[21]对大豆的研究结果显示,当温度从18 ℃上升到28 ℃时,异黄酮含量降低,可能是其合成途径中CHI或CHS的活性降低。Mori等[8]研究表明,夜间高温会抑制UFGT的表达,降低酶活性,从而导致植物体内花青素积累减少。本试验结果表明,茄子果皮花青素含量随着高温处理时间的延长而逐渐降低,且花青素主要合成酶PAL的活性增加,而CHS、DFR、ANS和3GT的活性下降。这与前人的研究结果一致。
参考文献:
[1] 杨旭,王露,张宇,等.茄子种质资源苗期耐涝性与SSR标记的关联分析[J].分子植物育种,2017,15(11):4635-4641.
[2] 李植良,孙保娟,黎振兴.高温胁迫下茄子的耐热性表现及其鉴定指标的筛选[J].植物遗传资源学报,2009,10(2):244-248.
[3] 孙保娟,黎振兴,罗少波,等.持续高温胁迫对茄子幼苗几个主要生理指标的影响[J].热带作物学报,2010,31(9):1528-1534.
[4] 李艳艳,王俊青,李植良,等.短期高温胁迫对茄子主要生理指标及叶片解剖结构的影响[J].热带作物学报,2016,37(9):1774-1780.
[5] Bartwal A, Mall R, Lohani P, et al. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses [J]. J Plant Growth Regul, 2012, 32: 216-232.
[6] Janeczko A, Oklestkova J, Pociecha E, et al. Physiological effects and transport of 24-epibrassinolide in heat-stressed barley [J]. Acta Physiol Plant, 2011, 33: 1249-1259.
[7] Huh E J, Shin H K, Choi S Y, et al. Thermo susceptible developmental stage in anthocyanin accumulation and color response to high temperature in red chrysanthemum cultivars [J]. Korean Journal of Horticultural Science & Technology, 2008(6): 357-361.
[8] Mori K, Goto-Yamamoto N, Kitayama M, et al. Loss of anthocyanins in red-wine grape under high temperature [J]. Journal of Experimental Biology, 2007, 58: 1935-1945.
[9] 李涛,李植良,夏碧波,等.茄子创新资源的耐热及抗青枯病评价研究[J].中国农学通报,2017,33(36):72-77.
[10] 孙保娟,李植良,黎振兴,等.茄子耐热性苗期鉴定研究[J].广东农业科学,2007(2):27-29.
[11] 吴雪霞,朱月林,朱为民,等.外源一氧化氮对NaCl胁迫下番茄幼苗生理影响[J].中国农业科学,2006,39(3):575-581.
[12] 郭欣欣,李晓锋,朱红芳,等.淹水胁迫对不结球白菜抗坏血酸-谷胱甘肽循环的影响[J].植物生理学报,2015,51(12):2181-2187.
[13] 金春燕,郭世荣,施标,等.高温对不同番茄品种幼苗叶片抗氧化系统和渗透调节物质的影响[J].上海农业学报,2011,27(3):26-30.
[14] 张建光,陈少春,李英丽,等.高温强光胁迫对苹果果皮PPO活性的影响[J].生态学报,2008,28(10):4645-4651.
[15] Fang H H, Jing T, Liu Z Q, et al. Hydrogen sulfide interacts with calcium signaling to enhance the chromium tolerance inSetariaitalic[J]. Cell Calcium, 2014, 56: 472-481.
[16] Sun N, Wu Q, Liu Y Y, et al. Hydrogen-rich water reestablishes ROS homeostasis but exerts differential effects on anthocyanin synthesis in two varieties of radish sprouts under UV-A irradiation [J]. Journal of Agricultural and Food Chemistry, 2014, 62: 6454-6462.
[17] 李建建,郁继华,常雅君,等.高温胁迫对黄瓜幼苗叶片质膜透性及保护酶活性的影响[J].长江蔬菜,2007(9):59-61.
[18] 徐佳宁,刘钢,张利云,等.高温胁迫对不同番茄品种叶片抗氧化系统的影响[J].山东农业科学,2016(10):27-31.
[19] Shvarts M, Borochov A, Weiss D. Low temperature enhances petunia flower pigmentation and induces chalcone synthase gene expression [J]. Physiologia Plantarum,1997(99): 67-72.
[20] 魏海荣,韩红霞,刘庆忠,等.温度对甜樱桃花芽酚类物质含量和相关酶活性及休眠的影响[J].果树学报,2007,24(1):38-42.
[21] Caldwell C R, Brits S J, Mirecki R M. Effect of temperature, elevated carbon dioxide, and drought during seed development on the isoflavone content of dwarf soybean [Glycinemax(L.) Merrill] grown in controlled environments [J]. Journal of Agricultural and Food Chemistry, 2005, 53(4): 1125-1129.
