植物非整倍体研究进展
2018-05-30朱斌田贵福贺路英李再云
朱斌 田贵福 贺路英 李再云
摘 要:非整倍体(aneuploid)是指相对于正常个体(euploid)的染色體组增加、减少一条或若干条染色体的生物个体。由于非整倍体个体存在基因剂量效应的不平衡性(gene-dosage imbalance),非整倍体个体往往会表现严重的表型缺陷(aneuploid syndrom),如发育迟缓,个体矮小,难以繁殖后代等。在人类中,最为典型的例子为导致新生儿智力缺陷的唐氏综合症,由额外的一个21号染色体拷贝(部分拷贝)引起。此外,大多数癌细胞类型表型为严重的非整倍体。在大多情况下,非整倍体对于动物及人类是致命的,而植物对于非整倍体则往往表现出较强的耐受力,特别是在异源多倍体植物中。植物非整倍体对于植物的遗传、育种研究有重要意义,在基因及分子标记的物理位置确定,基因转移,连锁群与染色体的对应关系的确立上具有无可比拟的优势。该文综述了近些年来有关植物非整倍体研究的结果,介绍了非整倍体的几种重要成因和有关非整倍体鉴定手段的变迁,阐述了植物非整倍体对个体表型、基因表达以及表观遗传方面的影响,重点讨论了非整倍体在植物进化、基因组序列测定以及遗传改良方面的潜在作用。同时,探讨了植物非整倍体研究的新思路,以及利用非整倍体促进相关植物遗传改良、育种研究的新方法。
关键词:非整倍体,剂量效应,高通测序,遗传改良,基因重组,表观遗传
中图分类号:
文献标识码:A
文章编号:1000-3142(2018)10-1404-07
Abstract:The aneuploidy for one species refers to the occurrence of one or more extra or subtractive chromosomes or chromosomal segments in the normal chromosome number in the somatic cells. Because of gene-dosage imbalance,compared to normal organisms,aneuploidy usually gives rise to severe defective phenotypes,such as delayed development and growth,stunted individual architecture,barrier of reproduction due to severe reducedfertility and so on. In human,the most serious aneuploidy syndrome is Downs syndrome with irreversible cognitive impairment,muscle hypotonia,as well as dysmorphic features in newborn,caused by one extra copy of the whole or partialchromosome 21. And the majority of solid tumor cells show different complex level aneuploidy. It is long recognized that plants have better tolerance to the impact of aneuploidy than animals,particularly in allopolyploid plants. As rare but vital germplasm resources,aneuploidy in crops is essential and powerful for these genetic improvements and advanced breeding studies which are focusing on confirmation of the physical location of valuable genes and assistant markers,and transferring of desirable genes from relative species in cultivars. Aneuploidies are also taken as convenient materials to identify the relationship between chromosomes in cytology and corresponding linkage groups in genetic map. In this review,we provided a concise but comprehensive review of recent studies in aneuploidy plant,in which we introduced some important mechanisms responsible for the formation of aneuploidies,the transition of identification methods for some typical aneuploidies,detrimental effects on organisms and cells caused by unbalanced chromosome,and gene expression change and epigenetic alteration of aneuploidy compared to normal organisms. And we particularly discussed some potential values of aneuploidies for the research of genome evolution in plant,and introduced important function of aneuploidy in allopolyploid crops to improve their genome sequencing and genetic improvement. Meanwhile,we sought some novel ideas and methods in the aneuploidy reseach,and also discussed the possibility of some new insights intogenetic improvement and advanced breeding with the aid of aneuploidy in some crops.
Key words:aneuploidy,gene-dosage,high throughput technology,genetic improvement,genetic recombination,epigenetic
相对于整倍体生物(euploid)而言,非整倍体(aneuploid)指的是个体的染色体数目不是成倍的增加或者减少,而是单个或几个的增添或减少,进而导致个体不平衡的染色体组成。染色体数目增加的个体被称作超倍体(hyperploidy),染色体数目减少的个体被称作亚倍体(hypoploidy),常见的亚倍体主要有单体(monosomic),即相对于二倍体个体染色体组(2n)缺少一条染色体的个体(2n-1);缺体(nullisomic),即缺少一对同源染色体的个体(2n-2);超倍体有三体(trisomic),即增加一条染色体的个体(2n+1);四体(tetrasomic),即增加一对同源染色体的个体(2n+2)。相较于超倍体,亚倍体对个体造成更为严重的影响,因而不容易获得。此外,在异源多倍体植物中,出现高频率的隐性非整倍体(recessive aneuploid),即个体染色体数目与正常个体相同,但一条(对)染色体被其部分同源染色体所替代,保证染色体组水平上的剂量平衡。在人类中,最为典型的例子为导致新生儿智力缺陷的唐氏综合症,由额外的一个21号染色体拷贝(部分拷贝)引起。此外,大多数的癌细胞类型表型为严重的非整倍体。在动物中,除了部分性染色体的非整倍体类型,其它类型非整倍体都会对个体造成不可逆的影响。相较于动物,植物对非整倍体表现出较高的耐受力,且对于遗传育种研究者而言,植物非整倍体是不可多得的遗传及种质资源(Siegel & Amon,2012),多被用来定位基因或标记的物理位置,确定连锁群及染色体的对应关系,以及转移有利基因等。
1 非整倍体产生的途径及鉴定方法
非整倍体的产生途径主要是细胞有丝或减数分裂过程中染色体异常分离导致的,染色体在有丝分裂时期的异常分离往往导致嵌合型的非整倍体,若发生在减数分裂时期就有可能产生非整倍体配子,进而造成整个有机体的非整倍性。
有机体细胞分裂过程中,正常的染色体是由纺锤体产生的纺锤丝牵引姐妹染色单体拉向两极而进行分离的,此过程中,纺锤丝首先要结合到染色体上的纺锤丝组装位点上(spindle-assembly checkpoint,SAC)(Musacchio & Salmon,2007),如果细胞此时挟带了不正常的SAC或者SAC的靶標后期促进复合物(APC/C-Cdc20)超表达,染色体分离就会出现异常,进而产生非整倍性的细胞(配子);姐妹染色单体聚合蛋白(cohesion)的缺陷以及分离酶的超活性也会造成染色体的异常分离从而产生非整倍体细胞(Musacchio & Salmon,2007; Ganem et al,2009);着丝粒微管的畸变也能导致非整体,而且是主要来源;染色体异常分离,多极纺锤体等均能产生非整倍体细胞(配子)(Siegel & Amon 2012; Ganem et al,2009)。对于植物而言,由于存在倍性的变异,不同倍性间杂交或回交可产生大量的非整倍体个体,甚至是全套的单/缺体(Sears,1954; Chen et al,2007; Zhu et al,2015)。此外,植物进行远缘杂交的过程中由于存在外源基因组的(genomic shock)冲击效果,也能形成一些非预期的非整倍体,这种现象在异源四倍体油菜与近缘种的杂交中比较普遍(Hua & Li,2006; Tu et al,2010; Zhu et al,2016; 朱斌等,2017)。
植物非整倍体在遗传、育种方面的应用往往受限于其类型的鉴别。基于个体染色体组的核型分析是最为常见的鉴定方法。通常来说某一物种的染色体核型是稳定的,不同的染色体在长度、臂比、异染色质区,以及随体有无方面存在差异,因此根据这些特征可将染色体区别开来,进而确定个体非整倍体类型(Young et al,2012)。在此基础上已陆续发展出C带法、G带法、基于染色体特异探针的荧光原位杂交技术(FISH)来进行个体核型分析(Duan et al,2015; Jiang et al,2016; Zhang et al,2017)。由于FISH具有较强的可靠性和直观性,可鉴别较小的染色体变异等优点,因而被广泛应用到动植物的核型分析及植物非整倍体鉴别中(Mestiri et al,2010; Xiong et al,2011; Zhang et al,2013; Zhu et al,2016)。流式细胞术也是近年来常用的检测非整倍体类型的手段(Pavlíková z et al,2017),但往往对于基因组较大的物种有效。此外,基于染色体序列开发的特异性分子标记由于具有通量高、易操作等优点,也成为鉴定非整倍体的有效手段(Kang et al,2014; Chen et al,2014; Zhu et al,2016),但这些标记往往都是显性标记,不能有效鉴别出染色体的拷贝数,通常需要辅助普通细胞学的手段(Zhu et al,2016)。随着高通量技术的发展,基于信使RNA(mRNA)测序技术日趋成熟,由于多数基因的表达与其拷贝数存在线性关系,因此基于全基因组RNA测序技术可以准确确定某一个体的染色体组,因而该方法被陆续应用到非整倍体个体的鉴定中(Letourneau et al,2014; Zhu et al,2015; Zhang et al,2017)。
2 非整倍性对有机体的影响
基于二倍体生物的非整倍体个体研究表明,非整倍体往往在早期发育过程就会死亡,即使存活下来的个体通常也表现持续的发育缺陷。20世纪初,Boveri在以两个海胆(Sea urchin)精细胞与一个卵细胞受精发育的幼胚中观察到了这种致死或发育缺陷现象(Zhu et al,2015)。Torres et al(2007)对包含一整套酿酒酵母(Saccharomyces cerevisiae)基因组的附加一条或一对及更多染色体的材料进行生理生化、基因表达及蛋白水平的系统分析,结果证实了这些非整倍体系存在细胞增殖缺陷。对于玉米(Zea mays)、水稻(Oryza sativa)、油菜(Brassica napus)及拟南芥(Arabidopsis thaliana)的非整倍体研究也证实了植物非整倍体同样存在这种生长缺陷(McClintock,1929; Singh et al,1996; Henry et al,2010;Chen et al,2007)。而相对于动物而言,植物对于非整倍体的影响要有更好的适应性,这可能是由于同源基因或部分同源基因的剂量补偿导致的(Siegel & Amon,2012)。
非整倍体性不仅影响个体的基因表达,同样造成了个体表观遗传的变化,且这种变化是持久的、可遗传的(Letourneau et al,2014; Henry et al,2010; Gao et al,2016)。Henry et al(2010)在拟南芥非整倍体自交后代中检测到染色体组正常的个体,其同样也表现非整倍体母本的一些性状,说明这些持久的影响可能来自表观遗传的变化,随后这一假说在小麦非整倍体涉及全基因组甲基化的表观遗传研究中加以证实(Gao et al,2016)。
3 基于非整倍体的基因平衡假说
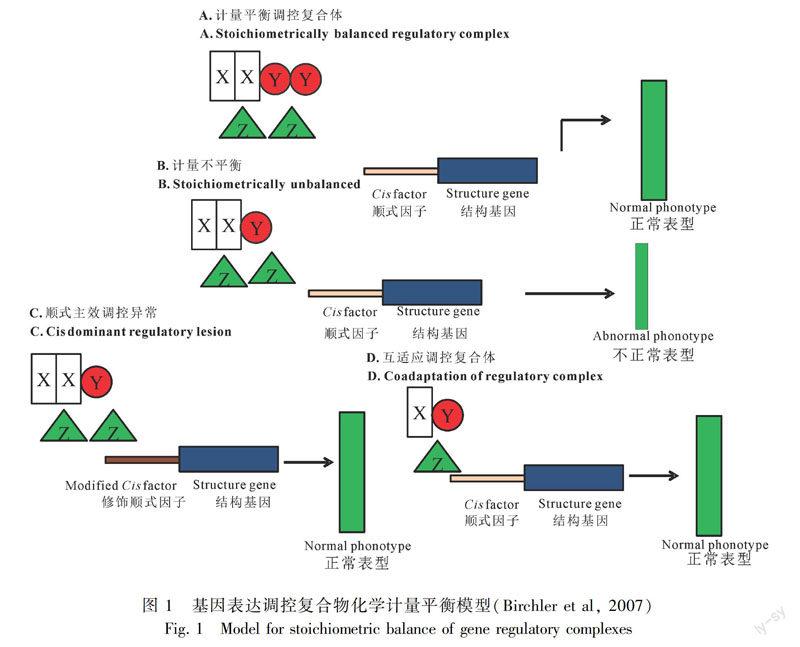
早在20世纪30年代,研究者就发现曼陀罗(Datura stramonium)的三体植株显现比三倍体(triploid)植株更为严重的表型缺陷,随后对于玉米的倍性变化及非整倍体个体材料的系统研究证实了这一结果(Guo et al,1996)。基于这些结果,Birchler et al(2001)提出了基因组平衡假说(gene/genome balance hypothesis),他认为在非整倍体个体中,一套或部分染色体编码的对于剂量敏感的基因产物,其严格的化学计量平衡被打破,从而导致了严重的表型及基因表达变化(图1,引自Birchler et al,2007)。随后,基因平衡假说在拟南芥、玉米、果蝇(Drosophila melanogaster)、甘蓝型油菜、以及普通小麦(Triticum aestivum)非整倍体个体涉及部分或者全基因基因表达的相关研究中被逐渐证实(Huettel et al,2008; Guo et al,1996; Malone et al,2012; Makarevitch & Harris,2010; Zhang et al,2013; Zhu et al,2015; Zhang et al,2017)。
4 植物非整倍体在遗传研究中的作用
4.1 利用植物非整倍体定位相关基因
在植物及低等动物中,非整倍体对发育的影响相对较小,且植物存在基因表达的剂量补偿机制,因此很多非整倍体类型得以存活下来,这为相关植物的遗传分析及改良研究提供了难得的材料。植物非整倍体可以方便基因及分子标记的物理定位,有利于目标基因的转移,确定连锁群与染色体的对应关系等(轩淑欣等,2002)。小麦次级代谢产物异羟肟酸能有效防御真菌病害的侵袭,Niemeyer & Jerez(1997)通过测定小麦的一系列单体及代换系,将异羟肟酸合成基因定位到小麦4A和4B染色体上。兰登硬粒小麦(langdon durum)携带有优异的抗叶锈病基因,Hussein et al(2005)通过对四倍体硬粒小麦(Triticum turgidum,AABB)D染色体组代换系抗叶锈病的研究,将硬粒小麦抗叶锈病基因定位到了6B和4A染色体上。对于拟南芥多个非整倍体个体的研究(Henry et al,2010)证实不同染色体对于表型的相应贡献,而对甘蓝型油菜缺失一对C2染色体的非整倍体材料的基因表达分析,确定了C2染色体携带有控制开花的主效QTLs(Zhu et al,2015)。Geleta et al(2012)通过对全套的白菜-甘蓝单体附加系进行PCR扩增分析,将64对特异SSR引物锚定到不同的C染色体上,这些特异引物极大的方便甘蓝及甘蓝型油菜的遗传图谱构建。需要指出的是,在异源多倍体植物中,由于部分同源染色体的补偿机制,一些不常见的单体(monosomics)、缺体(disomics)被完整的建立起来,例如在异源六倍体小麦所发现的一整套单体及缺体(Sears,1954)材料,对于这些材料的遗传学研究使得我们更为清楚地了解了小麦的基因组结构,部分同源染色体的相互关系。最近,基于一套小麦的单体材料,采用chromosome-base的测序方法成功组装了六倍体小麦高达17 G的基因组信息(international wheat genome sequencing consortium,IWGSC,2014)。
4.2 植物非整倍体参与异源多倍体植物的进化史
多倍化(polyploidization)过程被认为是植物进化历程中的一个主要驱动力。据研究,约80%的开花植物经历过或正在经历至少一次的多倍化过程(Van de Peer et al,2009)。其中经不同物种杂交及后续基因组加倍形成的异源多倍体拓宽了物种间的遗传基础,创造了更为丰富的遗传多样性,在生长势、产量、适应性等方面远远超过了二倍体亲本(Chen,2010)。近几年,研究者多以人工合成的或者自然界刚形成的异源多倍体植物模拟、演化植物异源多倍体的进化历程,值得注意的是,在人工合成或自然界刚形成的异源多倍体后代中普遍存在非整倍体或隐形非整倍體个体(Mestiri et al,2010; Xiong et al,2011; Zhang et al,2013; Pavlíková et al,2017),而且在人工合成的异源四倍体甘蓝型油菜(AACC),六倍体小麦(AABBDD)的高世代后代中非整倍体个体依然有较高的比例(Mestiri et al,2010; Xiong et al,2011; Zhang et al,2013)。其中,对来自16个系的1 000余株人工合成的六倍体小麦核型的研究发现(Zhang et al,2013),非整倍体植株覆盖了每个株系20%~100%的植株,同时还证实了B基因组染色体最容易丢失或增加,A基因组染色体次之,而新加入的C基因组染色体最为稳定。婆罗门参属Tragopogon miscellus是自然界最近形成的一种异源多倍体植物,起源于北美洲,由20世纪早期从欧洲引入的二倍体植物Tragopogon dubius 和 Tragopogon pratensis(2n=12)经自然杂交、染色体组加倍形成,大约仅有80 a的进化历史(约40代)。Chester et al(2012)以基因组分析以及原位杂交的技术手段对该物种的不同群体进行核型分析,发现没有任何一个群体有固定的核型组成,69%的植株表现一条或者多条染色体的非整倍性,其中高达85%的植株是隐性非整倍体(hidden aneuploid),即拥有整倍体个体的染色体数目,但染色体组成往往存在易位、部分同源补偿等。广泛的部分同源补偿在人工合成的甘蓝型油菜、六倍体小麦中也被证实,可能预示着基因剂量效应是非整倍体选择性保留的主要约束机制(Xiong et al,2011; Zhang et al,2013)。
这些结果无疑说明非整倍体过程在异源多倍体起源、进化历程中是持久的、广泛的。这一过程对于异源多倍体的进化可能是必须的,这将有助于新物种快速地产生大量的遗传变异,从而适应环境。部分同源染色体间的补偿机制也将有助于异源多倍体快速进行二倍体化。这种在新合成的异源多倍体植物中广泛存在的非整倍体,目前尚无系统的解释,猜测可能是由于亲本间遗传物质的不平衡造成的,如着丝粒组蛋白间的差异等。
4.3 植物非整倍体诱导染色体重组
变异和重组被认为是产生遗传多样性的两个基本来源,而重组又是有性生殖个体产生遗传多样性的主要机制,其频率远高于基因突变。近些年的研究发现,非整倍性可以大幅提高个体的重组频率。Nicolas et al(2009)发现甘蓝型油菜与白菜杂交形成的倍半二倍体(AAC)其同源重组的频率要远高于四倍体甘蓝型油菜,证明额外的C基因组会影响A基因组的重组,Suay et al(2014)以倍半二倍体杂种与白菜再次杂交,获得一系列的附加不同数目的C染色体非整倍体植株,对这些非整倍体植株的遗传分析发现不同的C组染色体及单拷贝染色体的数目对诱导同源重组的频率有差异。甘蓝型油菜与其近缘种的远缘杂交往往会形成仅包含甘蓝型油菜染色体组的非整倍体植株,Zhang et al(2013)对甘蓝型油菜与诸葛菜、荠菜杂交后,经多代回交得到的染色体数目为38的新分离甘蓝型油菜进行分子标记分析时发现,与原始亲本相比分离得到的甘蓝型油菜在基因组、表观水平有很大差异。这些研究证实了非整倍性个体可以诱导本身基因组产生高频率的重组,这有利于产生遗传多样性,可为育种提供材料基础,但这是否是非整倍体的共性特征,還需要在其他的物种的非整倍体个体中加以验证。
5 展望
随着高通量技术的发展,以基因组、转录组、蛋白质组、Chip-seq等组学的研究手段有望对植物非整倍体个体进行系统的阐释,深入了解非整倍体细胞及个体的成因,揭示其影响表型、基因表达、表观遗传的机理。近十年来,通过对非整倍体全基因组基因表达研究证实非整倍体基因异常表达不仅存在于缺失或增加的染色体上(顺式作用),而且广泛分布于拷贝数未改变的染色体上(反式作用),这一现象被认为与非整倍体的表型变化密切相关(Huettel et al,2008; Letourneau et al,2014; Zhu et al,2015)。此外,一些异源多倍体的非整倍体大大降低了植物基因组的结构复杂性,利用高通量技术,可以揭示一些复杂异源多倍体的基因组信息以及基因互作模式(IWGSC,2014; Zhu et al,2016)。同时,对于非整倍体的深入研究将有助于揭示基因间的互作关系。此外,人类的一些遗传疾病被证实与非整倍体相关(例如唐氏综合症),近些年的研究也证实大多数的癌变细胞与非整倍体存在因果关系,对植物非整倍体遗传机理的揭示有望为人类最终治愈非整倍体疾病提供可行的思路。
