高寒草甸湿润与干旱化生境中植物多样性的海拔分布格局
2018-05-28张大才李双智
张大才,李双智, 张 锐
(西南林业大学林学院,云南 昆明 650224)
物种多样性空间分布格局是生物多样性保护和生物地理单元划分的基础,一直受到高度关注[1-2]。生境受到外界干扰后,物种组成和多样性也随之改变[3],生境变化是否影响物种多样性的分布格局?高寒草甸是青藏高原分布面积最大的植被类型之一,受全球变化和外界干扰严重[4-5],生境趋向干旱化,群落退化演替,物种多样性下降[6]。藏东南高寒草甸在生境干旱化过程中,植物群落仍然以嵩草属(Kobresia)植物为优势种,根据土壤水分条件可分为湿润与干旱化两种生境类型[7]。生境类型与物种多样性密切相关,生境的变化都将引起植物群落类型和物种多样性的变化,如不同坡向高寒草甸物种多样性不同[8],林中空地、沟谷、湿地等不同生境物种多样性存在差异[9]。但是,更多研究都主要比较不同生境类型间物种多样性的差异,而很少分析不同生境物种多样性分布格局的差异,本研究将比较藏东南高寒草甸湿润与干旱化生境植物多样性沿海拔梯度分布格局的差异。
在外界干扰过程中,高寒草甸群落结构发生了很大的改变[10-12],一些物种的个体形态趋向小型化[13],植物体构件数量明显下降[14],在群落中的分布频度低而成为群落中的偶见种(occasional species)。同时,植物群落受到外界干扰后,改变了群落物种组成和种间关系,为其他物种侵入群落提供了机会[15],这些侵入性物种也可能是分布频度低的偶见种。藏东南高寒草甸受放牧干扰严重,群落中偶见种对物种多样性组成及海拔分布格局有何影响?本研究将植物群落中的物种区分为偶见种和常见种,比较其多样性沿海拔梯度的分布格局。
α多样性和γ多样性统称编目多样性,测度物种组成多样性,但二者测度的尺度不同,α多样性测度局域尺度上的物种多样性,而γ多样性测度区域尺度上的物种多样性[16]。β多样性测度环境梯度上物种组成的变化,其值对α和γ多样性起关键作用。β多样性的测度方法有多种,如倍性分配法、加性分配法、物种组成相似性等,然而众多方法的测度结果不完全一致[17-18]。β多样性概念的提出是为了反映物种组成沿环境梯度的变化[19],物种的分布频度必将影响β多样性,分布频度低的偶见种对β多样性的贡献更高[20-22]。γ多样性在更大空间尺度上测度群落物种多样性,与α和β多样性的分布格局是否具有一致性?因此,本研究主要研究目的有:1)分析高寒草甸植物群落种子植物多样性沿海拔梯度的分布格局;2)比较高寒草甸湿润与干旱化生境中物种多样性沿海拔梯度分布格局的差异;3)分析偶见种对物种多样性组成的贡献。
1 材料与方法
1.1 研究地概况

1.2 样地设置与群落调查
东达山高寒草甸以嵩草属植物为优势种,分布于林线与高山流石滩之间,分布的海拔范围近1 000 m。从林线边缘的高寒草甸开始设置样地,海拔每升高约100 m设置一个样地,直至高山流石滩边缘,共9个样地,除1、2号样地外,其余样地坡度相差不大(表1)。在每个样地设置10个1 m×1 m样方,其中第1-5号为湿润生境样方,设置于沟谷溪边地表稍有积水或土壤表面湿润的生境中;第6-10号样方为干旱化生境(干旱生境)样方,均设置于北坡,地表干燥;第10号样方生境极其干旱,为嵩草属植物的分布边缘,样方外为高山灌丛。调查样方内的物种组成、盖度和多度等调查因子。
1.3 数据分析
1.3.1偶见种的确定 偶见种指在群落中出现频度低的物种,从研究的实际出发,有多种确定方法,如将出现在所有样地的物种确定为常见种,反之则为偶见种[24];或者根据物种在群落中出现的频度确定,将频度低于一定标准的物种确定为偶见种[21-22,25];或者根据研究者的经验判断,将个体数量少的物种确定为偶见种[26]。本研究根据物种在样方中出现的频度确定群落中的偶见种,将频度低于群落中物种平均频度的物种作为偶见种。湿润和干旱生境中物种在样方中出现的平均频度分别为6.8和7.1,因此本研究中将频度≤7的物种作为偶见种,将频度>7的物种作为常见种,分析所有种、偶见种和常见种多样性沿海拔梯度的分布格局。

1.3.3数据统计分析 采用SPSS 16.0进行回归分析,分析物种多样性与海拔梯度之间的回归关系,并绘制回归关系图。使用ANOVA方差分析比较湿润与干旱两种生境中物种多样性的差异,用离散系数(coefficient of variance,CV)衡量同一样地不同样方间物种丰富度分布的均匀程度,CV值越大表示分布越不均匀,CV值为0则表明各样方物种丰富度相同,分布最均匀,CV=标准差/平均值。
2 结果
东达山90个样方中共记录种子植物160种,其中湿润生境132种,干旱生境107种,两种生境物种组成Simpson相似性系数为0.736。湿润生境中有偶见种71种,占湿润生境物种总数的68.9%;干旱生境中有偶见种75种,占干旱生境物种总数的70.1%。
2.1 物种丰富度沿海拔梯度的变化
湿润生境中各样方所有种、常见种丰富度显著高于干旱生境(P<0.05),而偶见种丰富度在两种生境中的差异不显著(P>0.05)(表2)。湿润生境中,所有种和偶见种丰富度随海拔的升高而呈显著线性下降(P<0.05),而常见种丰富度随海拔的增加略呈增加趋势(P>0.05)(图1)。干旱生境中,所有种、偶见种和常见种丰富度随海拔的增加均呈现出增加趋势(P>0.05)(图1)。常见种丰富度在两种生境中随海拔梯度增加的增幅均较小,且远远低于所有种和偶见种;湿润生境中所有种和偶见种丰富度沿海拔梯度的变化速率也高于干旱生境(表2)。偶见种丰富度占所有种丰富度的百分比沿海拔梯度的变化呈现为单峰曲线格局,且最低值出现在中海拔段,即低、高海拔段偶见种所占的比例最高(图2)。同一样地内不同样方间物种丰富度差异明显,离散系数(CV)变化范围为0.05~1.02。湿润生境CV值高于干旱生境;两种生境中,CV值均表现为偶见种>所有种>常见种。
2.2 Beta多样性沿海拔梯度的变化
湿润生境中,βc多样性随海拔的升高而下降(图3),而干旱生境中,βc多样性在中海拔区域最低(图3);两种生境中,βw多样性总体上在中海拔区域最高(图3)。除湿润生境偶见种外,物种组成相似性与海拔高差之间均呈显著线性相关关系,偶见种物种组成相似性最低,而常见种物种组成相似性最高(图4)。两种生境中偶见种βC、βW多样性均显著高于常见种(P<0.05),且在9个样地中偶见种βW多样性始终高于常见种;但是,偶见种βslope多样性低于常见种;湿润生境βC和βW多样性高于干旱生境,而βslope多样性低于干旱生境(表2)。

表2 湿润与干旱生境物种多样性比较Table 2 The comparison of plant species diversity between wet and dry habitats
*表示同一指标干旱和湿润两种生境间差异显著(P<0.05),**表示同一指标干旱和湿润两种生境间差异极显著(P<0.01)。斜率,多样性与海拔高差线性回归的斜率。
* and ** indicate significantly difference of the same parameter between dry and wet habitats at 0.05 level and 0.01 level, respectively. Slope, slope of the linear regression between diversity and relative altitude.

图1 物种丰富度沿海拔梯度的分布格局Fig. 1 Patterns of plant species richness along an elevational gradient
2.3 Gamma多样性沿海拔梯度的变化
湿润生境中所有种和偶见种γ多样性随海拔增加而下降,而常见种γ多样性随海拔的增加而增加;干旱生境中所有种、常见种、偶见种γ多样性均随海拔的增加而增加;偶见种γ多样性高于常见种γ多样性,且常见种γ多样性在两种生境中随海拔增加的增幅很小,基本保持不变(表2,图5)。
3 讨论
东达山高寒草甸编目多样性沿海拔梯度的分布格局表现出3个特征。第一,常见种、偶见种多样性格局差异明显。偶见种与所有种编目多样性沿海拔梯度的分布格局几乎一致(表2,图1、5),说明偶见种对群落物种编目多样性的海拔分布格局起着重要作用。高寒草甸植物群落的优势种大多以克隆繁殖为主[30],这些优势种也是群落的常见种,它们的海拔分布范围大,物种编目多样性沿海拔梯度的变化小。同时,偶见种占所有种比例在高寒草甸分布的低、高海拔段最高,即在高寒草甸与森林、与高山流石滩的生态交错区最高(图2)。不同植被类型交错区植物多样性常常都很高[31],而且物种相互作用机制表明部分物种维持最低生物量以保持群落物种的多样性[32],这与偶见种分布频度低、个体数量少的分布特征一致,表明偶见种是生态交错区物种多样性的重要组成部分。第二,编目多样性与海拔梯度的相关性不高。同一样地不同样方间物种丰富度差异明显(图1),丰富度离散程度大,离散系数最大值超过1,且偶见种丰富度的离散程度最大。相邻样方间物种组成及多样性差异大,导致多样性与海拔梯度之间的相关性不高,与东灵山[33]、小五台山[34]亚高山草甸植物多样性沿海拔梯度的分布格局一致。因此,如果取样强度不够也可能导致物种丰富度沿海拔梯度分布格局的假相,如很多研究表明高寒草甸植物群落物种丰富度沿海拔梯度的变化呈现“单峰”曲线格局[35-36]。无论从地质历史、植被分布、气候因子、物种分布几何边界的中间膨胀效应等各个方面,似乎都无法解释这一分布格局。第三,湿润与干旱生境物种多样性沿海拔梯度的分布格局差异明显,两种生境中编目多样性基本呈现为相反的变化趋势(表2,图1、5)。在海拔梯度上编目多样性主要有两种分布格局,一是随海拔的增加而下降的偏峰格局,另一种是随海拔的升高先增加而后下降的单峰曲线格局[37]。但是,不管哪种分布格局,在高海拔段物种丰富度都一致地表现为随海拔的升高而下降的偏峰格局,尤其在高大的山体[38-42]。湿润生境中所有种和偶见种编目多样性沿海拔梯度的分布格局符合这一普遍现象,而干旱生境物种编目多样性与这一普遍分布格局相反。物种多样性沿海拔梯度的分布格局受多种因子的影响,如气候、土壤、植被、生境类型等[43]。本研究中,湿润与干旱生境位置相邻,气候、环境与植被因子沿海拔梯度的变化相似,但物种多样性的海拔分布格局不同,说明两种生境植物群落对外界干扰的抵抗力不同,干旱生境植物群落及物种多样性受外界干扰严重,物种多样性的分布格局已完全改变。大量针对草地开展的研究都表明物种多样性与群落稳定性之间呈现正相关关系,高物种多样性有利于维持群落稳定性[44-46]。高寒草甸干旱生境物种多样性低,群落稳定性低,生境干旱化过程是植物群落退化演替的过程。
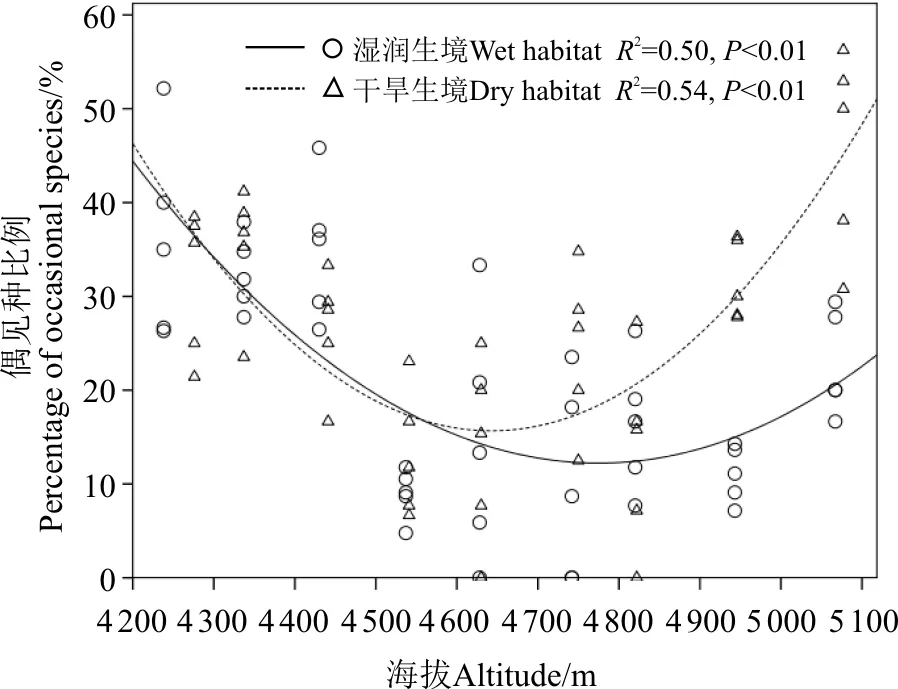
图2 湿润与干旱生境中偶见种丰富度占所有种丰富度比例沿海拔梯度的变化Fig. 2 Percentage of occasional species richness to total species richness varied along an elevational gradient

图3 β多样性沿海拔梯度的变化Fig. 3 Variation of beta diversity along an elevational gradient

图4 Simpson相似指数与海拔高差之间的线性回归关系Fig. 4 Linear regression between Simpson similar index and relative altitude
3种β多样性测度方法的结果不完全一致(图3、4),其中,βC和βW结果表明,偶见种β多样性高于常见种(图3),而βslope结果表明,偶见种β多样性低于常见种(图4)。偶见种在群落中的分布频率低,在环境梯度上的替换速率快,对群落β多样性贡献更高[20-22],与本研究中βC和βW的测度结果一致。βC和βW均为常见的β多样性测度方法,有广泛的使用,其测度结果可靠性高,而βslope指数使用相对较少,既不属于β多样性的倍性测度方法,也不属于β多样性的加性测度方法,其测度结果有待于进一步检验。同时,βC和βW多样性指数在海拔上的变化也不完全相同,偶见种βW多样性始终高于常见种,而对于βC而言,偶见种βC多样性大多数情况下高于常见种,而在一个海拔样地上偶见种βC多样性低于常见种(图3)。因此,βW多样性指数能够更好地区分群落物种的组成,对群落物种组成的变化更加敏感。
4 结论
高寒草甸植物群落中偶见种多样性高,海拔分布格局与所有种一致,直接影响群落物种多样性和海拔分布格局。高寒草甸干旱生境植物群落物种多样性更低,与多样性沿海拔梯度分布的普遍格局相反,是退化演替植物群落物种多样性的分布特征。偶见种在植物群落中的分布频度低,在环境梯度上的替代率高,对β多样性贡献高,且对其变化敏感,其结果可以反映不同β多样性指数的测度效果。
参考文献References:
[1] López-Pujol J,Zhang F M,Sun H Q,Ying T S,Ge S.Centres of plant endemism in China:Places for survival or for speciation?Journal of Biogeography,2011,38(7):1225-1432.
[2] Zhang D C,Ye J X,Sun H.Quantitative approaches to identify floristic units and centres of species endemism in the Qinghai-Tibetan Plateau,south-western China.Journal of Biogeography,2016,43(12):2465-2476.
[3] 马培杰,李亚娇,潘多锋,陈本建,李心诚,王德利.牦牛干扰下草原毛虫对小嵩草高寒草甸植物群落特征的影响.草业科学,2017,34(4):698-705.
Ma P J,Li Y J,Pan D F,Chen B J,Li X C,Wang D L.Effect of grassland caterpillar on the characteristics of the vegetation ofKobrsiapygmaeaalpine grassland under the interference of the yak grazing.Pratacultural Science,2017,34(4):698-705.( in Chinese)
[4] Dumont B,Garel J P,Ginane C,Decuq F,Farruggia A,Pradel P,Rigolot C,Petit M.Effect of cattle grazing a species-rich mountain pasture under different stocking rates on the dynamics of diet selection and sward structure.Animal:An International Journal of Animal Bioscience,2007,1(7):1042-1052.
[5] 林丽,张德罡,曹广民,欧阳经政,刘淑丽,张法伟,李以康,郭小伟.高寒嵩草草甸植物群落数量特征对不同利用强度的短期响应.生态学报,2016,36(24):8034-8043.
Lin L,Zhang D G,Cao G M,Ouyang J Z,Liu S L,Zhang F W,Li Y K,Guo X W.Plant functional groups numerical characteristics responses to different grazing intensities under different community succession stages of alpineKobresiameadow in spring.Acta Ecologica Sinica,2016,36(24):8034-8043.(in Chinese)
[6] 周丽,张德罡,贠旭江,董永平,王加亭,赵雅丽.退化高寒草甸植被与土壤特征.草业科学,2016,33(11):2196-2201.
Zhou L,Zhang D G,Yun Y X,Dong Y P,Wang J T,Zhao Y L.The vegetation and soil characteristics of degraded alpine meadow.Pratacultural Science,2016,33(11):2916-2201.(in Chinese)
[7] 谢妍洁,朱玉怀,张大才,李双智.2种嵩草属植物形态特征对生境干旱化的响应.西北植物学报,2016,36(3):796-803.
Xie Y J,Zhu Y H,Zhang D C,Li S Z.Response of morphological character to aridification of habitat for two plants of genusKobresia.Acta Botanica Boreali-Occidentalia Sinica,2016,36(3):796-803.(in Chinese)
[8] 邱波,任青吉,罗燕江,杜国祯.高寒草甸不同生境类型植物群落的α及β多样性研究.西北植物学报,2004,24(4):655-661.
Qiu B,Ren Q J,Luo Y J,Du G Z.Study on α diversity and β diversity of plant community of different habitats in alpine meadow.Acta Botanica Boreali-Occidentalia Sinica,2004,24(4):655-661.(in Chinese)
[9] 马凯,李永宁,金辉,龙在海,卢银平.不同生境类型金莲花群落物种多样性比较.草业科学,2011,28(8):1467-1472.
Ma K,Li Y N,Jin H,Long Z H,Lu Y P.A comparison of the species diversity ofTrolliuschinensiscommunity in different habitats.Pratacultural Science,2011,28(8):1467-1472.(in Chinese)
[10] 仁青吉,武高林,任国华.放牧强度对青藏高原东部高寒草甸植物群落特征的影响.草业学报,2009,18(5):256-261.
Ren Q J,Wu G L,Ren G H.Effect of grazing intensity on characteristics of alpine meadow communities in the eastern Qinghai-Tibetan Plateau.Acta Ecologica Sinica,2009,18(5):256-261.(in Chinese)
[11] 金晓明,韩国栋.放牧对草甸草原植物群落结构及多样性的影响.草业科学,2010,27(4):7-10.
Jin X M,Han G D.Effects of grazing intensity on species diversity and structure of meadow steppe community.Pratacultural Science,2010,27(4):7-10.(in Chinese)
[12] 纪磊,干友民,罗元佳,成平,冯国英,马超.川西北不同退化程度高山草甸和亚高山草甸的植被特征.草业科学,2011,28(6):1101-1105.
Ji L,Gan Y M,Luo Y J,Cheng P,Feng G Y,Ma C.Comparison between the vegetation characteristics of alpine and subalpine meadow with different degradation degrees in North-west of Sichuan Province.Pratacultural Science,2011,28(6):1101-1105.(in Chinese)
[13] 刘佳慧,白璐,梁宏伟,张韬,崔玉刚.典型草原退化群落恢复演替过程中双齿葱组织变化研究.内蒙古农业大学学报,2008,29(4):248-252.
Liu J H,Bai L,Liang H W,Zhang T,Cui Y G.Research on aLiliumbidentatumorgan cell change in the course of restoring succession of the typical steppe degraded community.Journal of Inner Mongolia Agricultural University,2008,29(4):248-252.(in Chinese)
[14] 杨元武,李希来.不同退化程度高寒草甸高山嵩草的构件变化.西北植物学报,2011,31(1):167-171.
Yang Y W,Li X L.Modular variety ofKobresiapygmaeaon alpine meadow with different degraded levels.Acta Botanica Boreali-Occidentalia Sinica,2011,31(1):167-171.(in Chinese)
[15] 郝建峰,李艳,齐锦秋,裴曾莉,黄雨佳,蒋倩,陈亚.人为干扰对碧峰峡栲树次生林群落物种多样性及其优势种群生态位的影响.生态学报,2016,36(23):7678-7688.
Hao J F,Li Y,Qi J Q,Pei Z L,Huang Y J,Jiang Q,Chen Y.Effects of anthropogenic disturbances on the species diversity and niche of the dominant populations in aCastanopsisfargesiisecondary forest community in Bifengxia,Sichuan.Acta Ecologica Sinica,2016,36(23):7678-7688.(in Chinese)
[16] Jurasinski G,Retzer V,Beierkuhnlein C.Inventory,differentiation,and proportional diversity:A consistent terminology for quantifying species diversity.Oecologia,2009,159(1):15-26.
[17] Koleff P,Gaston K J,Lennon J J.Measuring beta diversity for presence-absence data.Journal of Animal Ecology,2003,72(3):367-382.
[18] 陈圣宾,欧阳志云,徐卫华,肖燚.Beta多样性研究进展.生物多样性,2010,18(4):323-335.
Chen S B,Ouyang Z Y,Xu W H,Xiao Y.A review of beta diversity studies.Biodiversity Science,2010,18(4):323-335.(in Chinese)
[19] Whittaker R H.Vegetation of the Siskiyou Mountains,Oregon and Califomia.Ecological Monographs,1960,30(3):279-338.
[20] Ulrich W,Ollik M.Frequent and occasional species and the shape of relative-abundance distributions.Diversity and Distributions,2004,10(4):263-269.
[21] Klimek S,Marini L,Hofmann M,Isselstein J.Additive partitioning of plant diversity with respect to grassland management regime,fertilisation and abiotic factors.Basic and Applied Ecology,2008,9(6):626-634.
[22] 卢训令,汤茜,梁国付,丁圣彦.黄河下游平原非农植物多样性拆分研究.生态学报,2016,36(14):4395-4405.
Lu X L,Tang Q,Liang G F,Ding S Y.Additive partitioning of non-agricultural plant diversity in the lower reaches of the Yellow River plain.Acta Ecologica Sinica,2016,36(14):4395-4405.(in Chinese)
[23] 沈渭寿,刘波,颜长珍,张慧.青藏高原典型区生态状况时空变化及气候变化响应研究.北京:中国环境出版社,2013.
Shen W T,Liu B, Yan C Z,Zheng H.Temporal and Spatial Variation of Ecological Status and Response to Climate Change in Typical Areas of Qinghai-Tibet Plateau.Beijing:China Environmental Science Press,2013.(in Chinese)
[24] Dolan J R,Ritchie M E,Tunin-Ley A,Pizay M D.Dynamics of core and occasional species in the marine plankton:Tintinnid ciliates in the north-west Mediterranean Sea.Journal of Biogeography,2009,36(5):887-895.
[25] 牛钰杰,杨思维,王贵珍,刘丽,花立民.放牧干扰下高寒草甸物种多样性指数评价与选择.应用生态学报,2017,28(6):1824-1832.
Niu Y J,Yang S W,Wang G Z,Liu L,Hua L M.Evaluation and selection of species diversity index under grazing disturbance in alpine meadow.Chinese Journal of Applied Ecology,2017,28(6):1824-1832.(in Chinese)
[26] 程寅瑞,张小平.小气候对紫金山南北坡植物群落组成的影响.安徽师范大学学报(自然科学版),2012,35(1):44-48.
Cheng Y R,Zhang X P.Microclimate’s affection on the composition of plant community on southern and northern slope of Purple mountain.Journal of Anhui Normal University(Natural Science),2012,35(1):44-48.(in Chinese)
[27] Cody M L.Towards a theory of continental species diversities:Bird distributions over Mediterranean habitat gradients.//Cody M L,Diamond J M.(eds).Ecology and evolution of communities.Harvard:Harvard University Press,1975:214-257.
[28] Soininen J,McDonald R,Hillebrand H.The distance decay of similarity in ecological communities.Ecography,2007,3(1):3-12.
[29] Qian H.Beta diversity in relation to dispersal ability for vascular plants in North America.Global Ecology and Biogeography,2009,18(3):327-332.
[30] 邓自发,谢晓玲,王启基,周兴民.高寒草甸藏嵩草种群繁殖对策的研究.应用与环境生物学报,2001,7(4):332-334.
Deng Z F,Xie X L,Wang Q J,Zhou X M.Study on reproduction strategies ofKobresiatibeticapopulation on alpine meadow.Chinese Journal of Applied and Environmental Biology,2001,7(4):332-334.(in Chinese)
[31] 高俊峰,张芸香.关帝山次生林区典型森林交错带物种多样性研究.西北植物学报,2005,25(10):2017-2023.
Gao J F,Zhang Y X.Species diversity in overlapped zones of typical secondary forests in Guandishan Mountains.Acta Botanica Boreali-Occidentalia Sinica,2005,25(10):2017-2023.(in Chinese)
[32] Isbell F,Polley H W,Wilsey B J.Species interaction mechanisms maintain grassland plant species diversity.Ecology,2009,90(7):1821-1830.
[33] 向春玲,张金屯.东灵山亚高山草甸物种多样性变化及其影响因子.北京师范大学学报(自然科学版),2009,45(3):275-278.
Xiang C L,Zhang J T.Changes in species diversity and contributing factors in subalpine meadow in Dongling Mountain.Journal of Beijing Normal University(Natural Science),2009,45(3):275-278.(in Chinese)
[34] 白晓航,张金屯,曹科,王云泉,Sehrish S,曹格.小五台山亚高山草甸的群落特征及物种多样性.草业科学,2016,33(12):2533-2543.
Bai X H,Zhang J T,Cao K,Wang Y Q,Sehrish S,Cao G.Community characteristics and species diversity of subalpine meadows in Xiaowutai Mountain.Pratacu1tural Science,2016,33(12):2533-2543.(in Chinese)
[35] 卢慧,丛静,刘晓,王秀磊,唐军,李迪强,张于光.三江源区高寒草甸植物多样性的海拔分布格局.草业学报,2015,24(7):197-204.
Lu H,Cong J,Liu X,Wang X L,Tang J,Li D Q,Zhang Y G.Plant diversity patterns along altitudinal gradients in alpine meadows in the Three River Headwater Region,China.Acta Prataculturae Sinica,2015,24(7):197-204.(in Chinese)
[36] 刘哲,李奇,陈懂懂,翟文婷,赵亮,徐世晓,赵新全.青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响.生物多样性,2015,23(4):451-462.
Liu Z,Li Q,Chen D D,Zhai W T,Zhao L,Xu S X,Zhao X Q.Patterns of plant species diversity along an altitudinal gradient and its effect on above-ground biomass in alpine meadows in Qinghai-Tibet Plateau.Biodiversity Science,2015,23(4):451-462.(in Chinese)
[37] 唐志尧,方精云.植物物种多样性的垂直分布格局.生物多样性,2004,12(1):20-28.
Tang Z Y,Fang J Y.A review on the elevational patterns of plant species diversity.Biodiversity Science,2004,12(1):20-28.(in Chinese)
[38] Vetaas O R,Grytnes J A.Distribution of vascular plant species richness and endemic richness along the Himalayan elevational gradient in Nepal.Global Ecology & Biogeography,2002,11(4):291-301.
[39] Bhattarai K R,Vetaas O R.Variation in plant species richness of different life forms along a subtropical elevational gradient in the Himalayas,east Nepal.Global Ecology & Biogeography,2003,12(4):327-340.
[40] Bhattarai K R,Vetaas O R,Grytnes J A.Fern species richness along a central Himalayan elevational gradient,Nepal.Journal of Biogeography,2004,31(3):389-400.
[41] Carpenter C.The environmental control of plant species density on aHimalayanelevational gradient.Journal of Biogeography,2005,32(6):999-1018.
[42] Zhang D C,Zhang Y H,Boufford D E,Sun H.Elevational patterns of species richness and endemism for selected taxa in the Hengduan Mountains,southwestern China.Biodiversity Conservation,2009,18(3):699-716.
[43] Báldi A.Habitat heterogeneity overrides the species-area relationship.Journal of Biogeography,2008,35(4):675-681.
[44] Bai Y F,Han X G,Wu J G,Chen Z Z,Li L H.Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Nature,2004,431:181-184.
[45] Hector A,Hautier Y,Saner P,Wacker L,Bagchi R,Joshi J,Scherer-Lorenzen M,Spehn E M,Bazeley-White E,Weilenmann M,Caldeira M C,Dimitrakopoulos P G,Finn J A,Huss-Danell K,Jumpponen A,Mulder C P H,Palmborg C,Pereira J S,Siamantziouras A S D,Terry A C,Troumbis A Y,Schmid B,Loreau M.General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding.Ecology,2010,91(8):2213-2220.
[46] Tilman D,Reich P B,Knops J M H.Biodiversity and ecosystem stability in a decade-long grassland experiment.Nature,2006,441:629-632.
