关帝山不同海拔土壤碳矿化和微生物特征
2018-05-26李君剑杜宏宇严俊霞李洪建
李君剑,杜宏宇,刘 菊,严俊霞,李洪建
关帝山不同海拔土壤碳矿化和微生物特征
李君剑1*,杜宏宇1,刘 菊2,严俊霞1,李洪建1
(1.山西大学黄土高原研究所,山西 太原 030006;2.山西省林业科学院,山西 太原 030012)
以山西省关帝山不同海拔的8个样地土壤为研究对象,分析不同海拔土壤碳氮含量、土壤碳矿化和细菌、真菌、放线菌、固氮菌、硝化细菌和反硝化细菌6类微生物量的变化.结果表明,海拔为2163m时,土壤有机碳和活性碳1(LC1)最高,分别为74.41和6.72g/kg,在海拔2700m总氮含量最高为6.54g/kg.土壤碳矿化累积量和碳矿化速率因海拔的升高而显著增加(<0.05),土壤碳矿化累积量和活性碳2(LC2)之间有显著正相关.土壤细菌、放线菌和固氮菌的数量在海拔为1791m时最高,分别为3.41´106、1.90´106和1.21´106个/g,细菌、真菌、放线菌和固氮菌的数量随着海拔的升高而逐渐下降,与海拔之间呈显著负相关;相反,硝化细菌和反硝化细菌数量却逐渐增加,在海拔为2700m时最高,分别为1.37´104和6.02´103个/g,它们与海拔和总氮之间的关系呈显著正相关(<0.01).总之,土壤有机碳、总氮、碳矿化累积量、硝化细菌和反硝化细菌随海拔显著增加,而细菌、真菌、放线菌和固氮菌随海拔的变化呈相反的趋势.
海拔;土壤碳矿化;微生物量;活性碳
土壤碳库是陆地碳库的重要组成部分,土壤有机碳的分解和累积,一方面影响了土壤肥力和植物生长,另一方面直接影响着全球的碳平衡[1].全球土壤有机碳储量约为1500Pg(1Pg=1015g),土壤有机质可能是土壤碳汇,通过植物光合作用固定CO2,相反也可能是土壤碳源,由土壤微生物和其他生物释放的CO2量约是石油燃烧排放量的10倍[2].土壤有机碳动态转化过程的研究,不仅是土地资源可持续利用的重要基础,而且在土壤碳循环与全球气候变化的相互作用研究中具有重要意义[3-4].
海拔高度的变化会改变植被的组成、结构、养分、有机质分解、微生物活性等一系列因子,它们都可能显著影响土壤有机碳的动态变化[5-6].微生物可以对有机质进行矿化和分解[7],土壤微生物群落是土壤有机碳动态的关键调节者,有机碳的矿化受微生物数量和活性调节,因此土壤微生物可能对土壤碳矿化有重要影响.
目前国内学者越来越重视土壤碳矿化和土壤微生物的研究,通过比较武夷山不同海拔土壤的矿化速率和矿化比率的差异,得出不同海拔高度土壤碳矿化速率随海拔高度的升高而加快,土壤碳矿化比率随着海拔的变化而产生显著差异[8].土壤微生物在陆地生态系统循环方面有着重要的作用,相关研究也越来越多.通过对贺兰山不同海拔的6个典型植被带的土壤微生物多样性研究,得出土壤微生物功能多样性随着海拔的增加发生变化,且不同海拔的微生物群落结构存在显著差异[9].然而对于不同海拔土壤碳矿化累积量,矿化速率,碳矿化势和微生物量的共同研究较少.因此,本研究以山西省庞泉沟关帝山8个不同海拔样地土壤为对象,对样地的土壤理化特征,碳矿化累积量,碳矿化速率和碳矿化势,以及土壤中细菌、真菌、放线菌、固氮菌、硝化细菌和反硝化细菌的数量进行测定.通过分析不同海拔土壤的理化特性,碳矿化累积量和碳矿化速率的差异性、不同指标之间的相关性,来揭示海拔对土壤有机碳矿化的影响,为土壤碳循环和全球变暖等气候变化的研究提供科学参考.
1 研究区域与方法
1.1 研究区域
关帝山位于吕梁山中段(37°20′N~38°20′N, 110°18′E~111°18′E),主峰是孝文峰,海拔2831m.该区属于温带半湿润大陆性气候,年平均气温3℃~4℃,年降水量830.8mm,多集中在7月和8月.土壤类型从低海拔到高海拔依次为淡褐土,淋溶褐土,棕壤和亚高山草甸土.
1.2 样品采集
研究区位于关帝山庞泉沟自然保护区实验区八道沟.本研究于2014年8月进行随机调查取样.共设置8个海拔分别为1791、1986、2105、2163、2264、2387、2621和2700m的样地,每个样地有3个重复区域,使用多点混合土样样品处理法.取样前,先将地表植物和枯落物去除,使用土钻随机取0~10cm的土壤,带回实验室后过2mm的筛子,土样分为两部分,一部分风干后进行土壤理化性质指标的测定,另一部分在4℃下保存以备土壤微生物量的测定.
1.3 样品分析
有机碳采用重铬酸钾外加热法,总氮采用半微量凯式定氮法测定[10].土壤活性碳采用Rovira和Vallejo酸水解法,分别加入2.5和13mol/L的H2SO4进行依次酸解并离心,酸解液体分别通过重铬酸钾氧化法测定碳含量,被低和高浓度H2SO4酸解并浸提出土壤有机碳分别为活性碳1和活性碳2(LC1和LC2)[11].
土壤碳矿化量测定采用室内恒温培养,碱液吸收法,在25℃下培养56d,分别在培养2,4,6,8, 15,22,29,35,54d时用HCl滴定吸收液NaOH,从而计算土壤碳矿化速率[12].
土壤碳矿化势主要是易降解碳分解,所以应用一阶动态方程(式(1)[13])分析不同处理下土壤碳矿化动态.单因素模型是基于土壤矿化量和时间成正比而拟合得到.
=(1-e-kt) (1)
式中:为碳矿化势;为矿化常数;为时间,d;为d时碳矿化累积量,mg/kg.
微生物的数量采用稀释平板法测定,细菌培养基为牛肉蛋白胨培养基,放线菌培养基为高氏一号(苯酚500mg/L),真菌培养基为马丁氏培养基(链霉素30mg/L),固氮菌培养基为无氮阿须贝氏培养基,硝化细菌和反硝化细菌培养基见参考文献[14].
1.4 数据分析
采用SPSS 19.0的单因素方差分析(one-way ANOVA)不同海拔土壤理化性质、土壤碳矿化累积量和碳矿化潜势之间的差异性,各变量之间的相关性采用Pearson相关分析.
2 结果与分析
2.1 土壤理化性质
从表1可以看出,随着海拔的升高,有机碳和LC1的含量有明显的增加趋势,总氮含量和碳矿化累积量有显著差异.不同海拔下,LC2未表现出规律性的变化趋势,而LC1有规律性的变化.总氮含量在海拔2700m时最大,在海拔为1791m时最低,并且随着海拔的升高而逐渐增加.
2.2 土壤碳矿化累积量和矿化速率
土壤碳矿化累积量在海拔1791m时最低,随海拔的升高显著增加,并且各海拔之间均呈较显著差异(表1).土壤的碳矿化速率随培养时间的增加而下降,随着海拔的升高,碳矿化速率也总体升高(图1).
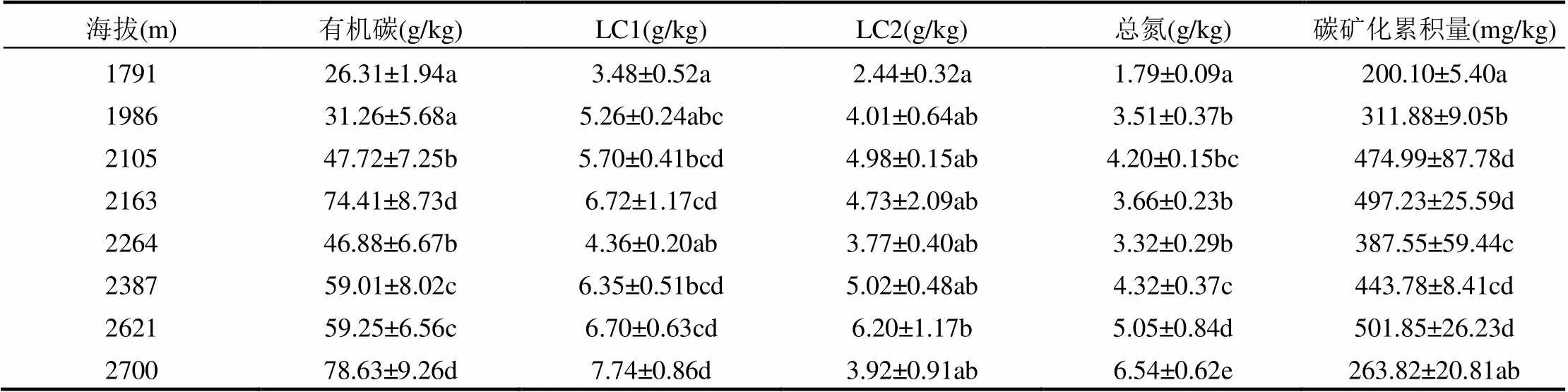
表1 不同海拔土壤的理化性质和碳矿化累积量
注:不同小写字母表示样地在0.05水平差异显著.
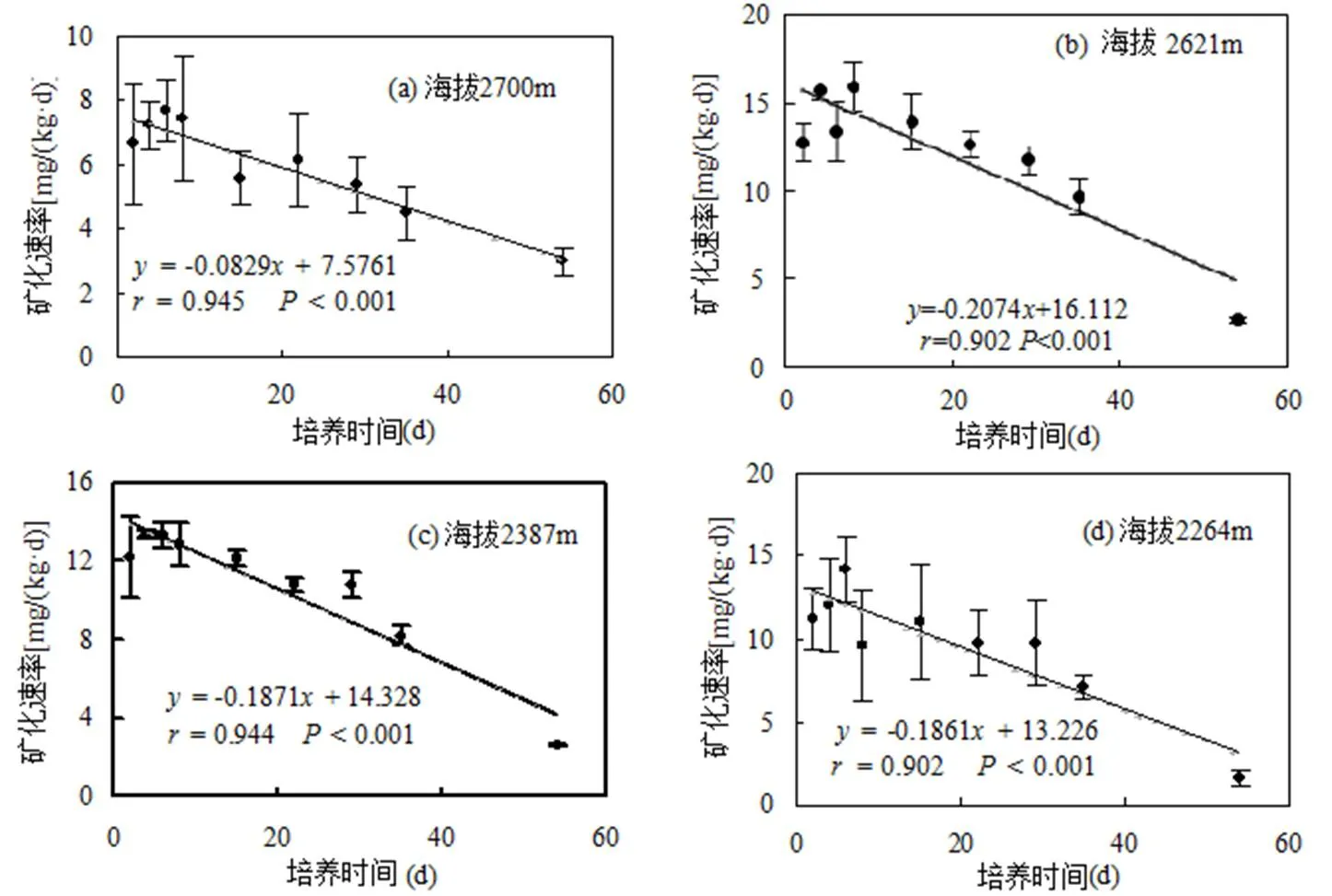
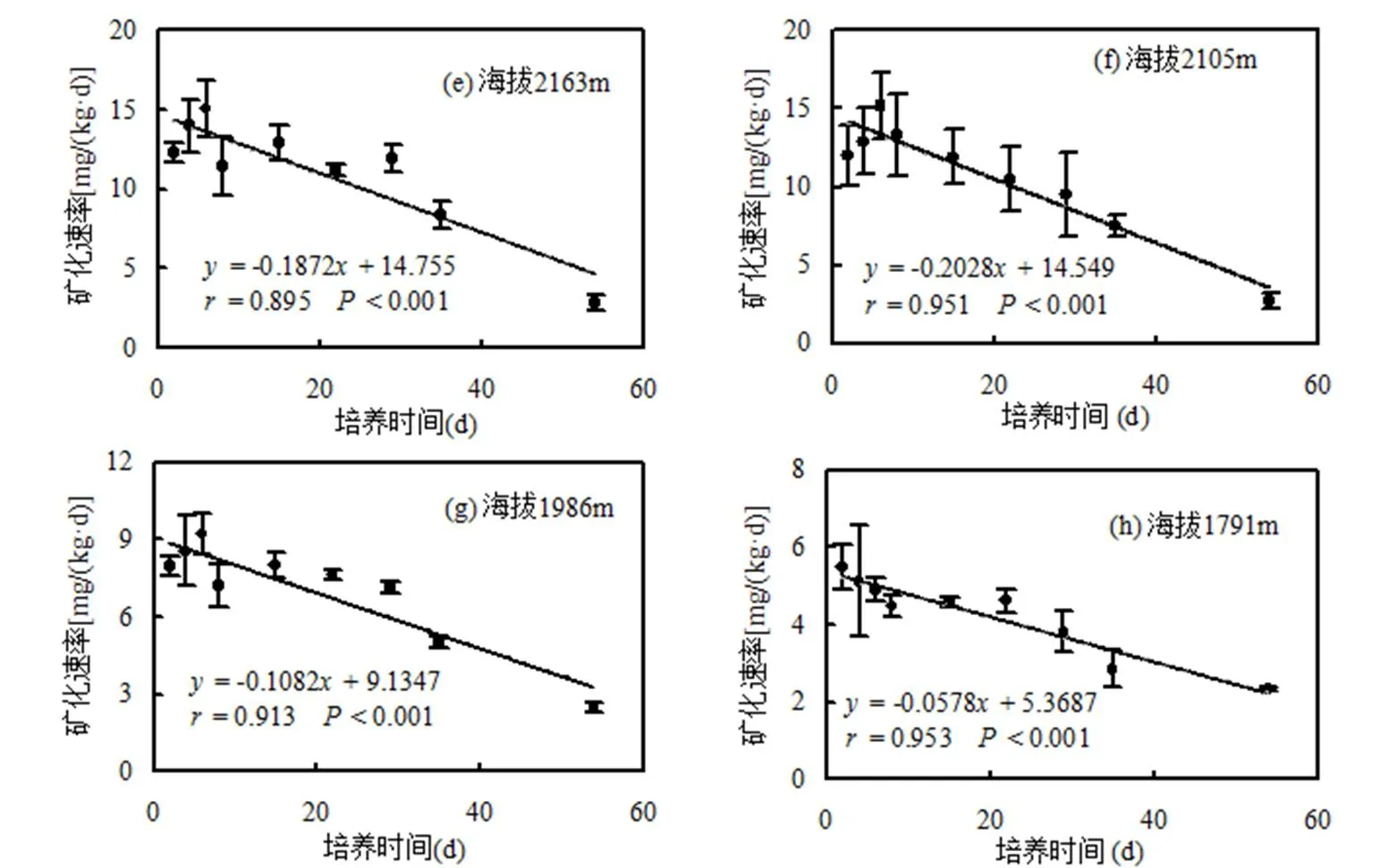
图1 不同海拔下土壤碳矿化速率随培养时间的变化
由表2可见,土壤碳矿化势与培养天数之间一阶动态方程的拟合系数均达到了极显著相关水平(<0.001).碳矿化势因海拔的升高而呈现总体升高的趋势.

表2 不同海拔土壤的碳矿化势
2.3 土壤的微生物量
随着海拔的升高,土壤中可培养细菌、放线菌、真菌和固氮细菌的数量都减少,而硝化细菌和反硝化细菌的数量却增多(图2).细菌、放线菌和固氮细菌在海拔1791m时数量最多,而真菌数量在2163m时呈现最多,但是硝化细菌和反硝化细菌在2700m时数量最多.在海拔1791~2387m之间的土壤细菌、放线菌、真菌和固氮细菌的数量都有较显著差异,海拔2387m以上的土壤中,细菌、放线菌、真菌和固氮细菌之间都没有显著差异.在海拔为2163m时,土壤的细菌、放线菌、真菌和固氮细菌和其他海拔的数量均有显著差异.硝化细菌在2387m以上的4个不同海拔之间有增加的趋势.
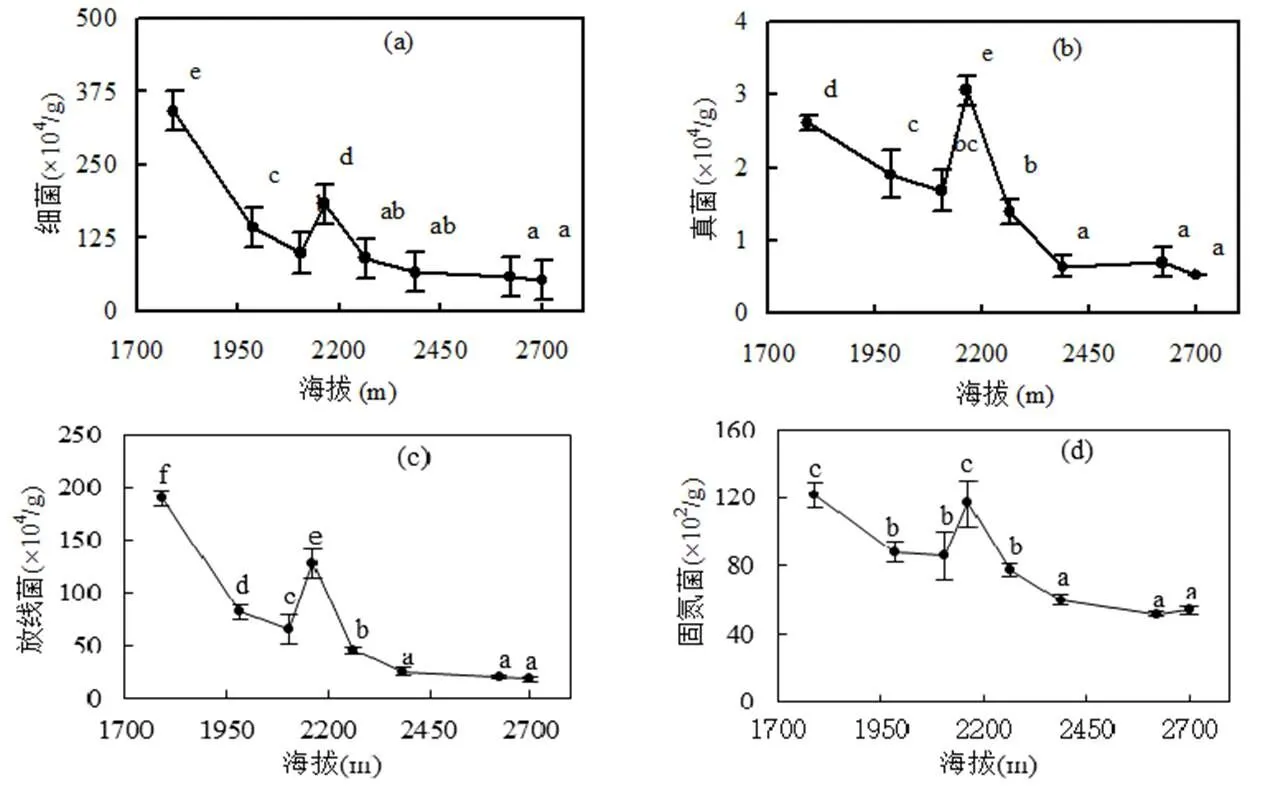
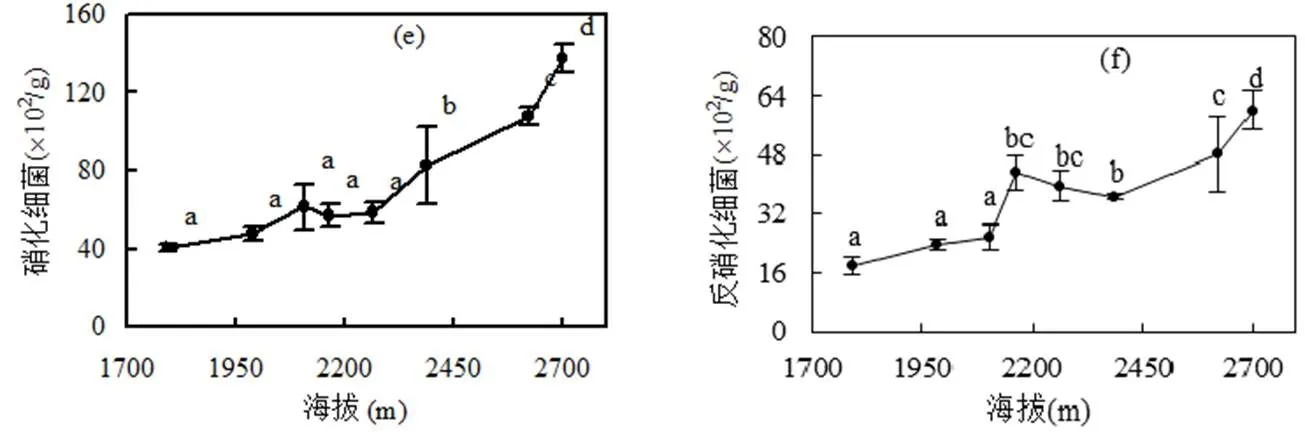
图2 不同海拔下土壤中不同微生物的数量
2.4 相关性分析
土壤有机碳与土壤LC1呈显著相关(<0.01).土壤碳矿化累积量和土壤LC2之间有极显著相关.土壤总氮含量与土壤LC1、海拔、硝化细菌和反硝化细菌的数量有显著正相关(<0.01)(表3).
海拔与细菌、真菌、放线菌和固氮菌的数量有显著负相关,而与硝化细菌和反硝化细菌的数量之间有显著正相关.固氮菌和细菌、真菌、放线菌之间有显著正相关(<0.01),硝化细菌和反硝化细菌之间在<0.01水平上呈正相关.
3 讨论
本研究表明,土壤有机碳、LC1和总氮因海拔的不同均有显著的变化(表1),海拔和总氮呈显著正相关,与有机碳,LC1之间在<0.05水平上呈正相关(表3).而不同的研究有不同的结果,有研究得出不同海拔植被带土壤化学性质差异显著,其中土壤有机碳、总氮等含量随海拔的上升呈现先降低后升高的趋势[15].而吴则焰等[16]研究表明中亚热带森林土壤的化学性质随海拔的上升而逐渐下降.这些均与本研究的结果不同,这可能与本研究不同的经纬度、气候类型和植被类型等有关.Du等[17]发现江西庐山土壤有机碳含量随着海拔的升高而增加.与本研究有着相同结果的是刘秉儒[18]得出贺兰山东坡典型植被土壤沿海拔梯度的变化土壤有机碳和总氮沿海拔梯度呈增加,是由于海拔的不同导致植物群落的种类组成和结构不同,植物根系对土壤的有机质的量有不同的影响.而本研究中可能是因为随着海拔的升高,温度降低,土壤微生物的活性也降低,对土壤有机质的分解变慢,所以随着海拔的升高土壤有机碳和总氮含量增加.土壤碳矿化累积量随着海拔梯度变化显著增加(表1).有研究表明,不同海拔土壤碳矿化速率随海拔的升高而加快[19-20],这与本研究结果类似,高海拔土壤碳矿化速率高于低海拔.这是因为在室内培养实验过程中,温度是恒定的,有机碳,活性碳含量随海拔的升高而升高,使得土壤的碳矿化速率也加快,正如表3中呈现的土壤碳矿化累积量和LC2呈显著正相关(<0.01),王红等[21]研究也发现土壤中有机碳含量的增加会显著提高碳矿化速率,土壤氮的含量也会对土壤碳矿化速率产生影响[21].土壤碳矿化速率和碳矿化累积量是正相关的,有机碳矿化势也是如此,所以因海拔升高土壤碳矿化速率加快,碳矿化势也增加.
土壤微生物是土壤生物中最活跃的部分,它参与土壤有机质的分解和土壤养分转化等方面.影响微生物量和活性碳的因素有很多,如土壤温度、含水量、营养成分、凋落物种类和数量等[21],而土壤温度、湿度和矿质元素水平是影响土壤微生物种群结构和生物量的关键[22].本研究发现土壤中细菌、真菌、放线菌和固氮菌的数量随着海拔的升高显著降低(图2),它们与海拔之间呈显著负相关(表3).由于高海拔地区温度较低,其土壤微生物活性受到温度限制,土壤的碳矿化速率也较慢[23-24],不利于微生物的生长和繁殖,微生物数量也较少,温度升高增加了参与呼吸作用的微生物数量和种类[7,25].真菌,放线菌的含量与总氮和有机碳含量之间在<0.05的水平上呈负相关.这个结果也得到印证,说明在本研究样地中土壤有机质含量和肥力不是影响土壤微生物量的主要因素,而土壤温度是影响微生物量的重要因素.表3中总氮含量与固氮菌数量呈负相关,与硝化细菌和反硝化细菌数量呈正相关(<0.01).这可能是由于海拔越高,土壤中的总氮含量也越高,固氮作用相对减少,则土壤中的固氮菌就会减少,而硝化细菌和反硝化细菌反而由于氮含量较多而增加.
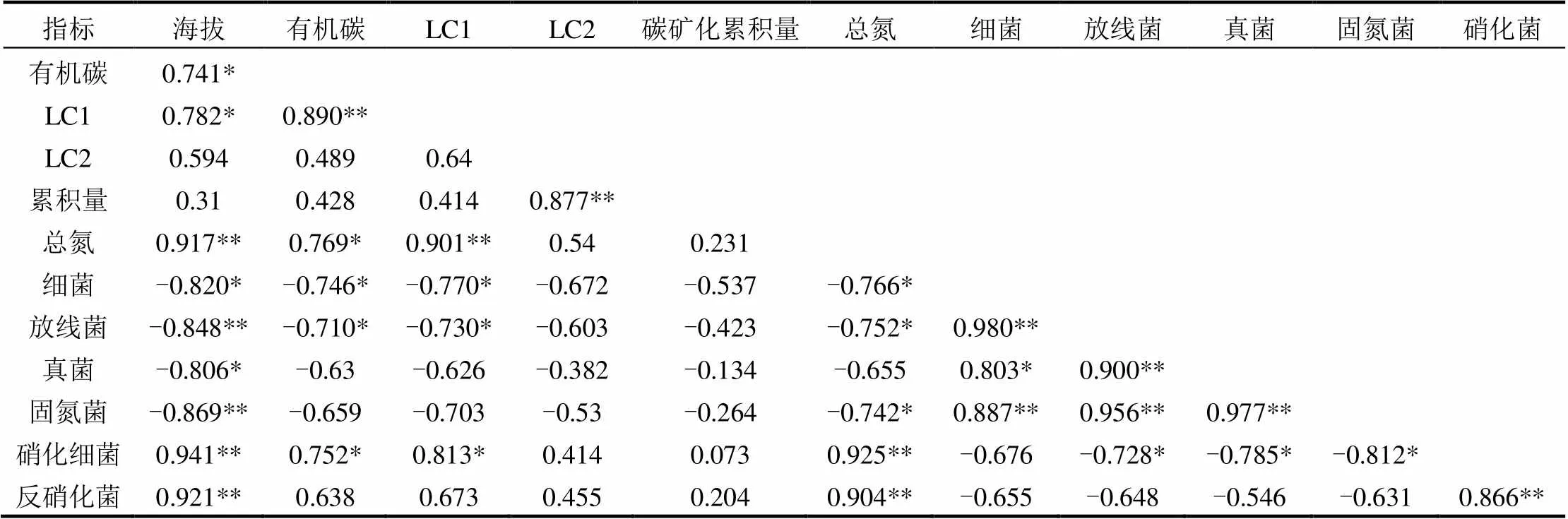
表3 土壤碳氮、碳矿化累积量以及微生物量之间的Pearson相关分析
注:*表示相关达到显著水平(<0.05).(双尾检验),**表示相关达到极显著水平(<0.01).(双尾检验).
4 结论
4.1 随着海拔的升高,土壤有机碳、活性碳、总氮、碳矿化累积量和碳矿化速率显著增加,土壤有机碳与LC1呈显著正相关,土壤碳矿化累积量和LC2之间呈显著正相关,土壤总氮与LC1呈正相关.
4.2 海拔与土壤细菌、真菌、放线菌和固氮菌的数量有显著负相关,而与硝化细菌和反硝化细菌的数量之间有显著正相关,土壤总氮与硝化细菌和反硝化细菌呈正相关.
[1] 周 焱.武夷山不同海拔土壤有机碳库及其矿化特征 [D]. 南京:南京林业大学, 2009.
[2] Amundson R. The carbon budget in soils [J]. Annual Review of Earth and Planetary Sciences, 2001,29:535–562.
[3] Six J, Conant R T, Paul E A, et al. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils [J]. Plant and soil, 2002,241:155-176.
[4] Haynes R J. Labile organic matter as an indicator of organic matter quality in arable and pastoral soils in New Zealand [J]. Soil Biology & Biochemistry, 2000,32:211-219.
[5] Zou X M, Ruan H H, Fu Y, et al. Estimating soil labile organic carbon and potential turnover rates using a sequential fumigation– incubation procedure [J]. Soil Biology & Biochemistry, 2005, 37:1923-1928.
[6] Rodeghiero M, Cescatti A. Main determinant s of forest soil respiration along an elevation /temperature gradient in the Italian Alps [J]. Global Change Biology, 2005,11:1024-1041.
[7] 刘 远,张 辉,熊明华,等.气候变化对土壤微生物多样性及其功能的影响 [J]. 中国环境科学, 2016,36(12):3793-3799.
[8] 周 焱,徐宪根,阮宏华,等.武夷山不同海拔高度土壤有机碳矿化速率的比较 [J]. 生态学杂志, 2008,27(11):1901-1907.
[9] 刘秉儒,张秀珍,胡天华,等.贺兰山不同海拔典型植被带土壤微生物多样性 [J]. 生态学报, 2013,33(22):7211-7220.
[10] 鲍士旦.土壤农化分析 [M]. 北京:中国农业出版社, 2005,66.
[11] Rovira P, Vallejo V R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: an acid hydrolysis approach [J]. Geoderma, 2002,107(1/2): 109-141.
[12] 邵月红,潘剑君,孙 波,等.不同森林植被下土壤有机碳的分解特征及碳库研究 [J]. 水土保持学报, 2005,19(3):24-28.
[13] Hyvönen R, Ågren G I, Dalias P. Analyzing temperature response of decomposition of organic matter [J]. Global Change Biology, 2005,11(5):770–778.
[14] 许光辉,郑洪元.土壤微生物分析方法手册 [M]. 北京:农业出版社, 1986.
[15] 宋贤冲,郭丽梅,田红灯,等.猫儿山不同海拔植被带土壤微生物群落功能多样性 [J]. 生态学报, 2017,37(16):1-8.
[16] 吴则焰,林文雄,陈志芳,等.中亚热带森林土壤微生物群落多样性随海拔梯度的变化 [J]. 植物生态学报, 2013,37(5):397-406.
[17] Du B M, Kang H Z, Jukka Pumpanen, et al. Soil organic carbon stock and chemical composition along an altitude gradient in the Lushan Mountain, subtropical China [J]. Ecological Research, 2014,29:433-439.
[18] 刘秉儒.贺兰山东坡典型植物群落土壤微生物量碳、氮沿海拔梯度的变化特征 [J]. 生态环境学报, 2010,19(4):883-888.
[19] Xu X, Zhou Y, Ruan H H, et al. Temperature sensitivity increases with soil organic carbon recalcitrance along an elevational gradient in the Wuyi Mountains, China [J]. Soil Biology & Biochemistry, 2010,42:1811-1815.
[20] Bu X L, Ruan H H, Wang L M, et al. Soil organic matter in density fractions as related to vegetation changes along an altitude gradient in the Wuyi Mountains, southeastern China [J]. Applied Soil Ecology, 2012,52:42-47.
[21] 王 红,范志平,邓东周,等.不同环境因子对樟子松人工林土壤有机碳矿化的影响 [J]. 生态学杂志, 2008,27(9):1469-1475.
[22] 薛 萐,李 鹏,李占斌,等.不同海拔对干热河谷土壤微生物量及活性的影响 [J]. 中国环境科学, 2011,31(11):1888-1895.
[23] 杨庆朋,徐 明,刘洪升,等.土壤呼吸温度敏感性的影响因素和不确定性 [J]. 生态学报, 2011,31(8):2301-2311.
[24] Mikan C J, Schimel J P, Doyle A P. Temperature controls of microbial respiration in arctic tundra soils above and below freezing [J]. Soil Biology & Biochemistry, 2002,34(11):1785- 1795.
[25] Weltzin J F, Bridgham S D, Pastor J, et al. Potential effects of warming and drying on peatland plant community composition [J]. Global Change Biology, 2003,9(2):141-151.
Soil organic mineralization and microbial characteristics along an altitudinal gradient in Guandi Mountain.
LI Jun-jian1*, DU Hong-yu1, LIU Ju2, YAN Jun-xia1, LI Hong-jian1
(1.Institute of Loess Plateau, Shanxi University, Taiyuan 030006, China;2.Shanxi Academy of Forestry Science, Taiyuan 030012, China)., 2018,38(5):1811~1817
Eight soil samples from different elevations were collected in Guandi Mountain, Shanxi Province. We measured soil organic and liable carbon, total nitrogen, carbon mineralization and microbial populations (including bacteria, fungi, actinomyces, azotobacter, nitrifying and denitrifying bacteria) along an altitude gradient. The results showed that the highest soil organic and liable carbon 1(LC1) at an altitude of 2163m were 74.41 and 6.72g/kg, respectively. The highest total nitrogen was 6.54g/kg at 2700m altitude. Both the accumulation and rate of soil carbon mineralization increased significant with the increase of altitude. There was a significantly positive correlation between soil carbon mineralization and liable carbon 2 (LC2). At an altitude of 1791m, the most populations of bacteria, actinomyces and azotobacter were 3.41´106, 1.90´106and 1.21´106cfu/g, respectively, however, there were the lowest nitrifying and denitrifying bacteria (4.05´103and 1.79´103cfu/g, respectively). Soil bacteria, fungi, actinomyces and azotobacter demonstrated significantly negative correlations with an altitude. However, both soil nitrifying and denitrifying bacteria were positively correlated with the altitude and total nitrogen. In summary, soil organic and liable carbon, total nitrogen, carbon mineralization accumulation, nitrifying and denitrifying bacteria increased along an altitudinal gradient, however, the contrary patterns were demonstrated among bacteria, fungi, actinomyces and azotobacter.
altitude;soil carbon mineralization;soil microbial population;liable carbon
X172
A
1000-6923(2018)05-1811-07
2017-09-28
国家自然科学基金资助项目(41771548)
* 责任作者, 副教授, lijunjian@sxu.edu.cn
李君剑(1975-),男,山西汾阳人,副教授,博士,主要从事土壤生态学研究.发表论文35篇.
