杧果炭疽病菌3个果胶裂解酶基因序列特征及受漆酶基因Lac1影响的分析
2018-05-14李鸿鹏钟昌开吴秋玉张艳杰张贺
李鸿鹏 钟昌开 吴秋玉 张艳杰 张贺


摘 要 果胶裂解酶是炭疽菌在侵染寄主过程中降解寄主细胞壁的一类重要水解酶。本研究在杧果炭疽病菌中克隆获得了3个果胶裂解酶基因Cgpel1、Cgpel2和Cgpel3,DNA全长分别为1 037、1 498、1 089 bp,cDNA全长分别为975、1 380、978 bp,分别编码324、459、325个氨基酸,均含1个果胶裂解酶保守结构域,均有典型的信号肽,不存在跨膜结构。其二级结构中α-螺旋分别占16.05%、20.26%、16.92%,延伸链分别占28.09%、21.79%、29.54%,β-转角分别占5.86%、8.93%、7.38%,无规则卷曲分别占50.00%、49.02%、46.15%。Cgpel1、Cgpel2和Cgpel3的氨基酸序列分别与C.tofieldiae(KZL77240.1)、草莓炭疽菌(C. gloeosporioides)(XP-007274932.1)、C.incanum(KZL84476.1)果胶裂解酶序列相似度在93%以上。qRT-PCR分析发现,3个果胶裂解酶基因在整个侵染过程中均持续高效表达,但在漆酶基因Lac1敲除突变体中,表达均下降約90%。可见Cgpel1、Cgpel2和Cgpel3是果胶裂解酶基因家族成员,序列差异较大,在侵染过程中起着重要的作用,且其表达受漆酶基因Lac1影响。
关键词 杧果;胶孢炭疽菌;果胶裂解酶基因;漆酶基因Lac1中图分类号 S432.1 文献标识码 A
DOI10.3969/j.issn.1000-2561.2018.12.019
杧果(Mangifera indicaL.)是重要的热带、亚热带果树,享有“热带果王”的盛誉,具有很高的经济价值[1]。杧果炭疽病是杧果生产上发生最普遍、为害最严重的一种病害,在世界杧果种植区内广泛发生,也是贮藏期的重要病害之一,发病重的果园减产达60%~70%,胶孢炭疽菌[Colle?totrichum gloeosporioides(Penz.)Sacc.]是其主要致病菌[2-3]。该病原菌寄主范围十分广泛,大多数热带亚热带果树是其重要寄主,还可侵染蔬菜、花卉、中草药及各种经济作物[4]。
炭疽菌主要靠黑色素化附着胞直接侵入寄主,再分泌大量细胞壁降解酶扩展[5]。DHN黑色素在植物病原真菌中普遍存在,它在附着胞膨压产生上起着关键的作用,其合成途径中有多个关键的酶基因,其中漆酶是催化最后一步黑色素合成的酶[6]。在杧果炭疽病菌中,漆酶Lac1在菌丝的生长、发育、分化、黑色素的沉着、产孢和漆酶的分泌以及对寄主的致病力等方面起着重要的调控作用[7]。
果胶酶是细胞壁降解酶中的一类,也是分解果胶质的酶总称,按其作用方式和底物类型可分为原果胶酶、果胶酯酶、裂解酶、多聚半乳糖醛酸酶[8]。许多病原真菌、细菌在侵染植物的过程中都可以产生果胶裂解酶(pel),裂解高度酯化的果胶,破坏植物组织的完整性,使薄壁细胞组织软化[9-10],从而达到降解植物细胞壁的效果。Erwinia chrysanthemi strain 3937有5个Pel基因PLa~PLe,其中只有PLa、PLd和PLe对病原菌的致病力有影响,且PLe比PLa和PLd的作用更强,PLe突变后失去了侵染能力[11]。辣椒疫霉菌(Phytophthora capsici)的12个候选果胶裂解酶基因中只有Pcpel1、Pcpel16、Pcpel20能够明显影响致病力[9]。可见果胶裂解酶基因多以基因家族形式存在,同一病菌中不同的果胶裂解酶基因作用存在明显差异。将杧果炭疽病菌接种至PD培养液,28 ℃、160 r/min振荡培养12 d的过程中,均能检测到果胶酶活性[12]。有关果胶裂解酶基因的分子特征及其在杧果炭疽病菌侵染寄主过程中的作用尚未见报道。因此本研究拟利用同源克隆技术,从杧果炭疽病菌(C. gloeosporioides)中克隆pel,分析其在不同侵染时段的表达情况,及与另一个侵染关键基因漆酶基因Lac1的关系,为探究果胶裂解酶基因在杧果炭疽病菌致病过程中的作用、揭示杧果炭疽病菌的致病分子机制打下基础。
1 材料与方法
1.1 材料
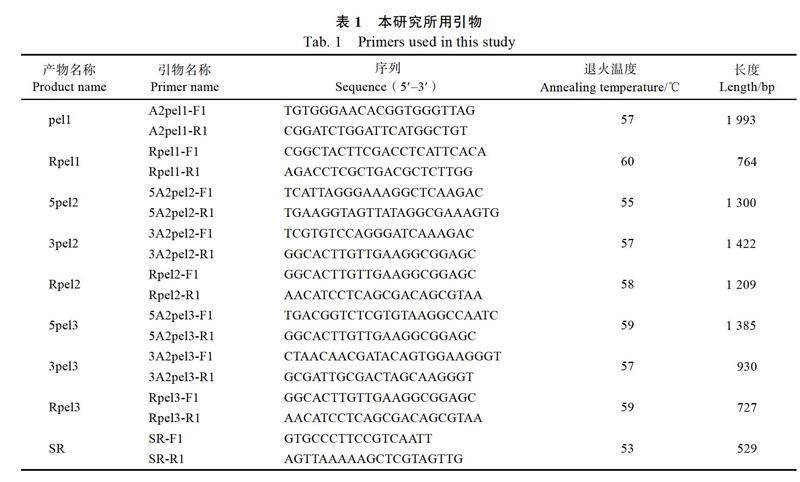
1.1.1 材料和引物杧果炭疽病菌胶孢炭疽菌[C. gloeosporioides(Penz.)Sacc.] A2菌株由中国热带农业科学院环境与植物保护研究所热带果树课题组提供。台农芒嫩叶采自海南大学热带农林学院基地。漆酶基因Lac1敲除突变体由本实验室提供。本研究所用引物如表1所示。
1.1.2 主要试剂 真菌gDNA提取试剂盒和凝胶回收试剂盒购于Omaga公司;RNA提取试剂盒和TIANScript cDNA First-Stand Kit购于TIANG?ENE公司;E. coli DH5α、10× PCR Buffer、dNTPs、TaqDNA酶、2Taq-Mixture、DNA marker DL2000等均购于TaKaRa公司;荧光定量PCR试剂盒2xTransStart? Tip Green qPCR SuperMix购于北京全式金生物技术科技有限公司,其他试剂均为常规试剂。
1.2 方法
1.2.1Cgpel1、Cgpel2和Cgpel3基因DNA序列的扩增 根据橡胶树胶孢炭疽菌HBCg01全基因组信息查找果胶裂解酶基因序列,设计引物A2pel1-F1/R1、5A2pel2-F1/R1、3A2pel2-F1/R1、5A2pel3-F1/R1和3A2pel3-F1/R1(表1),用于扩增杧果炭疽病菌3个果胶裂解酶基因DNA序列。PCR体系为50 μL:10× PCR Buffer 5.0 μL,dNTPs 4.0 μL,引物(10 μmol/L)各1.0 μL,模板1.0 μL,TaqDNA酶1.0 μL,ddH2O 37 μL。PCR程序:94 ℃ 4 min;94 ℃ 30 s,Tm 45 s,72 ℃ 2 min,35个
1.2.2Cgpel1、Cgpel2和Cgpel3基因cDNA的扩增 以杧果炭疽病菌gRNA反转录后的总cDNA为模板,用引物对Rpel1F1/R1、Rpel2F1/R1、Rpel3F1/R1(表1)扩增3个果胶裂解酶基因cDNA序列,其余同1.2.1节。
1.2.3Cgpel1、Cgpel2和Cgpel3序列特征分析 利用DNAssist 1.0软件比对分析所獲得的Cgpel1、Cgpel2、Cgpel3基因DNA和cDNA序列,拼接获得全长,并找出起始密码子、终止密码子、内含子、外显子。用Primer 5.0软件推测其编码的氨基酸序列。用ProtParam软件分析其理化性质,包括分子量、等电点等。用MEGA 4.1软件构建进化树。用Singal 3.0软件分析信号肽位置。用TMHMM软件分析跨膜区。用SOPMA软件分析二级结构。
1.2.4Cgpel1、Cgpel2和Cgpel3在杧果炭疽病菌侵染台农芒嫩叶过程中的表达分析 以SR作为内参基因,用扩增Rpel1、Rpel2和Rpel3的引物(表1)检测Cgpel1、Cgpel2和Cgpel3在杧果炭疽病菌侵染台农芒嫩叶过程中的相对表达量。参考张贺等[13]的方法制备浓度为4×106个/mL的分生孢子悬浮液,用于接种嫩叶。用清水洗净嫩叶,浸泡于1% NaClO溶液中15 min后,用超纯水冲洗3次,置于保鲜盒中,保持RH 100%,用束针刺伤嫩叶后滴加20 ?L孢子悬浮液,28 ℃下恒温黑暗培养。接种后的取样时间点为0、6、12、24、36、48、72 h,以0 h的为对照。取样方法:用直径为1 cm的打孔器取12个接种点,按照不同的时间点取样后至于液氮速冻,-80 ℃保存、备用。每组处理重复3次。
参照Tiangen RNA Kit中的方法提取不同时间点的病原菌总RNA,参照TIANScript cDNA First-Stand Kit试剂盒完成第一链cDNA的合成,反转录后的cDNA稀释10倍,备用。实时荧光定量PCR反应体系为10 μL:ddH2O 2 μL,cDNA 1 μL,引物各1 μL,2xTransStart? Tip Green qPCR SuperMix 5 μL,Rotor-Gene Q型实时荧光PCR仪检测。数据分析运用2–ΔΔCt法进行数据计算,实时荧光定量PCR仪系统导出各样品的Ct值,以接菌0 h的为对照。
PCR反应程序为:94 ℃ 30 s;94 ℃ 5 s,60 ℃ 15 s,72 ℃ 10 s,40个循环;在72 ℃时收集荧光信号。
1.2.5Cgpel1、Cgpel2和Cgpel3基因在漆酶基因Lac1敲除突变体的表达分析 提取漆酶基因Lac1敲除突变体和野生型A2总RNA,以在野生型A2中的表达量为对照,以SR作为内参基因,分析Cgpel1、Cgpel2和Cgpel3在漆酶基因Lac1敲除突变体中的相对表达量。其余参照1.2.4节。每组处理重复3次。
2 结果与分析
2.1 Cgpel1、Cgpel2和Cgpel3基因DNA片段的扩增
