土壤中高产蛋白酶菌株的筛选鉴定及发酵条件优化
2018-05-10杨绍青闫巧娟龚思怡江正强
耿 芳,杨绍青,闫巧娟,刘 军,龚思怡,江正强*
(1.中国农业大学 食品科学与营养工程学院,北京 100083;2.中国农业大学 工学院,北京 100083)
蛋白酶(protelytic enzymes,EC 3.4)是催化肽键水解的一类酶,在食品、饲料、医药、化工等行业有着广泛的应用[1]。与动、植物来源蛋白酶相比,微生物蛋白酶具有培养简便、易批量生产等优点[2],因此,微生物蛋白酶的研究一直备受关注。大部分商业中性和碱性蛋白酶都来源于细菌,目前,已报道产胞外蛋白酶的细菌种类很多[3],常见的有芽孢杆菌(Bacillus)、假单胞菌(Aeruginosa)等。
粘质沙雷氏菌(Serratiamarcescens),是一种革兰氏阴性兼性厌氧菌,主要存在于沿海盐碱土壤中,能够产生红色非扩散性色素[4]。据报道,粘质沙雷氏菌所产的碱性蛋白酶不仅可以有效消除芽孢菌酶制剂的弊端,而且还具有一定的抗肿瘤活性[5]。赵海霞等[6]从土壤中筛选出一株产胶原蛋白酶的粘质沙雷氏菌,天然酶活为21.19 U/mL,具有较强消化牛皮的能力。WAN M等[7]从二甲基亚砜中分离出一株产耐有机试剂蛋白酶的菌株粘质沙雷氏菌MH6,该菌在一定浓度的亲水有机溶剂中仍可保持稳定活性。这些研究表明粘质沙雷氏菌蛋白酶在工业应用上具有广阔的前景。此外,粘质沙雷氏菌产生的灵菌红素及灵杆菌脂多糖在医药行业有着重要的价值[8],其作为菌种制剂又具有抑癌、抗菌等作用,在农作物的灾害防治上效果显著[9-10]。同时粘质沙雷氏菌也是一种高产脂肪酶、几丁质酶的菌种,能够有效降解水产品副产物[11],具有很大的开发潜力[12]。
国外对粘质沙雷氏菌蛋白酶的研究主要集中在酶的纯化与性质上。NAM M S等[13]纯化得到大小约为50 kDa的碱性蛋白酶,比酶活力为289 U/mg;SALARIZADEH N等[14]在温泉附近分离到一株产金属蛋白酶的粘质沙雷氏菌,其在55℃、pH 10.0条件下酶活力最高;SALAMONE PR等[15]从土壤中得到的粘质沙雷氏菌来源蛋白酶,其酶活力高达865 U/mL。国内报道的野生型粘质沙雷氏菌发酵产蛋白酶的水平普遍偏低[16],且关于粘质沙雷氏菌产蛋白酶发酵条件优化的研究较少。通常,液体发酵培养基中的一些成分如碳源、氮源等对菌株产蛋白酶很大影响[17]。本试验从海南土壤中筛选得到一株高产蛋白酶的粘质沙雷氏菌(Serratiamarcescens)CAUH83,对菌株CAUH83形态观察、生理生化试验及16SrDNA序列分析,并对其产蛋白酶的发酵条件(碳源、氮源、pH、温度、表面活性剂及发酵时间)进行优化,以期为该菌株所产蛋白酶的应用提供一定的理论基础。同时,虽然蛋白酶的工业生产技术已经较为成熟,但为降低成本,对其进行发酵优化以提高蛋白酶活力仍然有待进行深入研究。
1 材料与方法
1.1 材料与试剂
1.1.1 材料
土壤样品:海南火山群公园;粘质沙雷氏菌(Serratia marcescens)CAUH83:筛选自土壤样品。
1.1.2 试剂
酪蛋白、Folin-酚:美国Sigma公司;蛋白胨、酵母提取物:英国Oxoid公司;Triton X-100基因组提取试剂盒:天根生化科技有限公司;r Taq DNA聚合酶:日本Takara公司;2 000bp DNA Marker:北京博迈德生物技术有限公司;其他试剂均为国产分析纯。
1.1.3 培养基
菌种筛选固体培养基:脱脂奶粉2.0%,琼脂2.0%。
种子培养基:酵母提取物0.5%,蛋白胨1.0%,葡萄糖2.0%,氯化钠0.5%,磷酸氢二钾0.7%,磷酸二氢钾0.3%。
菌种发酵液体培养基:酵母提取物0.5%,蛋白胨1.0%,葡萄糖2.0%,氯化钠0.5%,磷酸氢二钾0.7%,磷酸二氢钾0.3%。
培养基灭菌条件:高压蒸汽灭菌121℃、20 min。
1.2 仪器与设备
HZQ-F160全温振荡培养箱:江苏太仓实验设备厂;DK-S24恒温水浴锅:上海精宏实验设备有限公司;TU-1800PC紫外可见分光光度计:北京普析通用仪器有限责任公司;B104光学生物显微镜:上海光学仪器一厂;T100型梯度聚合酶链反应(polymerasechain reaction,PCR)扩增仪:美国伯乐公司;JY-E型电泳仪:北京六一仪器厂;JY02G型凝胶成像仪:上海小原科技有限公司。
1.3 方法
1.3.1 样品的采集
拨开土壤表面,向下约10 cm处收集土样。
1.3.2 产蛋白酶菌株的筛选
(1)菌株的初筛
取0.1 g土样溶于1 mL无菌水中,稀释10倍后取200μL混合液涂布于菌种筛选固体培养基上,于37℃静置培养2d。
(2)菌株的复筛
挑取呈现透明圈的菌株,于菌种筛选固体培养基上37℃静置培养2 d。经分离纯化后挑取单菌落接入菌种发酵液体培养基中,于200 r/min、37℃振荡条件下培养2 d。发酵液经离心后取上清液测定蛋白酶活力。
1.3.3 菌株的鉴定
菌株形态与生理生化鉴定:将分离纯化好的菌株于筛选固体培养基上37℃静置培养2 d,观察该菌菌落形态。挑取单菌落涂片,经革兰氏染色后在光学显微镜下观察细胞形态,并对该菌进行生理生化的鉴定[4]。
分子生物学鉴定:采用细菌基因组DNA提取试剂盒提取菌株的基因组DNA。以菌株的基因组DNA为模板,采 用 16S rDNA通用引物27F(5‘-AGAGTTTGATCCTG GCTCAG-3‘)、1492R(5‘-GGTTACCTTGTTACGACTT-3‘),聚合酶链式反应扩增菌株的16SrDNA的部分基因序列。PCR扩增条件:94℃预变性5 min;94℃变性30 s,55℃退火30 s,72℃延伸2 min,34次循环;72℃再延伸10 min。PCR扩增产物经过琼脂糖凝胶电泳验证后,送至生工生物工程有限公司进行测序。将测序结果在GenBank数据库中进行BLAST比对,使用MGEA 6.0软件中的邻接(neighborjoining,NJ)法构建系统发育树[18]。
1.3.4 蛋白酶活力的测定
粗酶液的制备:将单菌落接入种子培养基进行活化,取1%的种子液接种至液体培养基中,于200 r/min、37℃条件下振荡培养2 d,发酵结束后取发酵液于10 000 r/min离心10 min,得到的上清液即为粗酶液。
蛋白酶活力的测定:采用GB/T 23527—2009《蛋白酶制剂》法。将1 mL粗酶液与1 mL底物(酪蛋白溶液)混合均匀,于40℃反应10 min,然后加入2 mL三氯乙酸终止反应,12 000 r/min条件下离心3 min。在1 mL上清液中加入5 mL Na2CO3以及1 mL Folin-酚试剂,混合均匀后于40℃保温20 min,采用紫外可见分光光度计于波长660 nm处测定吸光度值。以先加入三氯乙酸终止反应的一组为对照组。
蛋白酶活力的定义:1个酶活力单位定义为每分钟水解酪蛋白产生1μg酪氨酸所需的酶量(U)。
1.3.5 粘质沙雷氏菌CAUH83发酵产蛋白酶条件优化
采用单因素试验优化粘质沙雷氏菌CAUH83的产蛋白酶发酵条件,考察碳源、氮源、培养基初始pH值、培养温度以及培养时间对菌株产蛋白酶的影响。
在初始发酵培养基的基础上,考察不同碳源(酒糟、麸皮、葡萄糖、蔗糖、乳糖、脱脂棉和可溶性淀粉)及碳源添加量(1%、2%、3%、4%、5%、6%、7%)对粘质沙雷氏菌CAUH83产蛋白酶的影响。考察不同氮源(酵母提取物、蛋白胨、大豆蛋白胨、豆粕、大豆粉、干酪素、脱脂奶粉、尿素和硫酸铵)及氮源浓度(1%、2%、3%、4%、5%、6%、7%)对产蛋白酶的影响。设计2因素(碳源、氮源浓度)3水平正交试验,以确定产蛋白酶最佳碳氮比。调节培养基的初始pH值,考察培养基初始pH值(3、4、5、6、7、8、9、10)对产蛋白酶的影响,考察不同培养温度(20℃、25℃、30℃、32℃、35℃、40℃)对产蛋白酶的影响。最后在上述最适条件下培养该菌株,确定最适发酵时间。
1.3.6 聚丙烯酰氨凝胶电泳及酶谱分析
参照LAEMMLIUK[19]的方法,将粗酶液与含有十二烷基硫酸钠(sodiumdodecyl sulfate,SDS)和二巯苏糖醇(dithiothreitol,DTT)的缓冲液混合均匀后于沸水中加热5min,冷却离心后上样,进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)检测蛋白酶纯度,并计算其分子质量。
蛋白酶酶谱:参照JIANGWB等[20]的方法。在电泳分离胶中加入0.1%的明胶作为底物,在含有2.5%Triton X-100的50mmol/LTris-HCl缓冲液中反应30min后,置于50mmol/L Tris-HCl缓冲液中,37℃反应2 h。
2 结果与分析
2.1 菌株的筛选、分离和鉴定
2.1.1 蛋白酶产生菌的筛选结果
从不同土壤中筛选获得多株产蛋白酶的菌株,测定其蛋白酶活,结果见表1。由表1可知,通过比较这几株菌的菌落直径、水解圈比值以及发酵液的酶活力,筛选出一株高产蛋白酶菌株CAUH83,该菌株初筛蛋白酶活力达到126U/mL。

表1 蛋白酶产生菌的筛选结果Table 1 Screening results of protease-producing strains
2.1.2 形态学观察及生理生化鉴定结果
根据菌株CAUH83的菌落形态、革兰氏染色以及生理生化反应结果,按《常见细菌系统鉴定手册》进行初步分类[21],该菌产红色菌落,菌落表面光滑,边缘不规则,不透明,且粘性易挑起(见图1),菌落形态与赵海霞等[6]报道的产胶原蛋白酶的沙雷氏菌(Serratiamarcescens)描述一致。
根据分离菌的菌落和菌体形态、革兰氏染色以及生理生化反应,按《常见细菌系统鉴定手册》进行初步分类[19],生理生化特征见表2。由表2可知,菌株革兰氏染色、产芽孢、精氨酸双水解酶及葡萄糖、木糖、棉子糖、阿拉伯糖利用试验结果呈阴性;脂酶、DNA酶、明胶液化及蔗糖、乳糖利用试验结果呈阳性。

图1 菌株CAUH83菌落形态Fig.1 Colonial morphology of strain CAUH83

表2 菌株CAUH83生理生化特性鉴定结果Table 2 Identification results of physiological and biochemical characteristics of strain CAUH83
2.1.3 分子生物学鉴定

图2 菌株CAUH83基于16s rDNA基因序列系统发育树Fig.2 Pylogenetic tree based on 16S rDNA sequences of strain CAUH83
提取菌株CAUH83的基因组DNA,经PCR扩增、测序后得到全长约1500bp的基因序列,在美国国家生物技术信息中心(ntional center of biotechnology information,NCBI)数据库中进行基本局部比对搜索工具(basic local alignment search tool,BLAST)比对可知,该菌株序列与粘质沙雷氏菌的16SrDNA序列同源性高达99%。利用软件MEGA 6.0构建系统发育树,结果见图2。由图2可知,该菌株CAUH83与粘质沙雷氏菌(Serratiamarcescens)聚于一支,亲缘关系最为接近。因此结合形态学、生理生化鉴定结果,鉴定菌株CAUH83为一株粘质沙雷氏菌(Serratia marcescens)。
2.2 菌株CAUH83发酵产蛋白酶条件优化
2.2.1 碳源对菌株CAUH83产蛋白酶的影响
不同碳源(含量2%)对菌株CAUH83发酵产酶的影响结果见图3。由图3可知,当以蔗糖作为碳源时,菌株CAUH83发酵产蛋白酶活性最高,蛋白酶活力为162.9 U/mL,其次为葡萄糖(159.6 U/mL)和可溶性淀粉(126.3 U/mL)。发酵过程中,蔗糖先分解为葡萄糖和果糖。果糖可以直接转化为1,6-二磷酸果糖,进入糖酵解过程。因此,相对于葡萄糖,果糖更易代谢,能够更加有效地提高菌体量[22]。因此,选择蔗糖作为最适碳源。

图3 不同碳源对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.3 Effects of different carbon resources on protease activities produced by S.marcescens CAUH83

图4 蔗糖添加量对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.4 Effects of sucrose addition on protease activities produced by S.marcescens CAUH83
以蔗糖为碳源,研究不同蔗糖添加量对菌株产酶的影响结果见图4。由图4可知,蔗糖添加量对产酶的影响较为明显,蔗糖添加量由1%增加到3%时,酶活逐渐升高,当蔗糖添加量为3%时,蛋白酶的酶活力最高为238.8 U/mL,继续增加蔗糖添加量时,酶活力呈缓慢下降趋势。因此选择3%作为蔗糖的最适添加量。
2.2.2 氮源对菌株CAUH83产蛋白酶的影响
不同氮源(含量2%)对菌株产蛋白酶的影响结果见图5。由图5可知,氮源对菌株CAUH83产酶的影响大于碳源[23],有机氮源更有利于菌株产酶,以大豆粉作为氮源时,蛋白酶的酶活力最高为378.4U/mL,其次为脱脂奶粉与豆粕。从营养成分来看,大豆粉蛋白质含量高达40%,并且为优质蛋白。此外,大豆粉还富含各种氨基酸、矿物质和维生素等营养成分。因此,选择大豆粉为最适氮源。

图5 不同氮源对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.5 Effects of different nitrogen resources on protease activities produced by S.marcescens CAUH83
确定大豆粉为最适氮源后,进一步研究大豆粉的添加量对菌株产蛋白酶的影响,结果见图6。由图6可知,当大豆粉添加量为3%时,蛋白酶酶活力最高,当大豆粉的含量继续增加时,酶活力开始逐渐下降。因此,大豆粉最适添加量为3%。

图6 大豆粉添加量对粘质沙雷氏菌CAUH83产蛋白酶的影响Fig.6 Effects of soybean flour addition on protease activities produced by S.marcescens CAUH83
2.2.3 培养基初始pH值对菌株CAUH83产蛋白酶的影响
液体发酵培养基不同初始pH值对粘质沙雷氏菌CAUH83产酶的影响结果见图7。由图7可知,随着初始pH值升高,酶活力逐渐增大;当初始pH值>6时,酶活力开始下降。故该菌产酶最适初始pH值为6。中性偏酸环境更适宜该菌株产蛋白酶,与文献报道一致[24]。
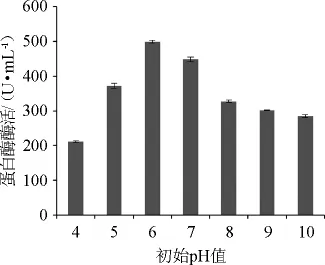
图7 培养基初始pH值对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.7 Effects of initial pH on protease activities produced by S.marcescens CAUH83
2.2.4 培养温度对菌株CAUH83产蛋白酶的影响
培养温度对菌株CAUH83产蛋白酶的影响结果见图8。由图8可知,培养温度对菌株CAUH83发酵产蛋白酶的影响较大。当菌株在30℃培养时,蛋白酶活性最高,达到1 862.2 U/mL。当培养温度继续升高时,酶活力迅速下降,因此,选择30℃作为发酵培养基的培养温度。

图8 培养温度对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.8 Effects of incubation temperature on protease activities produced by S.marcescens CAUH83
郝林华等[26]研究粘质沙雷氏菌的生长温度时发现,高温对该菌的生长非常不利,在26~30℃时该菌生长效果最好,高于30℃时该菌的耐受性下降,产菌量也偏低。菌株CAUH83最适培养温度为30℃,且培养温度对该菌分泌蛋白酶的影响较为显著。受温度影响较大的还有假单胞菌Pseudomonas sp.W7,康传红等[27]在优化该菌产低温蛋白酶的发酵条件时发现,在20℃时该酶蛋白酶活力达到最大值,继续升高培养温度,酶活力逐渐下降,温度升至30℃时,酶活力仅为最大酶活力的15%。
2.2.5 培养时间对菌株CAUH83产蛋白酶的影响
确定以上最适条件后,进一步考察发酵时间对粘质沙雷氏菌CAUH83发酵产蛋白酶的影响结果见图9所示。由图9可知,菌株CAUH83在优化后的最适发酵条件下,12h已开始产酶,发酵48 h时酶活力达到最高水平,之后继续延长培养时间,酶活力逐渐下降。因此,最佳培养时间为48 h。

图9 培养时间对粘质沙雷氏菌CAUH83产蛋白酶酶活的影响Fig.9 Effects of incubation time on protease activities produced by S.marcescens CAUH83
2.3 SDS-PAGE及酶谱分析
粘质沙雷氏菌CAUH83在最适条件下培养48 h,其粗酶液的SDS-PAGE及蛋白酶酶谱如图10所示。由图10可知,菌株CAUH83分泌的主要蛋白分子质量约为50 kDa。酶谱结果表明,该酶为蛋白酶。作为野生菌株,菌株CAUH83的发酵粗酶液中蛋白质组分相对简单,SDS-PAGE电泳条带接近单一,这一特征有利于该酶的纯化。

图10 粘质沙雷氏菌CAUH83发酵液SDS-PAGE电泳图谱及酶谱图Fig.10 SDS-PAGE electrophoretogram and zymogram of fermentation liquid from S.marcescens CAUH83
3 结论
本研究从海南土壤样品中筛选得到一株高产蛋白酶的粘质沙雷氏菌(Serratiamarcescens)。采用单因素试验优化得到了该菌株液体发酵产蛋白酶的最优条件为:蔗糖添加量30%、大豆粉添加量30%、初始pH值6.0、培养温度30℃、培养时间48 h。在此发酵条件下,蛋白酶的酶活力达到1 932.3 U/mL,较优化之前提高了15.3倍,为目前已报道野生型粘质沙雷氏菌产蛋白酶的最高水平。SDS-PAGE及酶谱分析表明,该菌所产的蛋白酶具有蛋白酶活性且蛋白质分子质量约50 kDa。本研究通过研究高产蛋白酶菌株粘质沙雷氏菌的发酵条件,为进一步研究其产酶特性以及在工业领域的应用提供理论依据。
参考文献:
[1]NIYONZIMA FN,MORE S.Detergent-compatible proteases:microbial production,properties,and stain removal analysis[J].Prep Biochem Biotech,2015,45(3):233-258.
[2]方海红,胡好远,黄红英,等.微生物碱性蛋白酶的研究进展[J].微生物学通报,2002,29(2):7-59.
[3]KAMAN WE,HAYSJP,ENDTZ H P,et al.Bacterial proteases:targets for diagnosticsand therapy[J].Eur J Clin Microbiol,2014,33(7):1081-1087.
[4]黄文芳.粘质沙雷氏菌(Serratia marcescens)的研究Ⅰ—Serratia marcescens 9-2菌株分离、分类鉴定和形态特征[J].华南师范大学学报:自然科学版,2003(3):108-111.
[5]黄利群,彭珍荣.粘质沙雷氏菌合成碱性蛋白酶的阻遏作用[J].武汉大学学报:自然科学版,1993,29(1):88-92.
[6]赵海霞,赵培培,陈 惠,等.一株产胶原蛋白酶沙雷氏菌的分离及鉴定[J].中国饲料,2012(1):9-11,14.
[7]WAN M,WU B,REN W,et al.Screening,characterization,and cloning of a solvent-tolerant protease from Serratia marcescens MH6[J].J Microbiol Biotechn,2010,20(5):881-888.
[8]郝名慧,楼志华,张 梁,等.一株新粘质沙雷氏菌发酵产红色素及其结构的研究[J].天然产物研究与开发,2007,19(3):439-442.
[9]HEARN W R,WILLIAMS R H,WILLIAMS R P,et al.Separation of prodigiosenes and identification as prodigiosin syntrophic pigment from mutantpairsof Serratiamarcescens[J].Appl Microbiol,1972,24(4):591-595.
[10]WILLIAMSRP.Biosynthesis of prodigiosin,asecondary metaboliteof Serratiamarcescens[J].Appl Microbiol,1973,25(3):396-402.
[11]ZHANGH,FANGJ,DENGY,et al.Optimized production of Serratia marcescens B742 mutants for preparing chitin from shrimp shells powders[J].Int J Biol Macromol,2014,69:319-328.
[12]GAO L,XU JH,LIX J,et al.Optimization of Serratia marcescens lipaseproduction for enantioselective hydrolysisof 3-phenylglycidic acid ester[J].J Ind Microbiol Biot,2004,31(11):525-530.
[13]NAM M S,WHANGK S,CHOISH,et al.Purification,characterization,and properties of an alkaline protease produced by Serratia marcescens S3-R1 inhabiting Korean ginsengrhizosphere[J].J Sci Food Agr,2013,93(15):3876-3882.
[14]SALARIZADEH N,HASANNIA S,AKBARINOGHABIK,et al.Purification and characterization of 50 kDa extracellular metalloprotease from Serratia sp.ZF03[J].Iran J Biotechnol,2014,12(3):18-27.
[15]SALAMONEPR,WODZINSKIRJ.Production,purification and characterization of a 50-kDa extracellular metalloprotease from Serratia marcescens[J].Appl Microbiol Biotechnol,1997,48(3):317-324.
[16]柯 野,伍嘉慧,曾松荣,等.一株产蛋白酶菌株的筛选鉴定及酶学性质分析[J].广东农业科学,2015,42(2):137-141.
[17]SECADESP,ALVAREZ B,GUIJARRO JA.Purification and characterization of a psychrophilic,calcium-induced,growth-phase-dependent metalloprotease from the fish pathogen Flavobacterium psychrophilum[J].Appl Environ Microbiol,2001,67(6):2436-2444.
[18]FELSENSTEIN J.Confidence-limits on phylogenies-an approach using thebootstrap[J].Evolution,1985,39(4):783-791.
[19]LAEMMLIUK.cleavageof structural proteinsduring assembly of head of Bacteriophage-T4[J].Nature,1970,227(5259):680-685.
[20]JIANG W B,LERSA,LOMANIEC E,et al.Senescence-related serine proteasein parsley[J].Phytochemistry,1999,50(3):377-382.
[21]郑朝成,周立祥.污泥中一株产耐高温蛋白酶菌株的分离、鉴定及酶学特性[J].环境科学学报,2012,32(3):577-583.
[22]吴家鑫,王凤寰,王 峥,等.蔗糖与葡萄糖作为辅助碳源对重组Klebsiella pneumoniae发酵生产1,3-丙二醇的影响[J].过程工程学报,2014,14(4):660-663.
[23]KUMAR R S,PRABHU D,SHANKAR T,et al.Optimization of alkalophilic protease production by Pseudomonas aeruginosa isolated from the gut of Penaus monodon[J].World J Fish Marine Sci,2011,3(5):371-375.
[24]许曌昕,曾 辉,曾德勇,等.响应面法优化解淀粉芽孢杆菌产中性蛋白酶发酵条件[J].中国酿造,2017,36(4):78-82.
[25]黄小龙,彭 可,周双清,等.粘质沙雷氏菌产几丁质酶发酵条件的研究[J].生物技术,2010,20(3):64-66.
[26]郝林华,陈靠山,牛德庆,等.粘质沙雷氏菌(Serratia marcescens)发酵培养基优化的研究[J].海洋科学进展,2006,24(1):66-73.
[27]康传红,田亚新,李秀凉,等.Pseudomonas sp.W7产低温蛋白酶培养基及培养条件的优化[J].食品科学,2015,36(19):163-169.
