纳板河自然保护区土壤酶对不同海拔、坡向的响应
2018-05-05申佳艳李小英温婧媛
申佳艳, 李小英, 袁 勇, 温婧媛
(西南林业大学 环境科学与工程学院, 昆明 650224)
土壤酶活性是土壤中生物学活性的总体现,土壤酶利于土壤养分的归还与转化,是森林植被与土壤养分间的重要桥梁[1-2],对土壤空间异质性的形成具有重要生态学意义。目前关于森林土壤酶活性研究方面取得了较好的进展,并且大量的研究结果显示土壤酶活性不仅能对土壤理化性质及养分含量产生作用,也会对土壤理化性质、土壤类型、植被、气候等综合环境因素做出响应[3]。在森林生态系统中,土壤酶的功能多样性能够更加完整的体现在土壤理化性质、养分含量、动植物群落变化等方面。森林土壤酶活性变异规律的研究对于探索森林生态系统过程与功能起到了综合性作用[4]。
近年来纳板河流域国家级自然保护区对动植物资源现状与保护、部分区域土壤理化性质等方面做了系列研究[5-8],但基于保护区特殊的地理环境和气候特点所做的土壤酶活性研究鲜见。因此本研究采用相关分析、通径分析、主成分分析等方法对西双版纳纳板河流域国家级自然保护区(以下简称纳板河保护区)的典型区域土壤酶活性及养分状况进行分析,旨在探讨在亚热带气候条件下,土壤酶活性与各养分指标及环境因素间的关系以及将其作为保护区森林土壤肥力指标的可行性。
1 材料与方法
1.1 研究区概况
纳板河流域国家级自然保护区,位于西双版纳傣族自治州中北部,景洪市与勐海县的接壤地带,地理坐标为100°32′—100°44′E,22°04′—22°17′N,地处热带北缘向南亚热带过渡的区域,具有大陆性气候与海洋性气候的优点,总体气候类型属北热带湿润气候,年均温18~22℃,最冷月均温12~16℃,最热月均温22~26℃,年降雨量1 100~1 600 mm,雨量充沛而集中,干湿季分明。保护区垂直高差较大,山地气候垂直带明显。海拔高度800~900 m及以下属北热带,800(900)~1 400(1 500)m属南亚热带,1 400(1 500)m以上的山地属中亚热带。
1.2 土样采集与处理
于2016年4月(旱季)在保护区北部过门山管理站附近已经建设完成的20 hm2季风常绿阔叶林大样地内进行采样。样地海拔在1 000~1 600 m,部分地段可下延到600 m左右,高差较大,垂直带较明显,故采样按照海拔梯度分两个坡向(阴坡、阳坡)进行。在海拔600~1 500 m每150 m为一个梯段,每个梯段内按垂直高度均匀挖三个剖面,剖面间距为50 m左右,每个土壤剖面分0—20 cm,20—40 cm和40—60 cm三个层次取样,共42个土壤剖面,126个土样,包括鲜土(用于测定土壤酶活性)、混合土样(土壤化学性质分析),每种均为126个。采样的同时用GPS记录采样点经纬度及坡向、坡度,并记录样地林分结构和立地特点,取好的混合土壤样品带回实验室,置于通风清洁无氨气、灰尘等污染的室内自然风干后过筛,装密封袋保存用于测定土壤基本理化性质。鲜土样品密封后置于2~4℃的密封保温箱中带回实验室进行2℃恒温冷藏,用于测定土壤酶活性。具体样地情况见表1。
1.3 测试方法
有机质用K2CrO7氧化还原滴定法;全氮用高氯酸—硫酸消化法;全磷用NaOH碱熔—钼锑抗比色法;水解氮用扩散吸收法测定;速效磷用盐酸—氟化铵浸提—钼锑抗比色法;速效钾用NH4OAC—火焰光度计法测定;pH值用pH值酸度计测定。
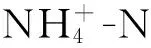
1.4 分析方法
不同海拔、坡向及土层间酶活性差异特征采用单因素方差分析(ANOVA)法,以检验土壤酶活性在不同海拔、不同坡向、不同土壤层次间差异显著性;土壤酶活性与养分间关系采用通径分析;主成分分析用于论证保护区土壤酶是土壤肥力的重要指标。所有数据均为每个海拔梯段内三个剖面分层平均值,通过Excel 2007和SPSS 21.0软件进行统计分析并且所有分析及假设均采用α=0.05为显著性水平进行检验。
2 结果与分析
2.1 不同海拔土壤养分及酶活性特性
由图1可知,各土层土壤有机质含量随海拔升高均表现出显著地垂直分布特征。阳坡环境下有机质含量最大值在1 500 m海拔处,达到109.8 g/kg。阴坡环境下,有机质含量在海拔1 050~1 500 m随海拔升高呈波动增加趋势,最大值出现在1 200 m海拔区域,达到100.49 g/kg,低于阳坡环境最大值。图1)示,土壤全氮含量随海拔升高呈现出较为显著的垂直变化规律。阳坡环境下,600~900 m海拔间表层土壤全氮含量随海拔显著升高,1 350 m海拔处达到最大值0.449 g/kg;阴坡环境下,表层土壤全氮含量在600~900 m海拔间逐渐增加,1 050~1 500 m海拔之间波动增加,1 500 m海拔区域到达最大值0.525 g/kg,中下层含量变化规律与表层基本一致。阳坡环境表层土壤全氮含量最大值在中高海拔区域,阴坡含量最大值在最高海拔区域。
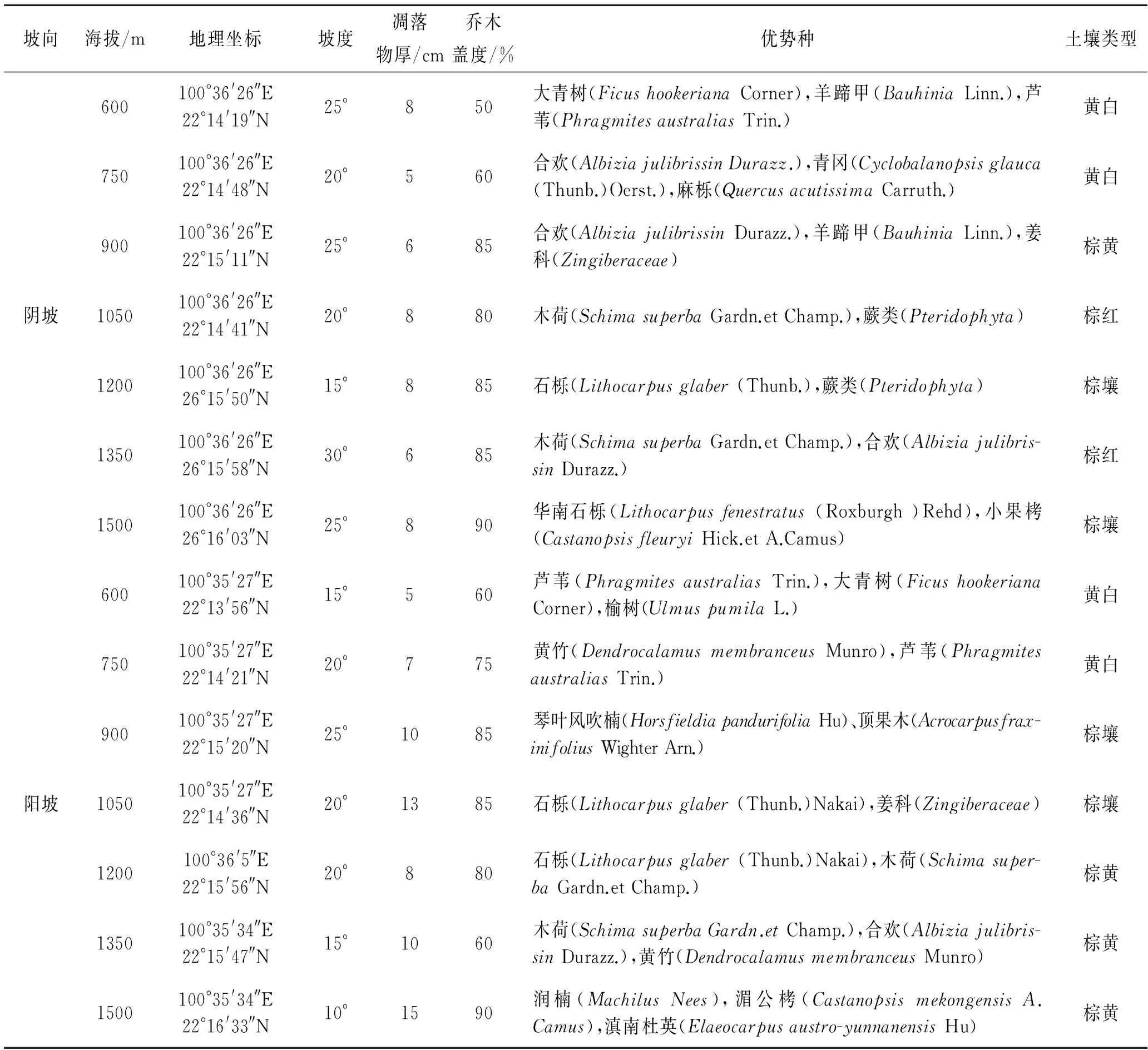
表1 样地概况
由图2可知,土壤水解氮含量随海拔升高垂直变化特征明显,阳坡表层土壤水解氮含量随海拔升高逐渐增大,1 500 m海拔处达到最大值342.4 mg/kg,中下层水解氮含量在600~1 050 m海拔间显著增加,在1 200 m海拔处略微下降,之后回升;阴坡表层水解氮含量在600~1 200 m海拔区域显著增加,1 350 m海拔处略微减小,之后回升并在1 500 m海拔处达到最大值440.4 mg/kg。中下层含量随海拔变化规律与表层基本一致,增加趋势弱于表层。图2显示,不同坡向土壤速效磷含量随海拔升高变化趋势差异较大,阳坡表层土壤速效磷含量呈现由900 m海拔处向两端减小趋势,中下层含量在600~900 m海拔之间随海拔升高逐渐增加,之后呈现减小—增加—减小的波动性变化;阴坡表层含量在900 m海拔处达到最大,之后随海拔减小,1 350 m处含量最低,1 500 m海拔处回升,中下层变化特征与表层类似。
由图3可知,不同坡向土壤蛋白酶活性随海拔升高变化趋势差异明显。阳坡表层酶活性随海拔升高总体增加,600~900 m区域随海拔变化平缓,之后随海拔急剧升高,1 500 m海拔处酶活性最强。中下层酶活性在600~1 350 m变化规律与表层一致,1 350~1 500 m与表层趋势相反;阴坡表层酶活性在600~750 m海拔间急剧升高,900 m海拔处出现大幅减小,900~1 200 m海拔间大幅度升高,1 200 m处达到最大。中层土壤蛋白酶活性与表层随海拔变化规律一致;图3显示,不同坡向表层土壤过氧化氢酶活性随海拔升高变化均较小,中下层不同坡向环境随海拔变化有所差异,阳坡环境呈现由中间向两端减小,阴坡环境呈增大—减小—增大的变化趋势,1 050 m海拔处活性最强。
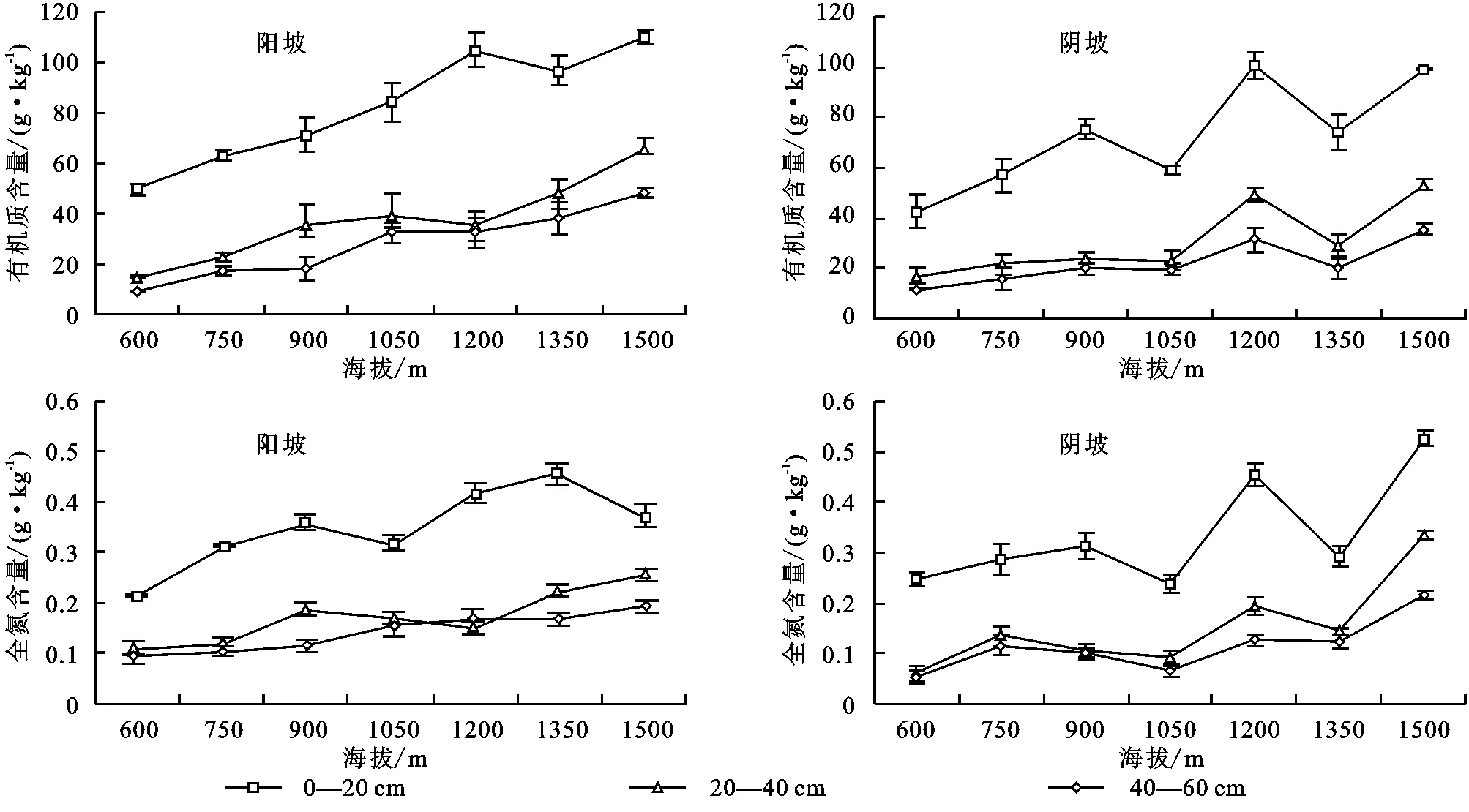
图1 不同坡向土壤有机质、全氮随海拔变化折线
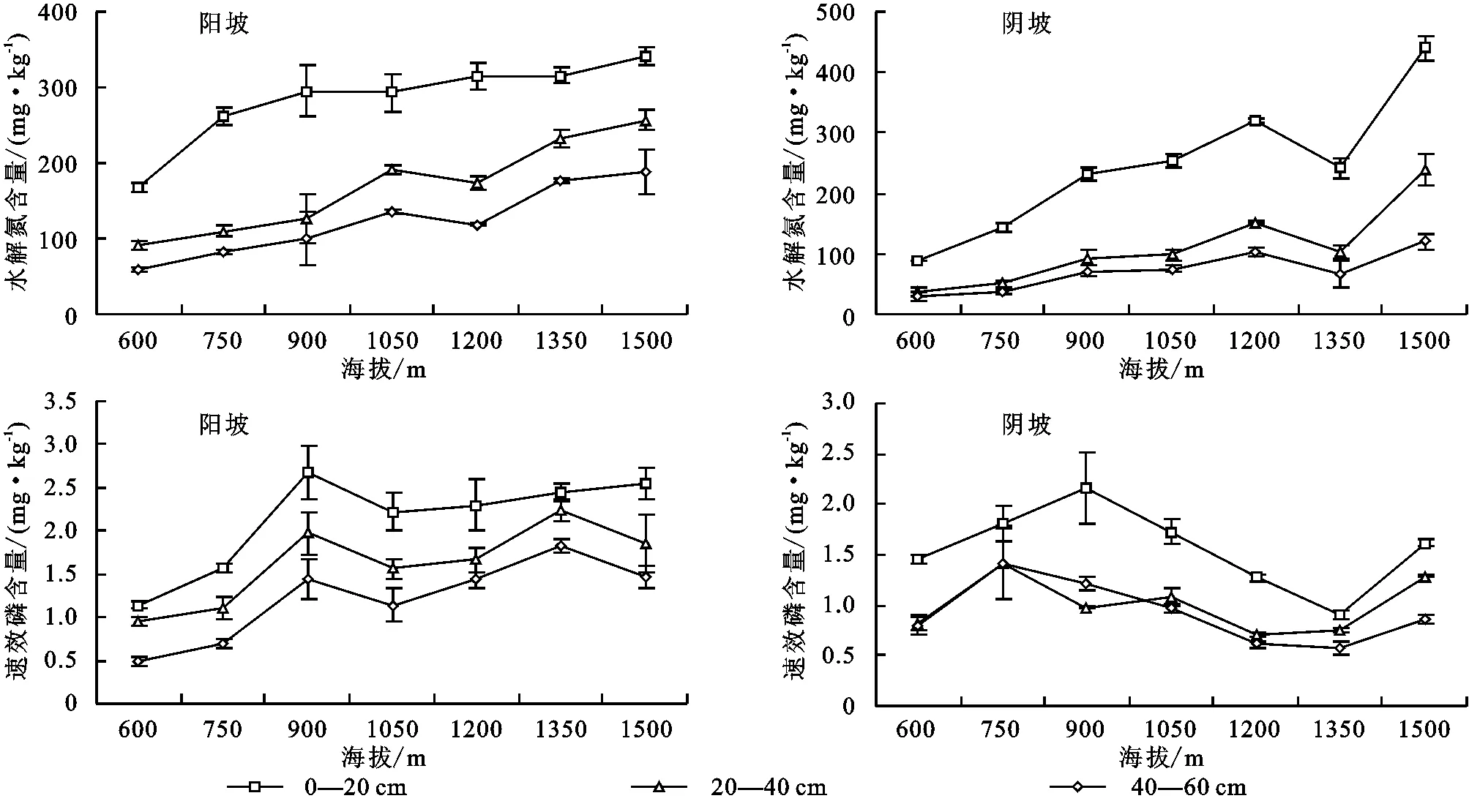
图2 不同坡向土壤水解氮、速效磷含量随海拔变化折线
由图4可知,不同坡向土壤脲酶、蔗糖酶活性随海拔变化规律差异较大,阳坡脲酶活性呈由中间向两端减小趋势;阴坡表层酶活性在600~750 m海拔间显著升高,750~900 m海拔间急剧减小,900 m处最低,之后回升,1 500 m海拔处酶活性最高。中下层酶活性随海拔变化趋势与表层一致,变幅较表层小;图4B显示,阳坡表层土壤蔗糖酶活性从900 m海拔向两端减小,1 050 m海拔处活性最低,之后有所回升。中下层变化规律与表层一致,变幅较小;阴坡表层土壤蔗糖酶活性随海拔升高整体上升。中下层土壤酶活性在600~1 050 m海拔间无明显变化,1 050~1 500 m海拔间显著增加。

图3 不同坡向土壤蛋白酶、过氧化氢酶活性随海拔变化折线
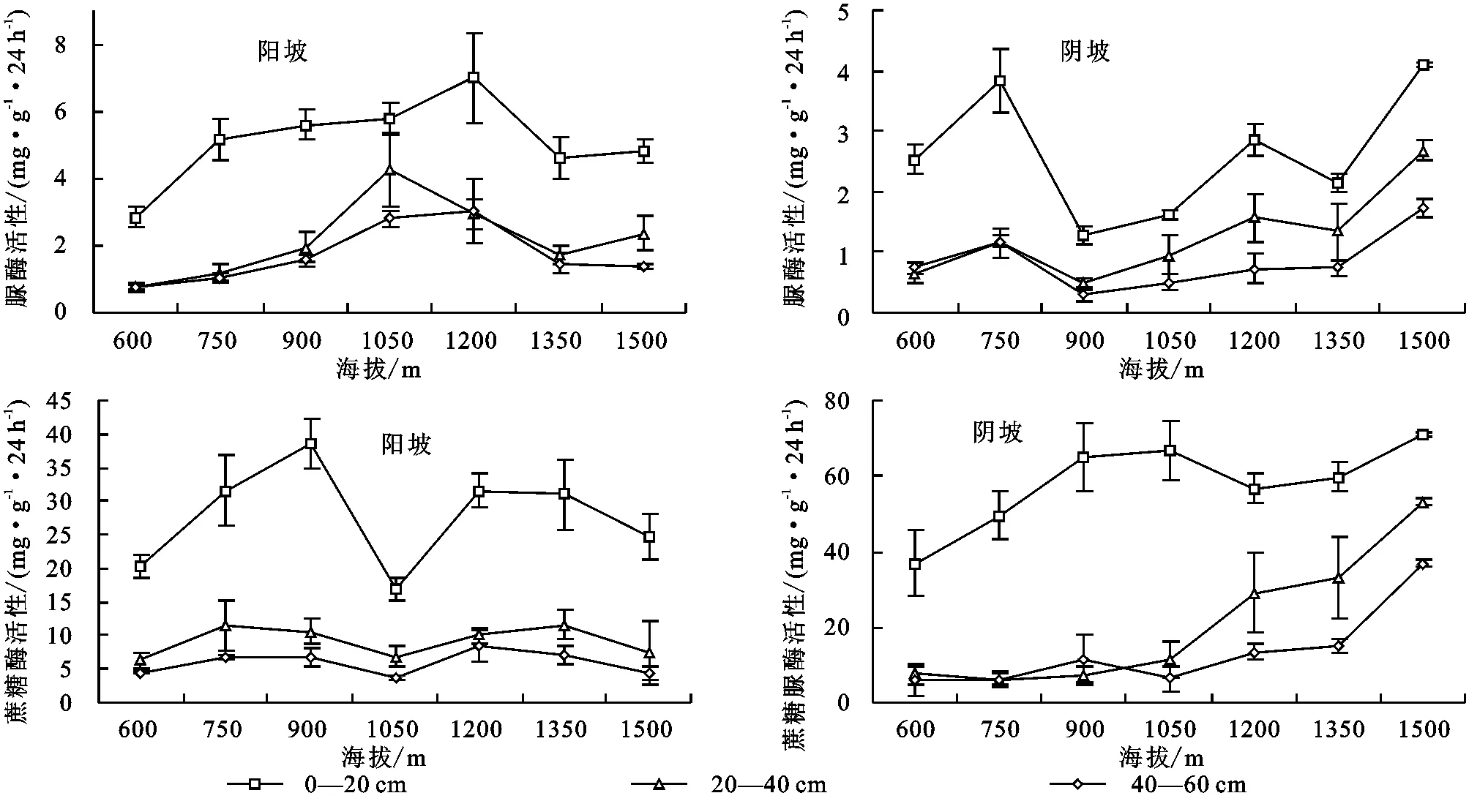
图4 不同坡向土壤脲酶、蔗糖酶活性随海拔变化折线
2.2 不同坡向土壤养分及酶活性变化特征
由图5可知,阴阳坡分异对土壤有机质含量影响不大,阳坡土壤有机质含量略高于阴坡,差异不显著(p>0.05);阴阳坡分异对全氮含量影响较小,阳坡各土层全氮含量略大于阴坡,差异不显著(p>0.05);坡向对不同土层水解氮含量影响有所不同,表层土壤水解氮含量阳坡略高于阴坡,差异不显著(p>0.05)。中下层含量阳坡显著高于阴坡(p<0.01);坡向对土壤速效磷含量影响显著(p<0.01),阳坡>阴坡。
由图6可知,土壤蛋白酶活性受坡向影响较小,阴坡各土层蛋白酶活性略高于阳坡,差异不明显(p>0.05);脲酶活性受坡向影响较大,各土层脲酶活性阳坡>阴坡,差异显著(p<0.05);过氧化氢酶活性受坡向影响较小,各土层过氧化氢酶活性阴坡>阳坡,差异不明显(p>0.05);蔗糖酶活性受坡向影响较大,各土层蔗糖酶活性阴坡>阳坡,差异极显著(p<0.01)。
变异系数大小反映了保护区土壤各指标随海拔、坡向的空间变异程度。由表2可知,土壤有机质、全氮、速效磷、速效钾、水解氮在不同土层中的变异系数均在0.1~1.0之间,属于中等变异,说明了保护区土壤养分分布相对均一的特点;土壤酶活性随海拔、坡向体现出不同程度变异性,不同酶活性随海拔、坡向变异程度不同。

图5 土壤有机质、全氮、水解氮、速效磷坡向柱形图
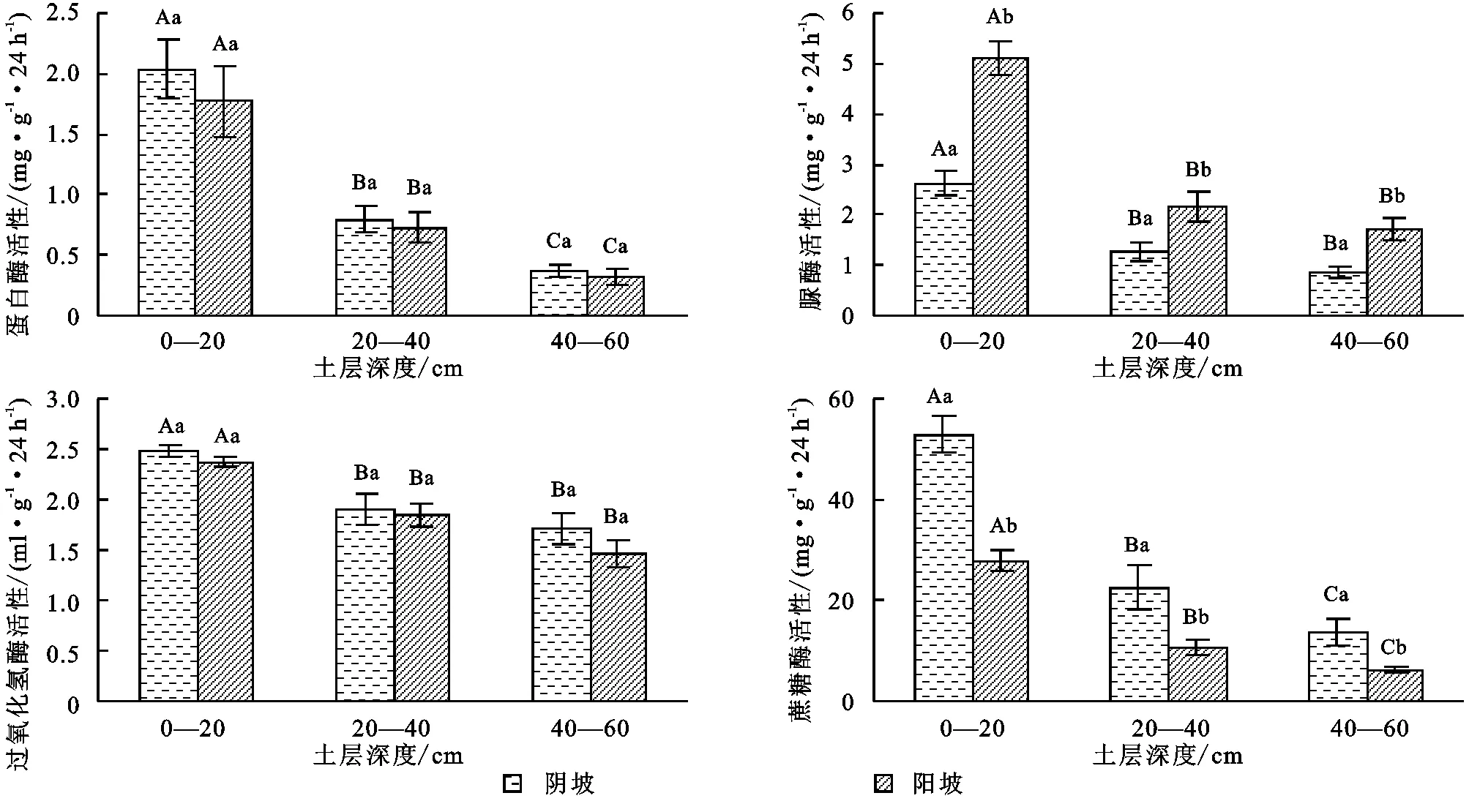
图6 土壤蛋白酶、脲酶、过氧化氢酶、蔗糖酶活性坡向柱形图
2.3 土壤酶活性与土壤肥力因素关系分析
2.3.1 相关性分析 相关研究表明土壤肥力在一定程度上受土壤酶活性影响[9],土壤肥力水平与酶活性之间相关关系显著。通过相关性分析得到土壤酶活性与主要肥力因子间的相关系数,结果表明(表3)蛋白酶、脲酶与土壤有机质、全氮、全磷、速效磷、速效钾、水解氮之间相关关系极显著(p<0.01);过氧化氢酶与土壤有机质、全氮、速效钾、水解氮之间相关性极显著(p<0.01),而与全磷、速效磷之间相关性不显著(p>0.05);蔗糖酶与土壤有机质、全氮、速效钾、水解氮之间相关性极显著(p<0.01),与全磷和速效磷之间相关性显著(p<0.05)。蛋白酶和脲酶活性与所有肥力因子之间均存在极显著相关关系(p<0.01),过氧化氢酶与蔗糖酶只与部分肥力因子间存在显著相关关系(p<0.01),与部分肥力因子间相关关系不明显,不能完全代表土壤肥力水平。这个结果说明用蛋白酶和脲酶活性作为纳板河保护区土壤肥力指标具有一定可行性。
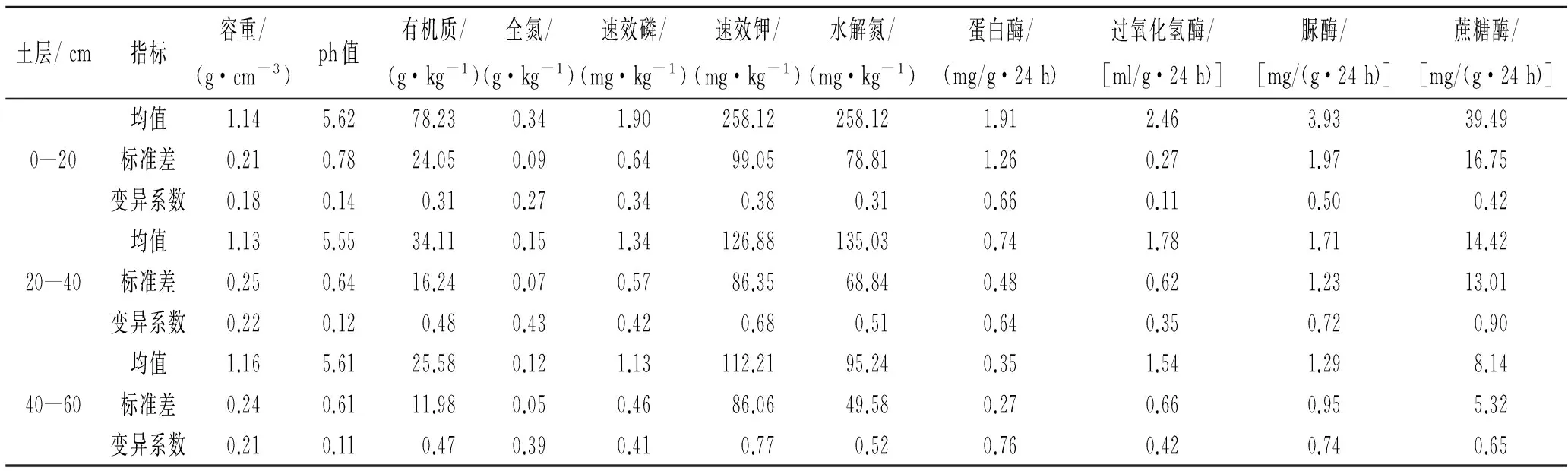
表2 土壤各指标变异特征分析
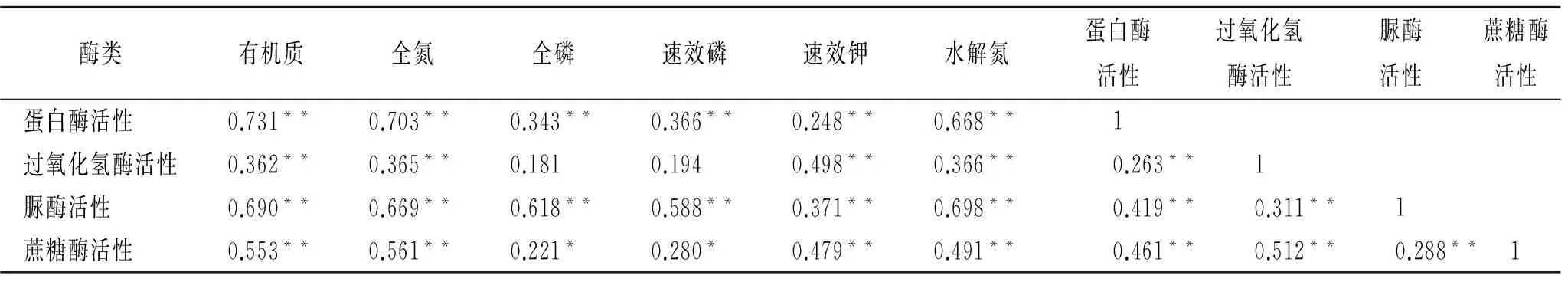
表3 土壤酶活性与主要养分相关关系分析
**.在.01水平(双侧)上显著相关;*.在0.05水平(双侧)上显著相关。
2.3.2 通径分析与多元回归分析 多个变量的系统中,两变量之间的关系在一定程度上会受其他变量影响,为准确说明两变量之间的相关关系,采用通径分析与多元回归分析相结合的方法对所有指标进行处理与分析。多元回归方程用于描述随机变量在多个回归因子中的平均变化规律,通径分析则是标准化的多元线性回归分析。将蛋白酶、脲酶与主要肥力因子的测定结果进行回归,得到两个标准多元回归方程:
p=0.600X1+0.249X2-0.383X3+0.141X4-0.095X5+0.068X6
U=0.117X1+0.073X2+0.455X3-0.255X4+0.099X5+0.368X6
式中:P,U表示标准化后的蛋白酶、脲酶活性;X1表示有机质;X2表示全氮;X3表示全磷;X4表示速效磷;X5表示速效钾;X6表示水解氮;X1—X6表示标准化后的土壤养分含量。回归方程中的系数为直接通径系数,反映了土壤各肥力因子对蛋白酶和脲酶活性的直接影响。间接通径系数来源于方程中的直接通径系数与各个肥力因子之间相关系数的乘积[10-12],结果见表4。

表4 土壤主要肥力因子对土壤酶活性的通径系数分析表
注:划横线的数据为直接通径系数。
直接通径系数反映出保护区土壤各主要肥力因子对酶活性的直接影响程度,间接通径系数反映出主要肥力因子通过其他肥力因子对土壤酶活性产生的间接影响力大小,间接通径系数能够更加科学、客观的体现出土壤肥力因子对酶活性的影响程度。由表4可以得出各肥力因子对蛋白酶活性的直接影响力大小(按照绝对值排列)为:有机质>全磷>全氮>速效磷>水解氮>速效钾;对脲酶活性的直接影响力(按照绝对值排列)为:全磷>水解氮>速效磷>有机质>速效钾>全氮;土壤水解氮对蛋白酶的直接影响力很小(直接通径系数为0.068),但它通过有机质对蛋白酶活性产生的间接影响系数是直接系数的7.9倍,说明土壤水解氮主要通过间接作用影响蛋白酶活性。同理,速效钾对蛋白酶的间接通径系数是直接通径系数的26.7倍,速效钾主要通过有机质对蛋白酶活性产生间接影响。对于脲酶而言,除全氮以外的肥力因子对脲酶活性的直接通径系数绝对值均大于间接通径系数绝对值,说明这些肥力因子主要通过直接作用影响脲酶活性。
2.3.3 主成分分析 主成分分析是一种通过降维的方法,在损失很少信息的前提下,用较少的几个综合变量来代替原始变量的统计方法,这几个综合指标能够尽可能的表现出原始变量的信息,并且彼此之间相互独立[13-15]。为了论证纳板河自然保护区土壤酶能够作为土壤肥力的重要指标,将土壤酶与主要肥力因子做了主成分分析。由表5可以看出共提出三个主成分,第一主成分的方差贡献率最大,为57.89%,加上第二主成分方差贡献率13.55%和第三主成分方差贡献率10.27%后累计方差贡献率达到82.16%(大于80%),包含了原始变量80%的信息,因此这三个主成分基本能够反映出保护区土壤肥力的变异信息。
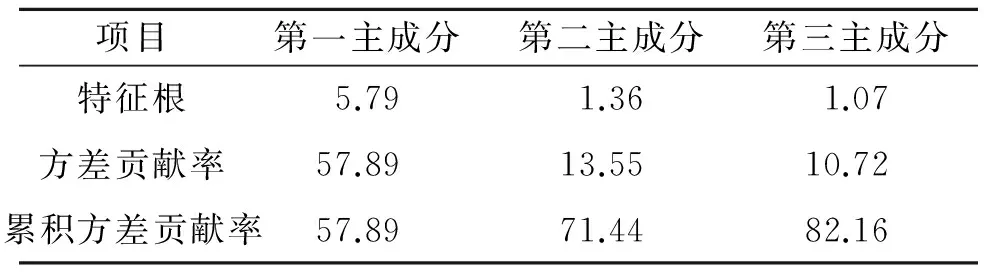
表5 测试土壤主成分特征根
注:特征根表示的是累积方差贡献。
通过计算得出各个肥力因子在各主成分上的载荷见表6,第一主成分主要综合了蛋白酶、脲酶、有机质、全氮、全磷、水解氮、速效磷的变异信息,第二主成分综合了蔗糖酶、过氧化氢酶的变异信息。第一主成分的累计方差贡献率最大,对土壤肥力水平的高低起到主要作用,通过载荷来看,蛋白酶与脲酶都集中在第一主成分上,蛋白酶和脲酶足以体现保护区土壤肥力,因此将蛋白酶和脲酶作为纳板河自然保护区土壤肥力指标是可行的。
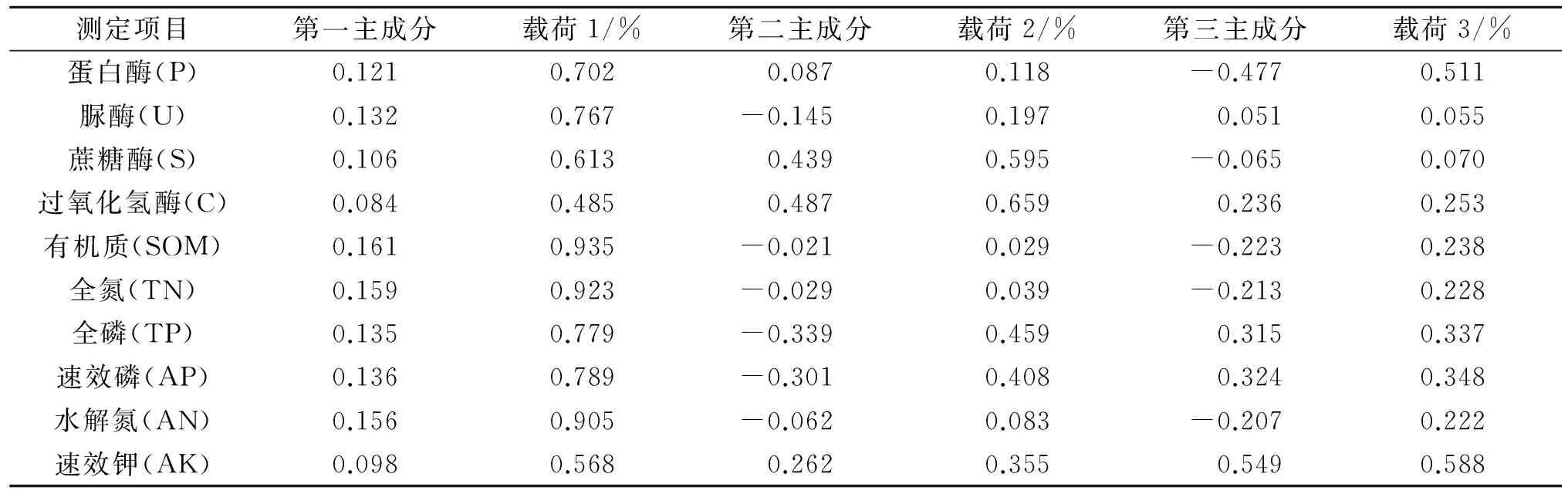
表6 测试土壤主成分特征向量
注:载荷1,2,3分别表示各主成分上承载的各因子的方差百分率。
3 结论与讨论
纳板河自然保护区海拔对有机质含量影响较显著,阳坡高海拔区域>低海拔区域,阴坡最大值在中高海拔区域。这与彭新华等[16]提出的有机质含量随海拔升高向复杂方向发展,最大值不出现在山顶的结论一致,而与杨克凤[17]、车宗玺[18]等的研究有所差异,说明土壤有机质含量随海拔升高而增大的趋势是针对一定的地理环境和研究尺度而言。纳板河保护区土壤有机质含量随海拔的变化与该区域由海拔升高呈现的由干热河谷地带竹林、灌木林到常绿阔叶林的植被变化有密切关系,发源于干热河谷地带的土壤有机质显著低于中高海拔区域;全氮含量随海拔升高变化趋势复杂,阳坡最大值出在1 350 m海拔区域内,说明1 350 m海拔区域土壤温度、湿度等方面的优势较高低海拔区域都更利于氮的积累,阴坡在1 200 m海拔处波动较大,可能是1 200 m海拔处于植被交替地带的原因。水解氮受海拔影响显著,最大值出现在受人为干扰较小的1 500 m海拔阳坡区域。速效磷含量受海拔影响显著,且不同坡向环境随海拔变化差异较大。
阳坡环境土壤蛋白酶活性随海拔升高总体呈递增趋势,高海拔>低海拔,这与杨万勤[19]、金裕华[20]等的研究结果相一致。阴坡环境土壤蛋白酶活性随海拔升高波动明显,活性最大值在中高海拔区域。表层土壤过氧化氢酶活性受海拔变化影响较小,中下层不同坡向环境随海拔变化有所差异,阳坡环境呈现由中间向两端减小,阴坡环境呈增大—减小—增大的变化趋势。这与金裕华[20]等对武夷山不同海拔植被带土壤酶活性的研究结果不大一致,主要原因可能是武夷山研究区域随海拔变化植被带分布特征较该研究区域明显。不同坡向土壤脲酶活性随海拔变化差异较大,阳坡脲酶活性最大值在中高海拔区域,阴坡呈中间低两端高的趋势,最大值在最高海拔区域。不同坡向土壤蔗糖酶活性随海拔升高变化特征差异较大,阳坡随海拔呈增大—减小—增大的变化趋势,阴坡酶活性随海拔升高整体上升。不同坡向各指标随海拔变化呈现差异的原因主要是保护区坡向差异引起土壤垂直带谱及同类土壤分布高度的差异[21],阳坡为棕黄壤(600~1 050 m)→棕壤(1 050~1 500 m),阴坡为黄壤(600~900 m)→棕黄壤(900~1 200 m)→棕壤(1 200~1 500 m),这种土壤分布高度的差异使得各指标随海拔变化规律出现坡向差异。
阴阳坡分异对土壤有机质、全氮含量影响不大,阳坡略高于阴坡。这与阴阳坡光照强弱引起的土壤温度差异对土壤动物、微生物活动的影响有关,阳坡土壤地温变异较大,土壤微生物活动受抑制,有机质和氮素分解转化慢,利于积累。阴坡土壤温度相对稳定,阴凉潮湿的土壤环境利于微生物活动,促进土壤有机质及氮素分解[22-23];对水解氮、速效磷含量影响显著,阳坡>阴坡;土壤蛋白酶、过氧化氢酶、蔗糖酶活性均为阴坡>阳坡,这与邱莉萍[24]等的研究结论相一致,研究区阴坡环境植被生长相对旺盛,凋落物丰富,微生物繁衍迅速利于酶活性增强。各土层脲酶活性阳坡>阴坡,这与土壤氮素含量变异规律一致,体现出土壤脲酶活性在森林生态系统中与氮素分解间的密切联系,即较高的土壤脲酶活性是土壤供氮水平的反映。
不同海拔、坡向表层土壤各养分含量及酶活性均明显高于中下层,土层间养分含量的差异性体现出保护区土壤垂直变异特点。土壤微生物代谢产酶能力随土层加深逐渐减弱是表层土壤酶活性较中下层更具优势的主要原因[25]。
土壤酶活性与土壤肥力因子间相关关系明显,土壤养分含量通对植物和微生物而间接作用于土壤酶,形成了土壤养分与酶活性之间不同程度的相关性[4]。各土壤酶之间存在的不同程度相关关系体现了土壤中各种酶促反应既相互独立又相互影响,相互关联的特点,各种酶的相互作用促进了土壤空间异质性的形成,这与张仕艳[26]、郑文教[27]等的研究结果相一致。
通径分析表明纳板河保护区对蛋白酶活性起主导作用的因子是有机质、全氮、全磷,速效磷和水解氮为次要因子。全磷、速效磷、水解氮为影响脲酶活性的主要因子,有机质、全磷、速效钾为次要因子。通径分析能够更为全面、客观的体现各个肥力因子与酶活性之间直接和间接地相关关系,使土壤酶活性与化学性质之间的相互作用更加明了[28-31]。主成分分析为蛋白酶和脲酶能够作为保护区土壤肥力指标提供了依据与强有力的理论支撑,但基于土壤酶活性高度的变异性,因此将其作为土壤肥力指标的可行性还需根据不同的环境条件进行更加深入的研究。
参考文献:
[1] 陶宝先,张金池,愈元春,等.苏南丘陵地区森林土壤酶活性季节变化[J].生态环境学报,2010,19(10):2349-2354.
[2] 蒋智林,刘万学,万方浩,等.紫茎泽兰与非洲狗尾草单,混种群落土壤酶活性和土壤养分的比较[J].植物生态学报,2008,32(4):900-907.
[3] 邱丽萍,刘军,王益权,等.土壤酶活性与土壤肥力的关系研究[J][J].植物营养与肥料学报,2004,10(3):277-280.
[4] 杨万勤,王开运.森林土壤酶的研究进展[J].林业科学,2004,40(2):152-159.
[5] 刘峰,樊国盛,陶国达.纳板河保护区野生药用植物资源研究[J].环境科学导刊,2010,29(4):21-24.
[6] 晁增华.纳板河流域国家级自然保护区森林资源现状及发展对策[J].山东林业科技,2010,40(3):104-106.
[7] 苏惠,杨云.纳板河自然保护区石斛属植物资源现状与保护对策[J].林业调查规划,2006,31(5):100-102.
[8] 王东升,施春蕾,刘峰,等.中国云南纳板河国家级自然保护区苔藓植物研究[J].热带亚热带植物学报,2008,16(5):452-465.
[9] 胡建忠.人工沙棘林地土壤酶分布及其与土壤理化性状间关系的研究[J].沙棘,1996,9(2):22-28.
[10] 陶宝先,张金池,愈元春,等.苏南丘陵地区森林土壤酶活性季节变化[J].生态环境学报,2010,19(10):2349-2354.
[11] 赵益新,陈巨东.通径分析模型及其在生态因子决定程度研究中的应用[J].四川师范大学学报:自然科学版,2007,30(1):120-123.
[12] 刘广深,徐冬梅,许中坚,等.用通径分析法研究土壤水解酶活性与土壤性质的关系[J].土壤学报,2003,40(5):756-762.
[13] 李新蕊.主成分分析,因子分析,聚类分析的比较与应用[J].山东教育学院学报,2007,22(6):23-26.
[14] 李新蕊.主成分分析,因子分析,聚类分析的比较与应用[J].山东教育学院学报,2007,22(6):23-26.
[15] 张子龙,王文全,缪作清,等.主成分分析在三七连作土壤质量综合评价中的应用[J].生态学杂志,2013,32(6):1636-1644.
[16] 彭新华,李元沅,赵其国.我国中亚热带山地土壤有机质研究[J].山地学报,2001,19(6):489-496.
[17] 杨克凤.马鬃岭自然保护区土壤碳蓄积的研究[D].安徽芜湖:安徽农业大学,2011.
[18] 车宗玺,刘贤德,车宗奇,等.祁连山青海云杉林土壤有机质及氮素的空间分布特征[J].水土保持学报,2014,28(5):164-169.
[19] 杨万勤,钟章成,陶建平,等.缙云山森林土壤酶活性与植物多样性的关系[J].林业科学,2001,37(4):124-128.
[20] 金裕华.武夷山不同海拔土壤微生物多样性的变化特征[D].南京:南京林业大学,2012.
[21] 蒲玉琳.横断山区北段山地土壤坡向性分异研究[D].成都:四川农业大学,2005.
[22] 郭永龙,毕如田,王瑾,等.华北典型山区忻州不同小生境坡耕地土壤肥力特征[J].水土保持学报,2013,27(5):205-208,218.
[23] 陈颖,雷波,罗承德,等.康定木格措冷杉林土壤有机质水平及其预测[J].水土保持学报,2013,27(4):252-257.
[24] 陈颖,雷波,罗承德,等.康定木格措冷杉林土壤有机质水平及其预测[J].水土保持学报,2013,27(4):252-257.
[25] 万忠梅,宋长春.土壤酶活性对生态环境的响应研究进展[J].土壤通报,2009(4):951-956.
[26] 张仕艳,原海红,陆梅,等.滇西北不同利用类型土壤酶活性及其与理化性质与微生物的关系[J].亚热带水土保持,2010(2):13-16.
[27] 郑文教,王良睦,林鹏.福建和溪亚热带雨林土壤酶活性的研究[J].生态学杂志,1995(6):16-20.
[28] 林绍霞,张清海,张珍明,等.基于通径分析法的土壤性质对茶叶品质的影响研究[J].水土保持通报,2013,33(4):189-194.
[29] 吴永铃,王兵,戴伟,等.杉木人工林土壤酶活性与土壤性质的通径分析[J].北京林业大学学报,2012,34(2):78-83.
[30] 刘广深,徐冬梅,许中坚,等.用通径分析法研究土壤水解酶活性与土壤性质的关系[J].土壤学报,2003,40(5):756-762.
[31] 杨晓娟,廖超英,李晓明,等.榆林沙区樟子松林土壤酶活性与土壤化学性质的通径分析[J].干旱区资源与环境,2013,27(9):109-112.
