黄土丘陵区微地形梯度下草地群落及土壤对氮、磷添加的响应
2018-05-05张少康刘海威
张少康, 刘海威, 焦 峰,
(1.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100; 2.中国科学院 水利部 水土保持研究所, 陕西 杨凌 712100)
人工添加含N,P元素的肥料直接作用于土壤,改善土壤养分状况,影响植物营养元素吸收分配利用,进而改变群落生产力及其生态化学计量特征。目前,人工N,P添加已成为研究大气N沉降增加对土壤养分含量、植物生长限制元素和生态系统生物地球化学循环影响的有效手段[1-2]。各N,P添加试验中,土壤养分含量呈现不同变化趋势,如宁夏荒漠草原进行的野外N添加试验表明添加少量N元素使得土壤全N显著增加,土壤C,P和N/P不存在明显变化规律[3];而对呼伦贝尔草原羊草割草场的研究表明N,P添加对土壤全N含量没有显著影响,土壤全P含量增大[4]。低浓度N,P添加均使群落地上生物量增大,而随施肥浓度的增大地上生物量累积将受到影响,与地上生物量对N,P添加的响应不同,群落地下生物量对N,P添加的响应存在较大差异,目前没有形成统一定论。施加N肥能够显著增大巨桉幼苗的N/P值,缓解巨桉受N元素限制的情况[5]。不同试验背景及环境条件下群落植物和土壤C,N,P,K对N,P添加的响应存在较大差异,且K作为植物生长所必需的重要营养元素在多数研究中没有涉及。
黄土高原丘陵区土壤类型为黄绵土,水土流失严重,且为生态环境脆弱带,各土壤养分含量均较少[6],大气N沉降增加有可能会对该地区生态体统生态化学计量学循环产生较大影响。研究表明,黄土高原丘陵区植物N含量与全球平均水平相近,而P含量低于全球陆生植物,植物N/P值高,说明黄土高原丘陵区植物生长更容易受P元素限制[7],土壤P元素相对匮乏。黄土高原区关于土壤养分状况的研究较多,地形、退耕年限、土地利用方式及人类活动等均对土壤养分状况造成影响[8-9],尤其是地形条件复杂,坡向坡位影响光照、温度、水分等自然条件[10],使得土壤养分条件在时空上均存在不同程度变异,从而影响植物群落分布及生长状况。目前很少有关于黄土丘陵区草本植物群落及土壤N,P,K对N,P增加响应的研究,而黄土丘陵区不同地形条件下草地群落对未来大气N沉降增加的响应情况有待研究。因此本试验在黄土丘陵区安塞综合试验站,以不同坡向坡位条件下草地群落为研究对象,基于N及N+P耦合营养元素添加的野外控制试验,研究草地群落地上及地下生物量、营养元素吸收分配以及土壤养分对N,P添加的响应情况,以期为大气N沉降增加对黄土丘陵区草地群落的影响提供科学参考依据。
1 材料与方法
1.1 样地概况
试验区位于陕北安塞县真武洞镇中国科学院水土保持研究所安塞综合试验站,东经109°13′,北纬36°41′,属典型黄土丘陵区,以黄绵土为主。气候类型为暖温带半干旱气候,年平均气温8.8℃,年平均降水量531.4 mm,雨热同期,且降雨多集中在7月,8月,9月份,多为暴雨。年日照时数为2 395.6 h,日照百分率达54%,全年无霜期为144 d,太阳总辐射量为527 kJ。气温日差较大。植被以牧草、林木和农作物为主。研究区在植被区划上属于森林—草原区,区域内草本植物主要为:禾本科(Poaceae)植物:白羊草(Bothriochloaischaemum)、长芒草(Stipabungeana)、中华隐子草(CleistogenesChinensis)、丛生隐子草(Cleistogenescaespitosa);豆科(Leguminosae)植物:达乌里胡枝子(Lespedezadavurica)、草木樨状黄芪(Astragalusmelilotoides);菊科(Compositae)植物:铁杆蒿(Artemisiasacrorum)、茭蒿(Artemisiagiraldii)、阿尔泰狗娃花(Heteropappusaltaicus);蔷薇科(Rosaceae)植物:菊叶委陵菜(Potentillatanacetiflolia)、二裂委陵菜(Potentillabifurca)等,在水分较好地段均会出现杠柳(Periplocasepium),狼牙刺(Sophoraviciifolia),铁线莲(Clematisfruticosa)等灌木的入侵,形成以中旱生草本植物占绝对优势的植物群落。
1.2 样地布设与N,P添加
本试验按照不同坡向(阴坡和阳坡)、坡位(坡下部、坡中部、坡上部、坡顶部)选择典型坡面进行样地布设,样地面积30 m×30 m,每个样地3个重复,样地内设置1 m×1 m样方,并采用单因素随机区组方法进行施肥样方布设,分别为CK处理:对照试验,无人工添加;N处理:施加N肥43.4 g(尿素,含氮率46%,纯氮量20 g);N+P处理:施加N肥43.4 g(尿素,含氮率46%,纯氮量20 g)和P肥21.7 g(过磷酸钙,含磷率46%,纯磷量10 g),每个处理设置3个重复。肥料等分为两份,分两次完成施肥,施肥时间为2014年的8月和2015年4月。2015年8月份进行样地信息采集见表1。

表1 样地信息
1.3 样品采集
1.3.1 生物量采集“收割法” 测定地上生物量,于2015年8月中旬从茎基部分收割样方内草本植物,室内105℃杀青5 min后85℃恒温烘至恒重,称取干重,计算地上生物量。地下生物量测定:样地内随机选10个点,用内径9.0 cm的根钻,按照0—20 cm,20—50 cm,50—80 cm,80—100 cm分层取样,室内用100目筛在流水中冲洗出根系,85℃烘干至恒重后称取干重。地下生物量(g/m2)=平均每根土芯根系干重/[π×(0.09/2)2]。
1.3.2 土壤、植物样品采集 每个样地内S形随机选取5个样点,取0—20 cm土样,带回实验室,自然风干后,研磨,过100目尼龙筛备用。采集样方内所有植物种的健康、完整叶片,晾干后装入自封袋中带回实验室,85℃烘干,研磨并过筛备用。样地内S形随机选取5个样点,用内径9.0 cm的根钻钻取0—100 cm土体,带回室内,冲洗出植物根系,烘干后研磨过筛备用。
1.4 样品养分含量测定
本研究中植物叶片N,P,K含量基于群落水平,计算时采用群落植物叶片养分含量的加权平均值,权重因子为物种重要值。物种重要值计算公式为:物种重要值=(相对盖度+相对多度+相对频度)/3
叶片及根系养分含量测定:开氏法测定叶片全N含量;钼蓝比色法测定叶片全P含量;火焰光度计法测定全K。土壤养分含量测定:重铬酸钾容量外加热法测定有机C;浓硫酸和混合加速剂消煮—凯氏定氮仪测定土壤全N;氯化钾浸提—流动分析仪测定速效N;浓硫酸—高氯酸消煮后,钼锑抗比色法测定全P;碳酸氢钠浸提—紫外分光光度计法测定速效P;醋酸铵浸提—火焰光度计法测定速效K。测定方法参照《土壤农化分析》[11]。
1.5 数据分析
用Excel软件进行数据预处理后,用SPSS 17.0对生物量和相关生态化学计量学特征值进行方差分析;F检验显著时,用最小差异显著法(LSD)进行均值间多重比较(α=0.05),最后Sigmaplot 10.0作图。
2 结果与分析
2.1 草地群落生物量对N,P添加的响应
由图1看出,不同地形条件下,与CK处理组相比,N处理组和N+P处理组草地群落地上生物量均增加,并且N+P处理组比N处理组群落生物量增加幅度大,CK处理组平均地上生物量为158.41 g/m2,变异系数为25.96%,N组和N+P组地上生物量分别比CK组增加24.21%和43.92%。方差分析显示,阳坡坡下部、上部和阴坡坡上部两施肥处理组与CK处理组相比地上生物量均显著增加(p<0.05),其他地形增加不显著(p>0.05)。群落地下根系生物量(图2)与地上生物量相反,根系生物量在施肥后总体呈下降趋势,阳坡坡下部N处理组地下生物量稍有增加;阴坡坡上部和阴坡坡中部施肥处理组地下生物量都略有增加;而阳坡坡中部、下部、坡顶和阴坡坡下部施肥处理地下生物量显著减小(p<0.05);CK处理组的平均地下生物量为731.80 g/m2,变异系数为22.40%,N处理组和N+P处理组平均地下生物量分别比CK处理组减少12.19%和8.53%。
2.2 草地群落化学计量特征对N,P添加的响应
2.2.1 叶片化学计量特征对N,P添加的响应 由图3看出,基于黄土丘陵区施肥控制试验,CK处理、N处理和N+P处理叶片N含量取值范围分别为17.31~30.96 g/kg,23.42~39.81 g/kg,25.19~41.35 g/kg,平均值分别为24.84 g/kg,28.97 g/kg,31.27 g/kg,变异系数分别为19.83%,19.62%和17.95%;平均叶片N含量的变化趋势为N+P处理组>N处理组>CK处理组,N处理和N+P处理叶片N含量分别为CK处理的117%和126%,方差分析表明阳坡坡下部和坡顶叶片N含量施肥处理组显著增加(p<0.05),其他地形增加不显著(p>0.05)。CK处理、N处理和N+P处理叶片P含量平均值分别为1.61 g/kg,1.50 g/kg,1.95 g/kg,其中阳坡施肥处理叶片P含量增加,与CK处理组相比N处理组和N+P处理组分别增加19.13%和36.46%,但无显著差异(p>0.05);阴坡N处理叶片P含量减少31.58%,N+P处理组增加1.32%,均无显著差异(p>0.05)。除阳坡坡中部外,施肥处理叶片K含量均增加,但增加趋势不显著(p>0.05)。不同地形条件下CK处理、N处理和N+P处理平均叶片N/P值分布范围分别为12.58~24.28,16.67~4.82,12.56~23.07,平均值分别为16.83,19.21,16.24,各处理组平均叶片N/P值总体呈现N>CK>N+P趋势,无显著差异(p>0.05)。
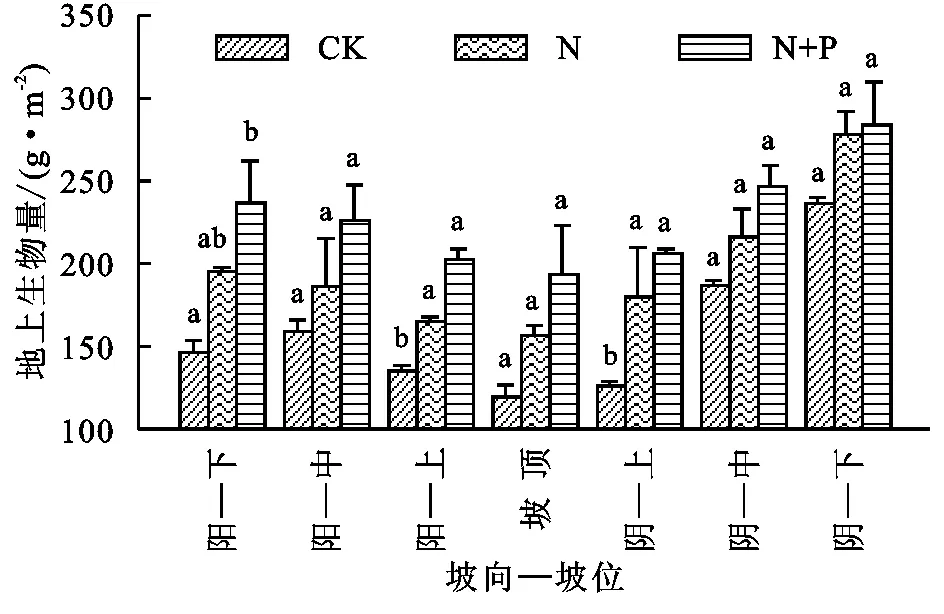
注:阳—下、阳—中、阳—上、阴—上、阴—中和阴—下分别表示阳坡坡下部、阳坡坡中部、阳坡坡上部、阴坡坡上部、阴坡坡中部和阴坡坡下部;CK,N和N+P分别表示CK处理组、N处理组和N+P处理组;不同小写字母表示不同施肥处理组间存在显著差异(p<0.05),下同。
图1处理组间草地群落地上生物量对比

图2 处理组间草地群落地下生物量对比

图3 处理组叶片化学计量特征
2.2.2 根系化学计量特征对N,P添加的响应 由图4看出,不同地形条件下CK处理、N处理组和N+P处理组根系N含量的取值范围分别为5.20~12.78 g/kg,5.42~14.30 g/kg,10.16~17.70 g/kg,平均值分别为8.94 g/kg,12.00 g/kg,14.61 g/kg,变异系数分别为26.76%,25.78%和15.50%,与CK处理相比较,N处理和N+P处理分别增加34.38%和63.45%,但是除阳坡坡下部外其他地形在处理间并没有形成显著性差异(p>0.05)。CK处理、N处理组和N+P处理组根系P含量的取值范围分别为0.22~0.38 g/kg,0.38~0.87 g/kg,0.63~0.94 g/kg,方差分析表明施肥处理组根系P含量显著增加(p<0.05),N处理和N+P处理分别为CK处理的2.3倍和2.8倍。
三处理组根系K含量取值范围为1.14~6.47 g/kg,变异系数为11.61%,平均值分别为5.30 g/kg,4.96 g/kg,5.22 g/kg,施肥处理与CK处理没有显著差异。CK处理、N处理和N+P处理根系N/P比的取值范围分别为26.02~49.94,14.23~20.67,12.46~21.28,平均值分别为37.38,17.67,16.95,变异系数分别为24.13%,12.13%和15.82%,施肥处理组根系N/P呈下降趋势,其中阳坡施肥与否差异不显著(p>0.05),而坡顶和阴坡差异达到显著水平(p<0.05)。

图4 处理组根系化学计量特征
2.3 草地生态系统土壤养分对N,P添加的响应
由表2看出,施肥与否对土壤表层有机C含量影响不大,CK处理、N处理和N+P处理三组的土壤有机C含量平均值范围分别为5.48~12.41 g/kg,7.15~16.98 g/kg和7.38~13.13 g/kg,其中阳坡坡中部和坡上部N处理组和N+P处理组土壤有机C含量减小,其他地形则稍有增加,但各地形条件下均无显著差异(p>0.05)。施肥处理土壤N含量显著增加(p<0.05),CK处理、N处理和N+P处理土壤全N含量平均值分别为0.63 g/kg,1.59 g/kg,1.58 g/kg,N处理和N+P处理分别比CK处理增加154%和151%,其中阳坡N处理土壤全N含量大于N+P处理组,阴坡则呈现相反趋势,但两组间无显著差异(p>0.05)。不同地形条件下N处理和N+P处理土壤P含量均增加,但增加不显著(p>0.05),CK处理、N处理和N+P处理土壤全P含量平均值分别为0.46 g/kg,0.52 g/kg,0.54 g/kg。施肥处理土壤速效K含量增加,其中阳坡坡下部、坡顶和阴坡坡上部达到显著水平(p<0.05),其他地形虽有增加但没有形成显著差异(p>0.05)。土壤N/P比值变化见图5,施肥处理土壤N/P比值显著增大(p<0.05),CK处理、N处理和N+P处理组的平均值分别为1.34,3.03,2.86。

表2 处理组土壤养分含量

图5 处理组间土壤N/P变化
3 结 论
3.1 生物量对N,P添加的响应
目前N,P添加已成为保证土壤营养元素供应和保持植物生产力的有效措施[12]。大量研究表明N,P添加可有效增加植物地上生物量[13-15]。本研究中,N处理组和N+P处理组均使群落地上生物量增加,且N+P处理组的增加幅度更大,但增加趋势没有形成显著差异,可能是因为黄土高原丘陵区植物生长除了受养分条件限制外还受其他环境因子,尤其是降雨量的限制。但本试验研究过程中未考虑土壤水分的影响作用,有待进一步的研究验证。与地上生物量响应施肥处理不同,植物根系生物量及特征对施肥的响应在各研究中争议较大,尚未形成统一定论。Majdi等[16]研究表明添加N肥云杉根系生物量增加。杨晓霞等[13]基于高寒草甸的施肥试验,研究表明施加N肥对群落地下生物量无显著影响,而施加P肥后地下生物量则呈现增加趋势。辛小娟等[14]对亚高山草甸施肥后植物养分分配的研究结果表明N,P混施条件下,地下生物量变化不明显。祁瑜等[15]研究发现随施N量的增加,植物地下部分特别是细根生物量的分配比例显著降低。詹书侠等[17]研究不同施肥梯度羊草的响应情况,结果表明羊草地下生物量主要受P限制,低N和中N水平条件下,施P肥显著降低羊草的地下生物量。本研究中施肥处理地下生物量减小,与上述研究结果存在差异可能是因为群落物种组成不同,响应环境变化的机制及敏感度不同,且土壤养分存在时空变异性。总体而言,添加N,P肥后,限制植物生长的因子由地下养分转为光照[13,18-19]等其他因素的竞争,同时结合生物的最优分配假说,由于土壤养分状况相对良好,植物光合作用产生的养分更多的用于供应地上部分生长,以便获得更多光照,增大其竞争优势。Müller等[20]也认为在养分条件较好时,植物倾向于将生物量较多地分配于地上部分,养分条件较差时倾向于分配至地下部分。
3.2 群落生态化学计量特征对N,P添加的响应
3.2.1 叶片群落生态化学计量特征对N,P添加的响应 近年来随全球气候变化,大气N沉降增加[8],因而探究陆地生态系统中N含量增加对群落生态化学计量学特征的影响具有重要意义[21]。施肥作为一种高效的增加土壤养分、提高土地生产力的方法,可以有效模拟N含量增加的自然状况。因此研究N,P添加对群落生态化学计量学特征的影响,对认识生态系统生物地球化学循环对N沉降的响应具有重要意义[22]。本试验研究N,P添加对陕北黄土丘陵区不同坡向坡位条件下群落生态化学计量学的影响,结果显示,施肥处理组叶片N含量增加,这与刘洋[5]和常云妮[23]等人的研究结果一致,在营养元素缺乏条件下外源添加促进植物群落养分吸收,因此本试验外源N,P添加增大了群落水平上的叶片N含量,且N,P肥的交互作用使得N+P处理叶片N含量大于单施N肥。叶片P含量在施肥处理与对照间没有显著变化,这与前人研究结果施加N肥使得巨桉幼苗P含量减小不同[5],可能是因为物种不同,生长习性不同,对环境变化的适应性也会不同。本试验叶片P含量平均值(1.69 g/kg)高于Han等[24]测定的全国植物平均水平(1.21 g/kg)和张海东等[25]测定的黄土高原丘陵区草地群落叶片P含量(1.49 g/kg),表明施肥促进叶片P吸收。施肥处理叶片K含量呈现增加趋势,但并没有形成显著差异,可能是因为植物有保持元素组成相对稳定的自我调节机制,且黄土高原区叶片N,P,K含量呈现显著正相关[8,21],叶片N,P含量增加促进植物对K元素的吸收。黄土高原区植物体内N含量高于全国及全球平均水平,而P含量低于全球平均水平[8],因此N/P比值偏高。本试验中N处理组N/P增大,N+P处理组N/P减小,原因为单施N肥,植物叶片N含量增幅大于叶片P含量增幅,而施加N+P肥不仅促进N的吸收,同时增大了叶片含P量,相对减缓黄土高原区植物受P元素限制。
3.2.2 根系群落生态化学计量特征对N,P添加的响应 相对于植物叶片而言,研究根系养分状况的文章较少,且通过施肥来研究根系养分变化情况的更少。而根系作为植物体吸收水分和营养元素、支撑植物生长的最主要器官,在植物生长发育过程中起重要作用。本文研究结果显示,施肥处理根系N,P,K含量及N/P比值都发生了改变。施肥处理根系N增加,这与刘洋等[5]的研究结果一致,施加N肥和N+P肥均使得根系N含量显著增大,且与叶片N含量变化趋势相同,其中N+P处理根系N含量增加更明显。本试验中N处理和N+P处理根系P含量均增加,这与前人研究结果不同,可能是受限营养元素不同,且试验材料不同,本试验基于群落水平,因此响应也存在差异。与叶片K含量略有增加趋势不同,根系K含量在施肥处理后并没有显著改变,这可能是因为植物个体不同器官对环境变化的响应机制不同,体现了植物体养分分配规律的变化。根系N/P比值显著减小,表明施肥处理减缓了植物根系受土壤N含量限制的状况。
3.3 土壤养分对N,P添加的响应
植物生长状况受土壤养分状况影响,施肥则直接改变土壤C,N,P含量,能够快速有效改善土壤养分含量[26]。目前,N,P添加对土壤有机C的影响尚无定论,黄菊莹等[3]研究表明添加不同浓度N肥后土壤有机C含量没有明显的变化规律。本试验N,P添加后土壤有机C含量有所增加但无显著性差异,可能是因为施肥后群落生物量增加,增大了土壤有机质的来源,同时增大了土壤中微生物的活性,加快了土壤有机质分解。魏金明等[27]对内蒙古典型草原土壤状况的研究结果表明施加N肥对土壤全N含量没有显著影响;而德科加等[28]设置不同施肥浓度对高寒草甸植物及土壤养分进行探究,结果表明随施N浓度增加土壤全N含量先增加后趋于稳定;本试验结果显示土壤全N含量施肥后显著增加,可能与施肥量有关,除满足植物吸收供应外,土壤中仍有剩余,而本试验未设置不同施肥浓度,有待进一步研究验证;同时土壤养分受样地群落、水分及气候条件的影响,地形条件的限制。德科加的研究中施肥处理土壤全P含量变化不明显;卡着才让[29]等研究表明施加不同浓度N肥后,土壤全P含量降低,但不显著;本研究施肥处理土壤全P含量增加,可能是土壤质地、施肥方式及施肥量影响土壤养分对施肥的响应。施肥处理土壤速效K含量增加,这与前人研究结果一致[27],可能是由于施N肥和P肥后2∶1型黏土矿物中固定的钾离子被铵根离子置换。土壤N/P比值增大,主要因为土壤N含量增幅较P含量大。
黄土高原丘陵区不同地形条件下施肥处理土壤养分及植物叶片、根系N,P,K含量变化是直接改变群落生物量的最主要因素,尤其是N元素,相关研究表明N对作物生产力的贡献率高达40%~50%[30]。且不同试验研究中,受群落组成、施肥方法和试验地域及尺度的影响,群落及土壤对施肥的响应存在较大差异,本试验未设置不同的施肥浓度,且施肥年限较短,需要进行后续相关研究,以探索植物及土壤对施肥的响应机制。
参考文献:
[1] 牟雪.氮添加对草地碳氮磷化学计量特征的影响[J].中国水运(下半月),2015(3):100-101.
[2] 高扬,徐亚娟,彭炎,等.紫色土坡耕地C、P与微生物生物量C、P对不同施肥的响应[J].应用生态学报,2015,(1):108-112.
[3] 黄菊莹,赖荣生,余海龙,等.N添加对宁夏荒漠草原植物和土壤C∶N∶P生态化学计量特征的影响[J].生态学杂志,2013(11):2850-2856.
[4] 代景忠,卫智军,闫瑞瑞,等.N、P添加对羊草割草场土壤养分及地上生物量的影响[J].中国草地学报,2016,38(2):52-58.
[5] 刘洋,张健,陈亚梅,等.氮磷添加对巨桉幼苗生物量分配和C∶N∶P化学计量特征的影响[J].植物生态学报,2013,(10):933-941.
[6] 宋丰骥,常庆瑞,钟德燕.黄土高原沟壑区土壤养分空间变异及其与地形因子的相关性[J].西北农林科技大学学报:自然科学版,2011,39(12):166-172.
[7] 郑淑霞,上官周平.黄土高原地区植物叶片养分组成的空间分布格局[J].自然科学进展,2006,16(8):965-973.
[8] 刘世梁,郭旭东,连纲,等.黄土高原土壤养分空间变异的多尺度分析:以横山县为例[J].水土保持学报,2005,19(5):105-108.
[9] 薛超玉,焦峰,张海东,等.黄土丘陵区弃耕地恢复过程中土壤与植物恢复特征[J].草业科学,2016,33(3):368-376.
[10] 汝海丽,张海东,焦峰,等.黄土丘陵区微地形梯度下草地群落植物与土壤碳、氮、磷化学计量学特征[J].自然资源学报,2016,31(10):1752-1763.
[11] 鲍士旦.土壤农化分析 [M].3版.北京:中国农业出版社,2000.
[12] 孙小妹,肖美玲,师瑞玲,等.营养元素添加对青藏高原亚高寒草甸物种多度分布格局的影响[J].兰州大学学报:自然科学版,2014,50(6):853-859.
[13] 杨晓霞,任飞,周华坤,等.青藏高原高寒草甸植物群落生物量对氮、磷添加的响应[J].植物生态学报,2014,38(2):159-166.
[14] 辛小娟,王刚,杨莹博,等.氮、磷添加对亚高山草甸地上/地下生物量分配的影响[J].生态科学,2014,33(3):452-458.
[15] 祁瑜,黄永梅,王艳,等.施氮对几种草地植物生物量及其分配的影响[J].生态学报,2011,(18):5121-5129.
[16] Majdi H, Andersson P. Fine root production and turnover in a Norway spruce stand in northern Sweden: Effects of nitrogen and water manipulation[J]. Ecosystems,2005,8(2):191-199.
[17] 詹书侠,郑淑霞,王扬,等.羊草的地上—地下功能性状对氮磷施肥梯度的响应及关联[J].植物生态学报,2016,(1):36-47.
[18] Lan Z, Bai Y. Testing mechanisms of N-enrichment-induced species loss in a semiarid Inner Mongolia grassland: Critical thresholds and implications for long-term ecosystem responses[J]. Philosophical Transactions of the Royal Society of London B: Biological Sciences, 2012,367(1606):3125-3134.
[19] 潘庆民,韩兴国,杨景成.氮素对内蒙古典型草原羊草种群的影响[J].植物生态学报,2005,29(2):311-317.
[20] Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants[J]. Perspectives in Plant Ecology, Evolution and Systematics, 2000,3(2):115-127.
[21] Güsewell S. N:P ratios in terrestrial plants: Variation and functional significance[J]. New Phytologist, 2004,164(2):243-266.
[22] 黄菊莹,赖荣生,余海龙,等.N添加对宁夏荒漠草原植物和土壤C∶N∶P生态化学计量特征的影响[J].生态学杂志,2013,32(11):2850-2856.
[23] 常云妮,钟全林,程栋梁,等.氮添加对华润楠苗木碳氮磷化学计量特征的影响[J].甘肃农业大学学报,2013,48(5):94-99.
[24] Han W, Fang J, Guo D, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005,168(2):377-385.
[25] 张海东,汝海丽,焦峰,等.黄土丘陵区退耕时间序列梯度上草本植被群落与土壤C、N、P、K化学计量学特征[J].环境科学,2016,37(3):1128-1137.
[26] 刘昊,赵宁,曹喆,等.干扰对草地植被与土壤的影响之研究进展[J].中国农学通报,2008,24(5):8-16.
[27] 魏金明,姜勇,符明明,等.水、肥添加对内蒙古典型草原土壤碳、氮、磷及pH的影响[J].生态学杂志,2011,30(8):1642-1646.
[28] 德科加,张德罜,王伟,等.施肥对高寒草甸植物及土壤N, P, K的影响[J].草地学报,2014,22(2):299-305.
[29] 卡着才让,德科加,徐成体.不同施肥时间及施氮水平对高寒草甸生物量和土壤养分的影响[J].草地学报,2015,23(4):726-732.
[30] 陆景陵,胡霭堂.植物营养学[M].北京:高等教育出版社,2006.
