离子注入对杉木幼苗光合生理的影响
2018-05-04张斌
张 斌
(中南林业科技大学 林学院,湖南 长沙 410004)
杉木(Cumninghamialanceolata)为杉科(Taxodiaceae)杉属高大乔木,在我国具有悠久的栽培和利用历史,也是我国重要的造林树种,其面积和蓄积量均居我国速丰林前列[1],我国的杉木丰产栽培和良种选育的研究从 “五·五”期间开始,杉木的栽培面积持续稳定的增加,杉木的良种化水平和单位面积蓄积量不断提高,截至到“十二·五”期间,杉木的木材产量占我国人工林总产量的1/4左右[2],杉木在我国人工林中的地位日益突出。但杉木人工林的发展也出现了产量增长乏力、多代连作导致的地力衰退以及良种化水平低等多方面的问题[3],目前国内主要开展了家系选择和优良无性系繁育[4]、杉木营养[5-6]、种子园营建[7]、杂交育种[8]、混交林营造[9]、密度调控等方面的研究,其目的是为了缩短育种周期并提高杉木林分产量和干形指标,杉木速丰林产业的发展在于多途径的实现快速育种目标[10-11],从而缩短杉木遗传改良和良种选育周期。
离子注入的生物效应最早发现于20世纪80年代,近年来随着离子注入技术的提高,在生物材料领域特别是农业和林业领域取得了大量的成果[12],研究发现离子注入种子、芽等材料后,不仅能提高作物的产量、品质,而且结果具有高突变率和广突变谱的特点,是比太空辐射和放射性同位素辐射更为安全经济的诱变新途径。由于氮元素也是生物材料的重要组成元素,且氮离子容易获得,因此氮离子注入是多年来主要采用的离子源;同时研究发现,同氮离子相比,钛离子的诱变效果更佳[13-14]。研究表明离子注入对山核桃(Caryacathayensis)[15]、伞树(Acaciasaligna)[16]、桉树(EucalyptusL. Herit)[17]等树种的光合速率、同工酶活性、荧光特性和叶绿素含量均有明显的促进作用,对水稻(Oryzasativa)[18]、辣椒(Capsicumannuum)[19]等农作物的光合作用也要显著的促进作用。正是基于离子注入技术的这些特点,本研究采用氮离子和钛离子作为诱变原,基于离子注入诱变育种理论,以未进行离子注入处理的实生苗为对比,探索不同离子注入对杉木实生苗光合性能的影响,探讨其光合生理的日变化和季节变化规律,为开展更广泛的杉木离子注入育种工作提供基础依据。
1 材料和方法
1.1 材料与试验设计
离子注入设备选择成都同创公司生产的LZD-1000型多功能离子注入机,离子源选择N+和T2+两种,能量为30 keV/10mA,脉冲间隔时间5 s,剂量5×1016ions/cm2[20-22],离子注入时的真空度控制在10-3~10-4Pa。
将种子园2代种子进行离子注入后常规播种育苗,当苗龄为2年时,选择苗高为60~65 cm、地径为4.0~4.3 mm、无机械损伤的健壮苗木进行光合生理指标连续测定。以真空时间相同但未经离子注入的杉木种子采用相同的办法播种育苗后为对照。为减少种子自身变异对实验结果带来的影响,每个处理选择生长健壮且形态指标基本一致的30株幼苗进行测定,测试前选定5片完整的当年生叶片进行标记,以后每次测量相同的叶片,采用LI-6400光合作用测定系统对每株苗木重复测定4次,当数据稳定时每隔10 s记录1次数据。
1.2 指标测定
光合作用日变化每月测定1次,选择晴朗的天气进行,08:00—17:00,每隔1 h测定1次净光合速率(Pn)、蒸腾速率(Tr)和光合有效辐射(PAR),并根据蒸腾速率和光合有效辐射计算水分利用效率和光能利用效率,其中水分利用效率WUE=净光合速率Pn/蒸腾速率Tr,光能利用效率SUE=净光合速率Pn/光合有效辐射PAR。光响应曲线测定时首先设定在0~2 000 μmol/(m2·s)光通量范围内,共设14个光照强度梯度,对不同离子注入处理后的杉木幼苗叶片进行光响应曲线测定,测定时的环境温度为(25±2) ℃,湿度(75±3)%。
1.3 数据统计与处理
数据处理和制图采用SPSS18.0完成,数据均采用平均值,08:00—17:00每小时数据的平均值,数据每月采集1次,并进行LSD多重比较。不同处理间总体的差异性分析采用巢式分析法。
2 结果与分析
2.1 光合生理因子的日变化规律
2.1.1 净光合速率的日变化 表1中不同处理在各时间段Pn均值和变化趋势表明,离子注入处理能影响到杉木的光合速率日变化:离子注入处理能提高杉木苗木的光合能力,减弱光合午休现象,特别是Ti2+处理注入对光合生理的影响更加积极。
光合日变化呈双峰型变化趋势,且均在09:00达到最高值,Ti2+处理的光合速率最高,峰值达到17.85 μmol/(m2·s),比N+和对照处理分别增加7.08%、17.05%(α=0.021),N+处理比对照增加9.3%,14:00出现第2个峰值,但较上午偏小,此时仍然是Pn(Ti2+)(14.57 μmol/(m2·s))>Pn(N+)(13.01 μmol/(m2·s))>Pn(CK)(12.11 μmol/(m2·s)),Ti2+处理和N+处理的光合速率分别比对照处理高出20.31%(α=0.013)和7.43%,离子注入处理之间差异不显著。在光合午休条件下,净光合速率也呈现Pn(Ti2+)>Pn(N+)>Pn(CK)的规律,此时的最低值比09:00最高值分别下降了35.80%(α=0.000)、37.19%(α=0.000)、60%(α=0.000),Ti2+处理的下降幅度最小。从其他时间段不同离子注入处理间的对比分析来看,Ti2+离子注入处理在09:00—14:00,净光合速率均显著高于对照处理(α=0.000~0.033),N+离子注入处理在11:00—13:00以及15:00的净光合速率显著高于对照处理(α=0.000~0.039),Ti2+、N+离子注入相比只有在12:00(α=0.010)、15:00(α=0.000)差异显著。
从全天平均光合速率来看,Ti2+(11.27 μmol/(m2·s))>N+(10.73 μmol/(m2·s))>CK(9.17 μmol/(m2·s)),但3个处理差异不显著。
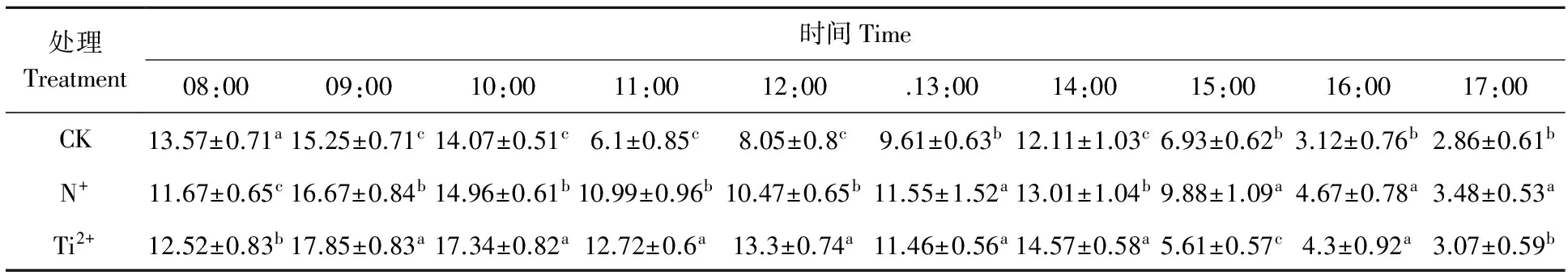
表1 不同处理净光合速率描述性统计结果
相同字母表示之间差异不显著,小写字母表示在0.05水平上差异显著。
Same letter means the difference between treatments is not significant,lower case means significant difference at the 0.05 level.
2.1.2 水分利用效率日变化 水分利用效率(WUE)的高低反应了杉木在不同处理条件下对体内水分利用的程度,杉木的水分利用效率是以杉木的净光合速率与蒸腾速率的比值来表示,反应了杉木的能量转化效率的高低,因此可以用水分利用效率的高低来揭示不同离子注入处理后杉木叶片气孔控制光合作用和蒸腾作用的过程。从表2不同处理在各时间段WUE均值和变化趋势来看,不同处理后的水分利用效率日变化呈现相同的L型曲线,上午光照强度较低的情况下水分利用效率最高,随着光照强度的增强,水分利用效率快速下降,08:00水分利用效率极显著的高于其他时段(α=0.000),随着光照强度的增加,对照处理在09:00、11:00以及16:00—17:00的水分利用效率高于时段(0.022~0.049),N+注入处理在09:00和17:00高于其它时段(α=0.017~0.040),Ti2+注入处理在09:00显著高于其他时段(α=0.000~0.021),其他时间段内不同处理间的水分利用效率差异不显著。从08:00与其他时段水分利用效率均值相比较来看,当09:00光照辐射大于800 μmol/(m2·s)后,水分利用效率变化幅度减弱,WUECK由08:00的15.66下降到其他时段的平均值3.96,WUEN+由17.21下降到3.47,WUETi2+由17.32下降到3.45,下降均达到极显著水平(α=0.000)。从WUE的日平均值来分析,对照处理的水分利用效率日均值最高(5.13),N+注入处理后的日均WUE下降到4.85,为对照处理的94.54%;Ti2+注入处理后的日均WUE下降到4.84,为对照处理的94.35%,由此可见,离子注入处理对杉木的水分利用效率影响不大。

表2 不同处理水分利用效率描述性统计结果
2.1.3 光能利用效率日变化 光能利用效率(SUE)反应的是杉木对于不同强度的光合有效辐射的利用强度,可直接体现杉木在离子注入处理后在不同光照强度下的光合生理指标的大小。从表3中不同处理在各时间段SUE均值和变化趋势来看,随着光照辐射强度的增加,杉木的光能利用效率都存在不同程度的下降,但仍保持SUETi2+>SUEN+>SUECK的趋势,离子注入处理对光能利用效率有一定的促进作用,提高了杉木幼苗潜在的光能利用能力。
从光能利用效率的最大值来看,SUETi2+的最大值是SUEN+的1.49倍、SUECK的2.78倍,均达到显著差异水平(α=0.000),其他时段内不同处理间的水分利用效率也达到显著差异水平(α=0.000)。从不同时段的光能利用效率来看,在11:00—16:00,相同的处理下光能利用效率差异不明显。从全天的SUE平均水平来看,SUETi2+(0.043)>SUEN+(0.029)>SUECK(0.013),也达到显著差异水平(α=0.000)。

表3 不同处理光能利用效率描述性统计结果
2.1.4 净光合速率的与主要影响因子的关系 根据各环境因子与光合速率的相关系数可以看出(表4),同对照处理相比,经氮离子注入处理后,杉木的Pn与PAR、Cond、Ci、MC呈极显著正相关关系,与RH呈显著正相关关系,与Tr、Vpdl的相关性不显著,其中Pn与PAR、MC的相关性增加(R2=0.924、0.907),与Cond、Ci、RH的相关性降低(R2=0.936、0.894、0.674)。
经钛离子注入处理后,杉木的Pn与PAR、Cond、Ci、MC呈极显著正相关关系,与RH呈显著正相关关系,其中除与Cond的相关性增加(R2=0.948),与PAR、Ci、MC、RH的相关性降低(R2=0.875、0.858、0.894、0.670),与Tr、Vpdl的相关性不显著。

表4 净光合速率与环境因子的相关分析
*表示在0.05水平上差异显著,**表示在0.01水平上差异显著
* mean significant difference at the 0.05 level,** mean significant difference at the 0.01 level
2.2 光合生理因子的年变化规律
2.2.1 净光合速率的年变化 图1表明,离子注入处理对杉木净光合速率季节变化有一定的影响。离子注入处理能提高杉木苗木的光合能力,其中钛离子注入对杉木净光合速率的影响略高于氮离子注入。
经氮离子和钛离子注入处理后,杉木苗叶片的净光合速率在7月份之前差别不大,在生长旺盛的7月至10月间,离子注入处理能提高杉木幼苗的光合作用强度。与对照处理相比,经氮离子处理后,净光合速率分别增加了9.87%~23.89%,差异显著(α=0.020~0.049),经钛离子处理后,净光合速率增加了20.35%~28.84%,差异显著(α=0.040 ~ 0.018)。速生期(7—10月)的平均净光合速率比缓生期(4—6月)分别增加了70.92%(CK)、99.50%(N+)、104.48%(Ti2+),从全年平均净光合速率来分析,氮离子处理比对照处理增加了13.57%(α=0.048),钛离子处理比对照增加了18.31%(α=0.039),差异均达到显著水平,巢式方差分析结果也表明,不同处理后杉木幼苗的净光合速率存在显著差异(α=0.001)。
2.2.2 水分利用效率的年变化 从图2可以看出,水分利用效率以未注入离子的处理下最高,氮离子注入处理次之,钛离子注入处理下的水分利用效率最低。水分利用效率在7月份均达到最大值,其他月份不同处理间WUE值变化不大,此时的水分利用效率分别为其他月份均值的5.73倍(CK)、5.19倍(N+)和5.31倍(Ti2+)。从全年均值来分析,WUEN+、WUETi2+分别是WUECK的93.23%、86.2%,通过对水分利用效率的总体差异性进行巢式方差分析,不同处理间在水分利用效率上表现出显著性的差异(α=0.000),说明经离子注入处理后,杉木的水分利用效率下降,离子注入对杉木幼苗形成胁迫,导致蒸腾速率升高,水分利用效率下降。

图1 氮、钛离子注入对净光合速率年变化的影响Fig.1 Yearly curves of Pn under N+and Ti2+ implantation

图2 氮、钛离子注入对水分利用效率年变化的影响Fig.2 Yearly courses of WUE under N+and Ti2+ implantation
2.2.3 光能利用效率的年变化 从图3来看,离子注入处理后,各月份间不同处理杉木的光能利用效率变化不大,4—6月份为杉木生长初始期,光能利用率较低;7—8月份进入杉木生长速生期,但同4—6月份相比,光能利用效率没有明显的变化,但到了9月份,光能利用效率显著提高(α=0.000),从10月份开始,杉木开始进入生长后期,此时正值湖南高温干旱季节,光能利用效率分别比9月份下降了93.28%(CK)、87.70%(N+)、77.44%(Ti2+),但仍然表现出SUETi2+>SUEN+≈SUECK的趋势,经方差分析,10月份不同处理间的光能利用效率差异不显著,说明此时空气湿度和土壤水分含量的下降是影响杉木光能利用效率的重要因素。但光能利用效率的年均变化结果表明,不同处理方式对杉木幼苗的光能利用效率存在显著的影响(α=0.000)。
2.3 光响应曲线
离子注入处理能提高杉木对光合有效辐射的利用效率,利用经典的Farquhar模型来对不同离子注入后的光响应曲线进行拟合,可以发现钛离子注入效果好于氮离子注入,在不同的光照强度下,钛离子处理后幼苗的净光合速率显著高于未经离子注入处理的杉木幼苗(α=0.022)(图4)。同样从不同离子注入处理后的光补偿点、光饱和点、光响应指标可以看出,离子注入处理提高了杉木幼苗对光合辐射的利用效率,氮离子注入处理后光补偿点降低了62.5%(α=0.006),光补偿点升高了19.2%,但无统计意义上的差异;经钛离子注入处理后,光补偿点较对照处理下降了68.8%(α=0.006),光饱和点升高了43.7%(α=0.019)。表观量子效率的大小反应了植物吸收和转换光能的潜力,代表了其利用弱光的能力,从处理的结果来看,处理后的杉木都较对照有较强的耐阴性,从表观量子效率和呼吸速率的大小来看,氮离子注入处理后杉木的耐阴性较高。从光饱和点来比较钛离子注入后,杉木具有较高耐受强光照射能力和喜光特征。

图3 氮、钛离子注入对光能利用效率年变化的影响Fig.3 Yearly courses of SUE under N+and Ti2+ implantation
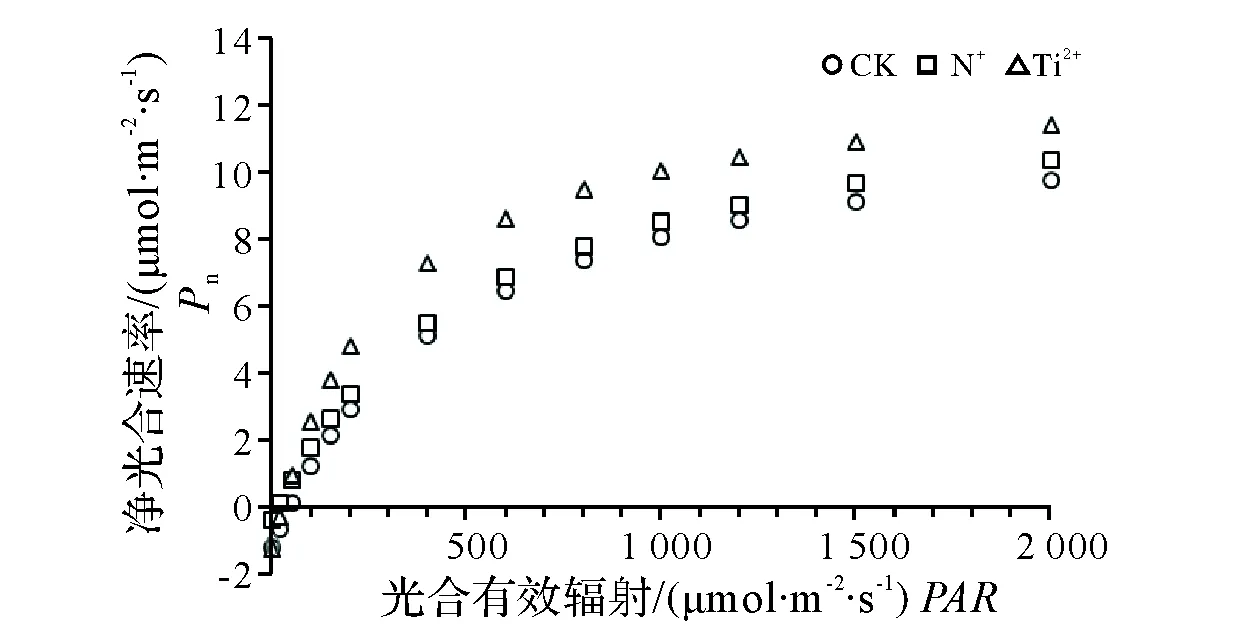
图4 不同离子注入处理后的光响应曲线Fig.4 Light responses of net photosynthetic rate for different ion beam implantation
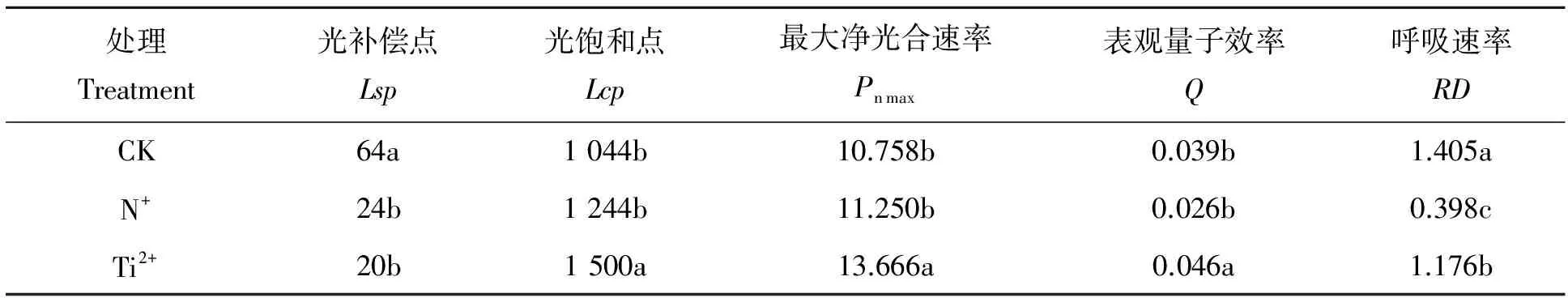
处理Treatment光补偿点Lsp光饱和点Lcp最大净光合速率Pnmax表观量子效率Q呼吸速率RDCK64a1044b10.758b0.039b1.405aN+24b1244b11.250b0.026b0.398cTi2+20b1500a13.666a0.046a1.176b
3 讨 论
(1)通过离子束辐射途径改变植物的生理活性是近几年离子生物辐射研究的热点,经不同能量和剂量的离子处理后,植物表现出较高的生理活性,在对农作物和木本植物的研究中发现低能离子束能增强净光合速率和水分利用效率等[20-22]。通过研究发现杉木的净光合速率不仅与光照强度、气温、CO2浓度等多种环境因子有关,也跟大多数植物一样,光合速率日变化存在双峰型的变化趋势[23-25]。上午的峰值高于下午,中午在光合辐射较高的情况下出现了光合午休现象,与未注入处理相比,氮离子和钛离子注入处理后光合午休现象减弱,与离子注入桉树[17]、光皮桦(Betulaluminlfer)[26]、闽楠(Phoebebournei)[27]等木本植物相似的结果。杉木光合速率年变化呈生长季节较高的单峰型变化趋势。在生长季节的7—10月,光合速率明显上升,其中以七月份的光合速率最高,但从不同处理后的净光合速率来看,离子注入处理下的净光合速率明显高于未处理的杉木苗木,除9月份氮离子处理稍好于钛离子注入处理,其他月份均属于钛离子注入的效果好于氮离子,根据张小全等[28]的研究结论,温度对针叶树种的净光合速率(Pn)有较大影响,由于9月份的气温高于其他月份,可能是由于经氮离子注入处理后提高了杉木幼苗耐高温能力,但离子注入处理后杉木光合能力与环境温度之间的关系还有待进一步研究。说明离子注入处理能提高杉木的光合利用效率和对强光辐射的难受能力。
(2)水分利用效率是以净光合速率与蒸腾速率的比值来表示,反应了杉木的能量转化效率的高低,因此可以用水分利用效率的高低来揭示不同离子注入处理后杉木叶片气孔控制光合作用和蒸腾作用的过程。水分利用效率既反应了杉木利用水分转化营养物质的能力,同时也反应了杉木在离子注入处理后在耐旱性上较对照处理表现略好,但差异不显著。水分利用效率的高低取决于净光合速率和蒸腾速率的大小,而蒸腾速率除了取决于杉木自身的生理特征外,还与空气湿度等外界环境有着密切的关系,根据净光合速率和蒸腾速率的相关性分析来看,离子注入处理后,由对照的显著相关减弱为不相关,虽然离子注入方式也能提高桉树的水分利用效率[17],但同水分和热胁迫带来的杉木永久水分利用效益更高的结果相比[29],离子注入处理后对杉木水分利用效率的影响能否像马尾松(Pinusmassoniana)、湿地松(Pinuselliottii)、桉树等树种具有持久的高水分利用效率还有待进一步的研究[30-33]。从杉木水分利用效率年变化规律来看,以生长旺盛的7月份最高,但都存在未经离子注入处理水分利用效率较高,钛离子注入处理的水分利用效率最低,但差异不显著,根据李荣生[34]的研究发现,植物的水分利用效率除了受环境等外界因素的影响外,还受到蒸腾速率、光合速率的影响,但同时也有研究表明[35],植物的水分利用效率受遗传物质的影响,试验中采用相同来源的杉木种子,在相同的环境条件下生长,不同离子注入处理后的水分利用效率表现大小不同的差异,这种差异的来源是否存在杉木遗传物质的改变还有待进一步的研究。
(3)光能利用效率能衡量植物的耐阴和喜光特性程度,也反应了植物利用光合辐射进行光合作用的能力和潜在的净生产力的大小[36-38],根据光响应曲线和光补偿点、光饱和点的变化趋势,可以得出,离子注入处理提高了杉木的光能利用效率,特别是钛离子的注入,杉木幼苗在耐阴性和适应强光辐射方面都有较佳的表现,效果好于氮离子的注入,也较杉木纯林的光能利用效率有明显的提高[37]。但从杉木幼苗的光能利用效率年变化来看,生长季节初期的光能利用率较低,此时的叶面积较小,叶片的光合功能发育不健全,同时由于气温较低,杉木的生长过程缓慢,温度是影响光能利用效率的主要因素;7月份开始,杉木进入生长旺盛的季节,但由于湖南进入干热少雨季节,杉木在高强度的光照辐射条件下易进入光合休眠状态,此时杉木的光能利用效率可能受土壤水分的影响较大,因此光能利用效率处于较低的水平;随着环境条件中湿度、温度等因素的充足,杉木的光能利用效率在9月份达到最大值,此时杉木的蒸腾能力增强,其光能利用效率也明显提高,但离子注入处理后的杉木的光能利用效率明显低于对照处理,不同离子处理之间差异不显著。根据仇宽彪[36]的研究发现,植物的光能利用效率受叶面积、湿度、蒸散比、叶龄等有关,离子注入处理后,杉木幼树的光能利用效率的变化较大,在水分条件较好的9月份,杉木处于生长季节的中后期,此时的净光能速率表现突出,研究结果与刘志高等[21]的研究结论类似,实验结果表明杉木的光能利用效率不仅与光合速率和光照辐射有关,在湖南地区,由于生长季节高温少雨,土壤中水分含量的多少、植物对水分的蒸散、杉木自身的生长规律等均不同程度的影响了杉木对光能利用效率的高低。
(4)离子注入处理能改变杉木种苗的生理活性指标,综合比较钛离子、氮离子和对照处理的杉木幼苗光合生理特性相关指标,从光能利用效率、水分利益效率和光响应曲线的变化趋势来看,钛离子注入的效果优于氮离子的注入,对杉木幼苗的生长具有一定的促进作用,离子注入后的生理特性变化规律在石蒜(Lycorisradiata)、油茶(Camelliaoleifera)植物上表现出类似结果[21-22]。在今后的木本植物诱变育种过程中采用钛离子诱导具有较高的效率。
离子注入处理带来的光合速率、水分利用效率、光能利用效率增加的同时,是否伴随着杉木抗逆能力的增加、生物量的变化、光合产物分配等变化情况[39,40],以及这种生理活性的提高能否具有遗传性[41,42]还要进一步的研究。
4 结 论
研究表明,杉木种子经过离子注入处理后,特别是经金属离子(Ti2+)注入处理后,其实生苗的净光合速率、水分利用效率、光能利用效率的日变化和年变化等生理活性明显提高,说明离子注入杉木种子对其实生苗的生长具有促进作用,其潜在的生长速率也会提高。与离子注入愈伤组织、芽等材料相比,离子注入处理杉木种子具有操作简便的优点,可以在杉木的离子注入诱变育种中使用,研究的结果可以为开展杉木等树种的选育提供新的途径,也能缩短育种的周期,提高林分的产量和质量。
参考文献:
[1] 张建国,盛炜彤,罗红艳,等.N、P、NP营养对杉木苗木生长和光合产物分配的影响[J].林业科学,2003,39(2):21-27.
Zhang J G,Sheng W T,Luo H Y,et al.Effects of N,P and NP nutrition on growth and allocation of photosynthetic products ofChineseFirseedlings[J].Scientia Silvae Sinicae,2003,39(2):21-27.
[2] 周学武,齐建文,刘金山,等.论储备林储备区域的选择—以五盖山国有林场杉木人工林为例[J].中南林业调查规划,2017,36(2):5-9.
Zhou X W,Qi J W,Liu J S,et al.Discussion on location choice of national reserve forest:a case study ofCunninghamialanceolateplantation in Wugai Shan national forest[J].Central South Forest Inventory and Planning,2017,36(2):5-9.
[3] 王伟峰,段玉玺,张立欣,等.不同轮伐期对杉木人工林碳固存的影响[J].植物生态学报,2016,40(7):669-678.
Wang W F,Duan Y X,Zhang L X,et al.Effects of different rotations on carbon sequestration inChinesefirplantations[J].Chinese Journal of Plant Ecology,2016,40(7):669-678.
[4] 郑勇平,孙鸿有,冯建民,等.杉木优良无性系龙15与闽33双系种子园遗传改良效应分析[J].浙江林学院学报,2009,26(2):201-208.
Zheng Y P,Sun H Y,Feng J M,et al.Genetic improvement on bi-clonal seed orchard ofChinesefirclone Long-15 and Min-33[J].Journal of Zhejiang Forestry College,2009,26(2):201-208.
[5] 韦丽丽,张小全,侯振宏,等.杉木苗木光合作用及其产物分配对水分胁迫的响应[J].植物生态学报,2005,29(3):394-402.
Wei L L,Zhang X Q,Hou Z H,et al.Effects of water stress on photosynthesis and carbon allocation inCunninghamiaLanceolataseedlings[J].Acta Phytoecologica Sinica,2005,29(3):394-402.
[6] 肖文发,徐德应,刘世荣,等.杉木人工林针叶光合与蒸腾作用的时空特征[J].林业科学,2002,38(5):38-46.
Xiao W F,Xu D Y,Liu S R,et al.The spatial and temporal characteristics of photosynthesis and transpiration of needles ofChinesefir[J].Scientia Silvae Sinicae,2002,38(5):38-46.
[7] 何贵平,徐肇友,王帮顺,等.杉木杂交试验苗期主要性状遗传分析[J].江西农业大学学报,2015,37(5):836-842.
He G P,Xu Z Y,Wang B S,et al.Genetic analysis of main traits ofChinesefirat the seedling stage in crossing test(NCⅡ)[J].Acta Agriculturae Universitatis Jiangxiensis(Natural Sciences Edition),2015,37(5):836-842.
[8] 颜志勤.三种化学诱变剂在杉木育种中的应用研究[D].福州:福建农林大学,2013.
Yan Z Q.Study on the application of three chemical mutagens in breeding ofChinesefir[D].Fuzhou:Fujian Agricultural and Forestry University,2013.
[9] 杨智杰,陈光水,谢锦升,等.杉木、木荷纯林及混交林凋落物量和碳归还量[J].应用生态学报,2010,21(9):2235-2240.
Yang Z J,Chen G S,Xie J S,et al.Litter fall production and carbon return inCunninghamialanceolata,Schimasuperbaand their mixed plantations[J].Chinese Journal of Applied Ecology,2010,21(9):2235-2240.
[10] 胡瑞阳,吴博,纳静,等.60Co-γ射线辐照处理对杉木种子萌发及幼苗生长的影响[J].中国农学通报,2016,32(4):1-4.
Hu R Y,Wu B,Na J,et al.Effects of60Co-γ radiation on seed germination and seedling growth ofCunninghamialanceolate[J].Chinese Agricultural Science Bulletin,2016,32(4):1-4.
[11] Shikha D,Jha U,Sinha S K,et al.Nitrogen ion beam modification of alumina for hard tissue implants[J].Macromolecular Symposia,2015,347(1):39-48.
[12] 焦浈,秦广雍,曹刚强,等.离子注入在生命科学中的应用研究[J].核农学报,2003,17(5):354-357.
Jiao Z,Qin G Y,Cao G Q,et al.Application of ion implantation in life science[J].Acta Agriculturae Nucleatae Sinica,2003,17(5):354-357.
[13] 龚洪恩,吕芳德,陈传松,等.离子注入对伞树幼苗叶绿素含量的影响[J].经济林研究,2014,32(1):140-143.
Gong H E,Lv F D,Chen C S,et al.Effects of ion implantation on chlorophyll content inAcaciasalignaseedlings[J].Nonwood Forest Research,2014,32(1):140-143.
[14] 郝杰,阮少宁,林思祖,等.Fe+与 Ti+注入对杉木苗高及相对电导率的影响[J].泉州师范学院学报,2014,32(2):13-16.
Hao J,Ruan S N,Lin S Z,et al.A comparative study of Fe+and Ti+ion implantation on seedling height and relative conductivity ofChinesefir[J].Journal of Quanzhou Normal University,2014,32(2):13-16.
[15] 龚洪恩.山核桃和金合欢属植物离子注入诱变育种的研究[D].长沙:中南林业科技大学,2005.
Gong H E.The study of ion implantation breeding on pecan and acacia species[D].Changsha:Central South University of Forestry and Technology,2005.
[16] 龚洪恩,吕芳德,陈传松,等.离子注入对伞树幼苗叶绿素含量的影响[J].经济林研究,2014,32(1):140-143.
Gong H E,Lv F D,Chen C S,et al.Effects of ion implantation on chlorophyll content inAcaciasalignaseedlings[J].Nonwood Forest Research,2014,32(1):140-143.
[17] 李光友,徐建民,杜志鹄,等.不同离子注入浓度对桉树家系光合特性的影响[J].华北农学报,2010,25(S2):156-159.
Li G Y,Xu J M,Du Z G,et al.A photosynthetic study on eucalyptus family with different N+dosage[J].Acta Agriculturae Boreali-Sinica,2010,25(S2):156-159.
[18] 李林玉,黄群策,张书艮,等.低能氮离子束与UV-B增强对水稻光合及蒸腾速率的影响[J].农业工程学报,2013,29(15):136-144.
Li L Y,Huang Q C,Zhang S Y,et al.Effects of low-energy N+ion beam irradiation on photosynthetic and transpiration in rice under enhanced UV-B radiation[J].Transactions of the Chinese Society of Agricultural Engineering,2013,29(15):136-144.
[19] 常晓蓓.低能氮离子注入对线辣椒当代的生物学效应[D].杨凌:西北农林科技大学,2015.
Chang X B.Biological effects of low energy N+implantation on chili pepper[D].Yangling:Northwest A&F Universtiy,2015.
[20] 张斌,李志辉,张冬林,等.N+注入对紫薇光合特性和叶绿素含量的影响[J].激光生物学报,2010,19(3):303-306.
Zhang B,Li Z H,Zhang D L,et al.Effects of N+beam implantation on photosynthesis characters and chlorophyll contents ofLagerstroemiaindicaLinn[J].Acta Laser Biology Sinica,,2010,19(3):303-306.
[21] 刘志高,张斌,童再康,等.钛离子注入对3种石蒜属植物种子生物化学特性的影响[J].核农学报,2013,27(2):146-151.
Liu Z G,Zhang B,Tong Z K,et al.Effects of Ti+implantation on biological and chemical characteristics of seeds from three species of Lycoris[J].Journal of Nuclear Agricultural Sciences,2013,27(2):146-151.
[22] 曾艳玲,谭晓风,张斌,等.钛离子注入对油茶种子生理生化反应及初期生长的影响[J].中国农学通报,2011,27(28):53-57.
Zeng Y L,Tan X F,Zhang B,et al.Effects of the Ti2+ion implantation on physiological-biochemical reaction and seminal growth ofCamelliaoleiferaseeds[J].Chinese Agricultural Science Bulletin,2011,27(28):53-57.
[23] Kocacinar F.Photosynthetic,hydraulic and biomass properties in closely related C3and C4species[J].Physiologia Plantarum,2015,153(3):454-466.
[24] 杜虎,曾馥平,王克林,等.中国南方3种主要人工林生物量和生产力的动态变化[J].生态学报,2014,34(10):2712-2724.
Du H,Zeng F P,Wang K L,et al.Dynamics of biomass and productivity of three major plantation types in southern China[J].Acta Ecologica Sinica,2014,34(10):2712-2724.
[25] 付威波,宋敏,彭晚霞,等.西南喀斯特典型树种光合特性的季节变化及主要影响因子[J].农业现代化研究,2015,36(5):890-894.
Fu W B,Song M,Peng W X,et al.Seasonal dynamics of photosynthesis and the underlying regulating factors for typical species in southwest karst[J].Research of Agricultural Modernization,2015,36(5):890-894.
[26] 郑丰河.光皮桦离子注入诱变育种研究[D].长沙:中南林业科技大学,2009.
Zheng F H.The study of ion implantation breeding onBetulaluminlfer[D].Changsha:Central South University of Forestry and Technology,2009.
[27] 胡婧楠.三种楠木光合生理特性及叶子养分年变化的比较研究[D].合肥:安徽农业大学,2010.
Hu J N.Comparative study on photosynthesis physiological characteristics and annual variety of leaf nutrient of threePhoebeplants[D].Hefei:Anhui Agricultural University,2010.
[28] 张小全,徐德应.温度对杉木中龄林针叶光合生理生态的影响[J].林业科学,2002,38(3):27-33.
Zhang X Q,Xu D Y.Effects of temperature on the photosynthetic physio-ecology of 18-year-oldChinesefir(Cunninghamialanceolata(lamb.)hook.)[J].Scientia Silvae Sinicae,2002,38(3):27-33.
[29] 胡化广,张振铭,吴生才,等.植物水分利用效率及其机理研究进展[J].节水灌溉,2013,38(3):11-15.
Hu H G,Zhang Z M,Wu S C,et al.Advance of research on water use efficiency of plant and its mechanism[J].Water Saving Irrigation,2013,38(3):11-15.
[30] 蒋建.南方主要造林树种养分利用效率研究[D].福州:福建农林大学,2006.
Jiang J.Research on the nutrient use efficiency of main planting species in south china[D].Fuzhou:Fujian Agricultural and Forest University,2006.
[31] 张秀娟.亚热带常绿林光能和水分利用效率研究—以江西千烟洲试验区为例[D].北京:北京林业大学,2011.
Zhang X J.Light and water-use efficiency of subtropical evergreen coniferous forest in qianyanzhou experimental station of Jiangxi provice[D].Beijing:Beijing Forestry University,2011.
[32] Zhou S X,Medlyn B E,Prentice I C.Long-term water stress leads to acclimation of drought sensitivity of photosynthetic capacity in xeric but not riparian Eucalyptus species[J].Annals of Botany,2016,117(1):113-144.
[33] Huang W W,Hu H L,Hu T X,et al.Impact of aqueous extracts ofCinnamomumseptentrionaleleaf litter on the growth and photosynthetic characteristics of Eucalyptus grandis seedlings[J].New Forests,2015,46(4):561-576.
[34] 李荣生,许煌灿,尹光天,等.植物水分利用效率的研究进展[J].林业科学研究,2003,16(3):366-371.
LI R S,Xu H C,Yin G T,et al.Advances in the water use efficiency of plant[J].Forest Research,2003,16(3):366-371.
[35] 腰政懋.辽东冷杉的光能和水分利用效率及其种源遗传变异规律研究[D].北京:北京林业大学,2015.
Yao Z M.Study on light and water use efficiency ofAbiesholophyllaand its provenance genetic variation[D].Beijing:Beijing Forestry University,2015.
[36] 仇宽彪,张志强,康满春,等.北京永定河沿河沙地杨树人工林光能利用效率[J].生态学报,2016,36(6):1-11.
Qiu K B,Zhang Z Q,Kang M C,et al.The dynamics of light use efficiency at a poplar plantation in Beijing[J].Acta Ecologica Sinica,2016,36(6):1-11.
[37] 张秀娟,韩海荣,王军邦.亚热带人工针叶林光能利用效率的差异[J].东北林业大学学报,2010,38(12):12-14.
Zhang X J,Han H R,Wang J B.Difference in light use efficiency of subtropical coniferous plantatio[J].Journal of Northeast forestry University,2010,38(12):12-14.
[38] 范晓明,袁德义,杨斐翔,等.锥栗不同树体结构对光的响应[J].生态学报,2015,35(22).7426-7434.
Fan X M,Yuan D Y,Yang F X,et al.Effects of different tree shapes on light use inCastaneahenryi[J].Acta Ecologica Sinica,2015,35(22).7426-7434.
[39] Guo J F,Chen G S,Xie J S,et al.Effect of heat-disturbance on microbial biomass carbon and microbial respiration inChinesefir(Cunninghamialanceolata) forest soils[J].Journal of Forestry Research,2015,26(4):933-939.
[40] Wen X D,Zhao Z H,Deng X W,et al.Applying an artificial neural network to simulate and predictChinesefir(Cunninghamialanceolata) plantation carbon flux in subtropical China[J].Ecological Modelling,2014,(294):19-26.
[41] 刘腾,郭九峰,那日,等.基于cDNA-AFLP的N~+离子注入甘草当代根部基因表达分析[J].核农学报,2015,29(6):1074-1080.
Liu T,Guo J F,Na R,et al.The analysis of differential rxpression ofGlycyrrhizauralensisrootgene by nitrogen ions implantationbased on cDNA-AFLP method[J].Journal of Nuclear Agricultural sciences,2015,29(6):1074-1080.
[42] 于立伟,张林,李静,等.空间诱变玉米自交系齐319的SSR标记变异分析[J].核农学报,2014,28(8):1345-1352.
Yu L W,Zhang L,Li J,et al.SSR analysis of mutants from maize ‘Qi319’inbredlines induced by space mutagenesis[J].Journal of Nuclear Agricultural sciences,2014,28(8):1345-1352.
