磷素对小麦籽粒支链淀粉合成及其链长分布的影响
2018-05-04张润琪付凯勇徐芳芳朱永琪覃安祥李春艳
张润琪,李 诚,付凯勇,李 超,徐芳芳,朱永琪,覃安祥,李春艳
(石河子大学农学院/新疆兵团绿洲生态农业重点实验室,新疆石河子 832000)
淀粉是小麦籽粒的主要组成成分,根据结构差异可分为直链淀粉和支链淀粉[1],其中支链淀粉占淀粉含量的70%~80%,其构型直接影响淀粉的理化特性和加工、食用品质[2]。支链淀粉的主链通过α-1,4-糖苷键连接而成,支链通过α-1,6-糖苷键与主链相连,可分为A、B和C三种链[3],其中,C链为主链,B链为C链的支链,而A链为B链的侧链。前人将支链淀粉的色谱图分为3 部分[4]:A链:DP 6~18,B1链:DP 19~34,B2链:DP>35。
淀粉的合成与结构不但与其遗传特性有关,还受栽培过程中营养水平的影响[5]。磷素是作物必需的三大营养元素之一,施加磷肥是缓解土壤低磷胁迫、改善作物产量和品质的有效方法。腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)是淀粉合成的关键酶之一,在植物叶片中受变构调节,被 3-磷酸甘油酸激活而被无机磷酸所抑制,3-磷酸甘油酸与无机磷酸的比值影响其活性,从而调节淀粉的合成[6]。可溶性淀粉合成酶(SSS)、淀粉分支酶(SBE)和淀粉去分支酶(DBE)是负责支链淀粉合成的关键酶[7],分别由其对应的酶基因编码[8]。其中,SBE的两种同工酶均可被磷酸盐激活[9]。磷素对淀粉的合成有重要影响。姜宗庆等[10]研究表明,在施磷(P2O5)量0 ~ 108 kg·hm-2范围内,小麦籽粒直链淀粉、支链淀粉、总淀粉积累量、积累速率均随施磷量的增加而上升。此外,磷对淀粉的品质特性也有显著影响,Li等[11]研究表明,施磷显著提高了小麦新冬20号籽粒淀粉中B型淀粉粒的比例,对其持水力、膨胀势、溶解性等也有显著影响。前人对于磷素影响小麦支链淀粉含量及支链淀粉合成关键酶活性已有研究,如Ni等[12]研究发现,施磷显著提高了成熟期小麦籽粒中的支链淀粉含量,王旭东[13]的研究表明,施磷提高了花后28 d之前籽粒中蔗糖合成酶(SS)、颗粒淀粉合成酶(GBSS)和可溶性淀粉合成酶(SSS)的活性,提高了花后14~28 d 籽粒中AGPase的活性,但关于磷素影响小麦支链淀粉合成关键酶基因表达及链长分布的研究则鲜见报道。本研究选用不同冬小麦品种,测定不同磷素水平下淀粉合成关键酶基因表达量以及对应酶的活性、籽粒支链淀粉含量、支链淀粉链长分布和籽粒全磷含量,探究不同磷素水平对小麦籽粒支链淀粉合成及其链长分布的调控效应,以期揭示小麦支链淀粉的积累特征,为预测小麦淀粉品质提供理论依据。
1 材料与方法
1.1 试验材料与设计
供试材料选用新冬20号(南疆主栽品种)和新冬23号(北疆强筋面包小麦),由石河子大学麦类作物研究所提供。
试验于2014年10月至2015年6月在石河子大学农学院试验站(44°17′N, 86°03′E)进行。前茬作物为向日葵,土质为灰漠土,0~20 cm 土层内含碱解氮63 mg·kg-1、速效磷15 mg·kg-1、速效钾208 mg·kg-1。播种时按75 kg·hm-2施加尿素,灌溉方式为滴灌,冬前浇水3次,返青至成熟每隔10~12 d浇水一次,共浇水6次,每次滴水量1 125 m3·hm-2,总滴水量10 125 m3·hm-2。在拔节期、抽穗期和扬花期分别随水施尿素45、75和120 kg·hm-2。
随机区组设计,3个重复,小区面积2.4 m×3 m,小区之间隔离带宽度为50 cm。设置3个磷素(P2O5)水平分别为0、105和210 kg·hm-2,分别用CK(对照)、CP(常规施磷)和HP(过量施磷)表示,所用肥料为重过磷酸钙,在播种后160 d(大约5%的植株已返青)开沟条施。
1.2 试验方法
1.2.1 样品采集
从每个小区选取同日开花的麦穗做好标记,分别在花后7、14、21、28和35 d在每个小区剥取穗中部的籽粒,同处理籽粒集中到一起,一部分经液氮速冻5 min后保存于-80 ℃冰箱,用于测定酶活性和提取RNA;另一部分于105 ℃杀青30 min,80 ℃烘干至恒重备用。
1.2.2 测定项目与方法
(1)籽粒支链淀粉含量的测定 取烘干的小麦籽粒用实验室粉碎磨(JFSD-70,上海嘉定)打成粉末,参照赵永亮[14]的方法测定。
(2)胚乳淀粉粒的提取 参照Peng等[15]的方法提取,常温干燥后于-20 ℃保存。
(3)支链淀粉链长分布的测定 参照贺 伟等[16]的方法。
(4)籽粒全磷含量的测定 取烘干的小麦籽粒用实验室粉碎磨(JFSD-70,上海嘉定)打成粉末,参照鲍士旦[17]的方法测定。
(5)支链淀粉合成相关酶活性的测定 胚乳SS酶活性的测定参照Douglas等[18]的方法;SBE酶活性的测定参照赵法茂等[19]的方法;DBE酶活性的测定参照刘 霞等[20]的方法。
1.2.3 支链淀粉合成相关酶基因相对表达量的测定
(1)引物设计 使用Primer Premier 5.0.软件,根据NCBI上公布的小麦籽粒淀粉合成相关酶基因的序列,设计 sbe1, sbe2a, sbe2b和 iso1基因的引物(表1),由生工生物工程(上海)股份有限公司合成。使用小麦actin基因(NCBI编号DN551593)作为内参。通过梯度PCR验证引物特异性同时优化PCR条件。
(2) RNA的提取和cDNA的合成 用Fruit-mate(Takara)和RNAiso plus(Takara)试剂盒提取RNA,按说明书操作,通过琼脂糖凝胶电泳检验提取的总RNA质量。使用TIANScript cDNA第一链合成试剂盒(Tiangen, Cat#KR104-02, China)合成cDNA,用其作为模板扩增actin基因以检验其质量。
(3)实时荧光定量PCR 使用SYBR Premix ExTaq试剂盒(Takara),在实时荧光定量PCR仪(Roche LightCycler 480 Ⅱ)上检测各个基因的扩增情况,具体操作按说明书进行。
1.3 数据处理
应用Excel 2003对数据进行统计分析和绘图,用SPSS 11.5进行单因素方差分析,采用邓肯法(Duncan)进行多重比较。
2 结果与分析
2.1 不同磷素处理对支链淀粉合成相关酶基因表达的影响
施磷显著提高了两个小麦品种花后14~28 d胚乳中 sbe1基因的相对表达量(图1),且CP处理显著高于HP处理(花后28 d除外)。新冬20号胚乳中 sbe1基因的相对表达量总体低于新冬23号。对照条件下,新冬20号胚乳中 sbe1基因相对表达量在花后28 d达到峰值,而新冬23号则在花后21 d达到峰值。CP处理下两个参试品种 sbe1基因的相对表达量基本呈先上升后下降的趋势,均在花后21 d达到峰值。HP处理下新冬20号胚乳中 sbe1基因的相对表达量呈双峰分布,而新冬23号则呈单峰分布。花后35 d,随着种子的成熟,两品种各处理下胚乳中 sbe1基因的相对表达量降至最低值。
施磷显著提高了两参试品种花后14~28d胚乳中 sbe2a基因的相对表达量(图2),且CP处理显著高于HP处理。新冬20号胚乳中 sbe2a基因的相对表达量整体低于新冬23号。对照条件下两品种胚乳中 sbe2a基因的相对表达量均较低;CP处理下新冬20号胚乳中 sbe2a基因的相对表达量在花后7 d最高,而新冬23号则在花后14 d达到峰值;HP处理下两参试品种胚乳中 sbe2a基因的相对表达量均在花后14 d达到峰值。

A:新冬20;B:新冬23。图柱上不同字母表示处理间差异在0.05水平显著。下同。
A:Xindong 20;B:Xindong 23.Different letters above columns mean significant difference among treatments at 0.05 level. The same below.
图1不同磷处理下小麦胚乳sbe1基因的相对表达量
Fig.1Relativeexpressionofsbe1inwheatendospermunderdifferentphosphoruslevels
如图3所示,施磷可不同程度提高两个小麦品种胚乳中 sbe2b基因的相对表达量。对照和CP处理下新冬20号胚乳中 sbe2b基因的相对表达量均在花后7 d最高,随后不断降低,而HP处理下则呈先上升后下降的趋势,在花后14 d达到峰值。各处理下新冬23号胚乳中 sbe2b基因的相对表达量均呈先升后降之势,都在花后14 d达到峰值。

图2 不同磷处理下小麦胚乳 sbe2a基因的相对表达量

图3 不同磷处理下小麦胚乳 sbe2b基因的相对表达量

图4 不同磷处理下小麦胚乳 iso1基因的相对表达量
如图4所示,施磷显著提高了两个参试品种花后7~28 d胚乳中 iso1基因的相对表达量(新冬20花后7、28 d除外),且CP处理下显著高于HP处理(花后28 d除外)。对照条件下两参试品种胚乳中 iso1基因的相对表达量均较低,且各时期变化不大。CP和HP处理下两品种胚乳中 iso1基因的相对表达量均呈先升后降之势,都在花后14 d达到表达峰值;花后35 d,随着灌浆的完成,两品种胚乳中 iso1基因的相对表达量下降至最低。
2.2 不同磷处理对支链淀粉合成相关酶活性的影响
磷处理对小麦胚乳中SS酶活性的影响存在品种间差异(图5),CP处理显著提高了新冬20号胚乳中的SS酶活性(花后14 d除外),促进了淀粉合成原料蔗糖向籽粒的供应;除花后14 d外,HP处理下新冬20号籽粒发育各时期胚乳SS酶活性均显著高于同期对照,花后35 d达到峰值;对照条件下胚乳中的SS酶活性呈升-降-升之势,在花后14 d达到最大值。CP处理下新冬23号的胚乳SS酶活性呈升-降-升的趋势,在花后35 d达到峰值且显著高于同期HP和对照处理;HP处理下新冬23号的胚乳SS酶活性整体呈先升后降之势,在花后21 d达到最大值且显著高于同期对照和CP处理;对照条件下的胚乳SS酶活性呈降-升-降的趋势,在花后28 d达到峰值。
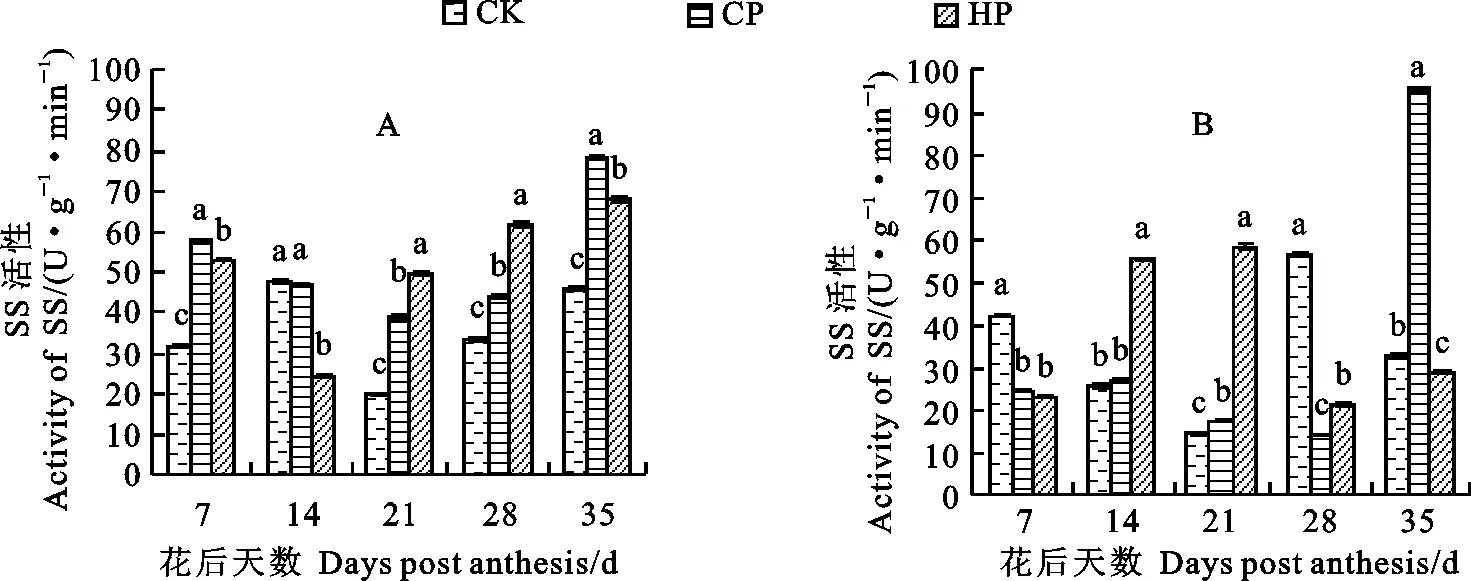
图5 不同磷处理下的小麦胚乳SS酶活性
如图6所示,新冬20号胚乳中的SBE酶活性CP处理下呈先升后降的趋势,在花后14 d达到最大值且显著高于同期HP和对照处理;在HP处理下呈升-降-升之势,分别在花后14 d和35 d有最高峰和次高峰,且显著高于同期对照处理;对照条件下胚乳SBE酶活性在籽粒灌浆期间逐渐下降。各处理下新冬23号胚乳中的SBE酶活性均在花后7 d酶活最高。

图6 不同磷处理下的小麦胚乳SBE活性
如图7所示,对照条件下新冬20号胚乳中的DBE酶活性呈降-升-降之势,花后28 d达到最大值;CP和HP处理下新冬20号胚乳中的DBE酶活性均在花后7 d最高,且CP处理下显著高于对照。相比于对照,施磷处理提前了新冬20号胚乳中DBE酶活性峰值的出现时间。对照和HP处理下新冬23号胚乳中的DBE酶活性均在花后7 d最高,之后则呈降-升-降的趋势;CP处理下胚乳中的DBE酶活性呈升-降-升之势,花后14 d达到最大值且显著高于同期对照和HP处理。
2.3 不同磷处理对籽粒支链淀粉含量的影响
如图8所示,两品种籽粒中的支链淀粉含量在灌浆早期都很低,随着籽粒的发育,其支链淀粉含量逐步提高。花后7 d时不同磷处理下的支链淀粉含量无显著差异,在此之后施磷显著或不显著提高了籽粒中支链淀粉的含量,成熟期(花后35 d)籽粒中CP处理下的支链淀粉含量最高,且显著高于对照和HP处理。
2.4 不同磷处理对籽粒支链淀粉链长分布的影响
由表2可以看出,CP处理提高了两品种支链淀粉中B链所占的比例。新冬20号各处理下支链淀粉中B链所占比例随籽粒的发育不断上升,而新冬23号对照的支链淀粉中B链所占比例呈先下降后上升的趋势,HP处理下呈先上升后下降的趋势,CP处理下则在花后各时期变化不大。

图7 不同磷处理下的小麦胚乳DBE活性

图8 不同磷处理下的小麦籽粒支链淀粉含量
2.5 不同磷处理对籽粒中全磷含量的影响
如图9所示,CP处理下,新冬20号花后7 d、21 d和28 d以及新冬23号花后21 d籽粒中全磷含量显著高于对照,其他时期及HP处理与对照相比并无显著差异,说明CP处理有提高籽粒全磷含量的趋势,但并不明显;花后35 d,随着种子的成熟,两参试品种各处理下籽粒中的全磷含量无显著差异。
3 讨 论
研究表明,低磷土壤(速效磷含量9.37~10.73 mg·kg-1)上随着施磷量的增加,小麦胚乳 sss3、 sbe1基因相对表达量升高,中磷(速效磷含量19.54~20.71 mg·kg-1)、高磷土壤(速效磷含量37.46~38.77 mg·kg-1)施磷对 sss3、 sbe1基因相对表达量调节不显著[21]。本研究表明,施磷可显著提高小麦胚乳 sbe1, sbe2a, sbe2b和 iso1基因的相对表达量,且CP处理下更加显著,这说明一定范围内施加磷肥有利于支链淀粉合成相关酶基因的表达。Wang等[22]认为,SSS、SBE和DBE可能在转录水平上调节淀粉的合成,与本研究结果基本一致。王树亮等[23]研究表明,随着土壤磷水平的提高,小麦各器官含磷量均有提高(其中籽粒磷含量提高幅度最小),但磷素利用效率降低。姜宗庆[24]的研究表明,成熟期籽粒磷含量随施磷量提高而增加,但施磷量超过一定水平则造成籽粒磷含量下降,与本研究的结果一致。由于AGPase与SBE酶活性均与磷素有关[8-9],因此,不同磷处理下籽粒磷含量的不同可能暗示了这两种酶活性的变化。

表2 不同磷水平下的小麦支链淀粉链长分布Table 2 Chain-length distribution of wheat amylopectin under different phosphorus levels %

图9 不同磷处理下的小麦籽粒全磷含量
支链淀粉的合成是多种酶协同作用的结果。姜宗庆等[10,24]研究发现,在缺磷土壤(速效磷含量4.1 mg·kg-1)上增加施磷量可提高小麦籽粒SS酶的活性、籽粒蔗糖含量和SBE酶的活性。本研究发现,施磷提高了新冬20号胚乳SS酶活性和SBE酶活性峰值,且提前了新冬20号胚乳中DBE酶活性峰值的出现时间。说明磷处理促进了淀粉合成原料蔗糖在籽粒中的积累,为淀粉合成提供了充足的原料,同时通过提高酶活高峰期的SBE酶活性和DBE酶活性峰值的出现时间,提高了支链淀粉的合成速率,促进了支链淀粉的积累。这与本研究结果中支链淀粉合成相关酶基因表达量、支链淀粉含量的提高相符。此外,本研究使用的试验材料新冬20号为南疆主栽弱冬性品种,早熟高产[25],全生育期较短(约238 d);而新冬23号则为北疆强冬性品种,中晚熟,中高产,全生育期较长(280~290 d)[26],品种特性差异可能是造成两品种支链淀粉合成关键酶活性对磷素敏感性不同的原因。石河子大学冬小麦课题组的水培试验亦表明,新冬20号根系比新冬23号更加发达(未发表),而根干重对植株磷素积累量有极显著的直接作用[27],此外,新冬20号生育期虽然较新冬23号短,但其产量却比新冬23号高,说明新冬20号对于磷素的吸收利用更加高效,因而其支链淀粉合成关键酶活性对磷素的敏感性相对较高。
本研究表明,适量施磷提高了支链淀粉中B链的比例,降低了A链的比例。这说明磷处理下支链淀粉的外侧链较短,对长链的空间影响作用力较小,长链的运动性较强,有利于相互靠拢形成双螺旋结构。前人研究表明,支链淀粉的链长分布对淀粉的品质特性有显著的影响。DP 6~11支链与相对结晶度呈极显著负相关,而DP 28~34支链与糊化温度呈极显著负相关[2];除崩解值外,长B链( DP≥65. 8)与黏度各特征值(峰值黏度、低谷黏度、最终黏度、反弹值、峰值时间)均呈极显著正相关[28]。此外,Li等[11]的研究表明,施磷显著提高了黏度特征值中的峰值黏度、低谷黏度、最终黏度和反弹值,显著降低了糊化温度,这与本研究中施磷提高B1链和B2链比例的结果相符。除长、短链外,内链、外链、平均链长、分支化度以及内外链比例对淀粉的结构也有影响,磷对淀粉品质特性的影响与这些因素的关系尚需深入研究。
本研究发现,磷素处理下两个小麦品种支链淀粉合成相关酶基因( sbe1、 sbe2a、 sbe2b、 iso1)的表达量显著上调,相关酶的活性也有所提高,从而增加了籽粒支链淀粉含量和支链淀粉B链段比例。不同磷素处理下支链淀粉含量与分子精细结构的变化可能还与其他淀粉合成酶基因表达量的变化、酶活性的改变等有关,其具体原因尚需进一步探究。
参考文献:
[1]WEI L,WANG L,YANG Y,etal.Abscisic acid increases leaf starch content of polyethylene glycol-treated wheat seedlings by temporally increasing transcripts of genes encoding starch synthesis enzymes [J].ActaPhysiologiaePlantarum,2015,37(10):2.
[2] 贺晓鹏,朱昌兰,刘玲珑,等.不同水稻品种支链淀粉结构的差异及其与淀粉理化特性的关系[J].作物学报,2010,36(2):277.
HE X P,ZHU C L,LIU L L,etal.Difference of amylopectin structure among various rice genotypes differing in grain qualities and its relation to starch physicochemical properties [J].ActaAgronomicaSinica,2010,36(2):277.
[3] 李海普,李 彬,欧阳明,等.直链淀粉和支链淀粉的表征[J].食品科学,2010,31(11):275.
LI H P,LI B,OUYANG M,etal.Advances in characterization of amylose and amylopectin starch [J].FoodScience,2010,31(11):275.
[4] NAGAMINE T,KOMAE K.Improvement of a method for chain-length distribution analysis of wheat amylopectin [J].JournalofChromatographyA,1996,732(2):257.
[5] 梁晓芳,于振文.施钾时期对冬小麦旗叶光合特性和籽粒淀粉积累的影响[J].应用生态学报,2004,15(8):1349.
LIANG X F,YU Z W.Effect of potassium application stage on photosynthetic characteristics of winter wheat flag leaves and on starch accumulation in wheat grains [J].ChineseJournalofAppliedEcology,2004,15(8)∶1349.
[6] MACDONALD P W,STROBEL G A.Adenosine diphosphate-glucose pyrophosphorylase control of starch accumulation in rust-infected wheat leaves [J].PlantPhysiology,1970,46(1):132.
[7] KALINGA D N,BERTOFT E,TETLOW I,etal.Evolution of amylopectin structure in developing wheat endosperm starch [J].CarbohydratePolymers,2014,112(2):316.
[8] RAHMAN S,LI Z,BATEY I,etal.Genetic alteration of starch functionality in wheat [J].JournalofCerealScience,2000,31:97,100.
[9] 高振宇,黄大年,钱 前.植物支链淀粉生物合成研究进展[J].植物生理与分子生物学学报,2004,30(5):490.
GAO Z Y,HUANG D N,QIAN Q.Advances in research on biosynthesis of plant amylopectin [J].JournalofPlantPhysiologyandMolecularBiology,2004,30(5):490.
[10] 姜宗庆,封超年,黄联联,等.施磷量对弱筋小麦扬麦9号籽粒淀粉合成和积累特性的调控效应[J].麦类作物学报,2006,26(6):81.
JIANG Z Q,FENG C N,HUANG L L,etal.Grain starch formation characteristics as affected by phosphorus application in weak-gluten wheat Yangmai 9 [J].JournalofTriticeaeCrops,2006,26(6):81.
[11] LI C Y,LI C,ZHANG R Q,etal. Effect of phosphorus on the characteristics of starch in winter wheat [J].Starch,2013,65:806.
[12] NI Y,WANG Z,YIN Y,etal.Starch granule size distribution in wheat grain in relation to phosphorus fertilization [J].JournalofAgriculturalScience,2012,150(1):45.
[13] 王旭东.磷对小麦产量和品质的影响及其生理基础研究[D].泰安:山东农业大学,2003:51-53.
WANG X D.Effect of phosphorus on kernel yield and quality and physiological basis in winter wheat [D].Tai’an:Shandong Agricultural University,2003:51-53.
[14] 赵永亮.一种同时测定小麦种子中直链淀粉、总淀粉含量的新方法-微量分光光度法[J].食品与发酵工业,2005,31(8):24.
ZHAO Y L.A novel micro-amount method with spectrophotometer to determine amylose and total starch content of wheat seeds [J].FoodandFermentationIndustries,2005,31(8):24.
[15] PENG M,GAO M,ABDEL-AAL E S M,etal.Separation and characterization of A- and B-type starch granules in wheat endosperm [J].CerealChemistry,1999,76(3):375.
[16] 贺 伟,丁 卉,王婕琛,等.高效阴离子交换色谱-脉冲安培法测定支链淀粉糖链长分布[J].分析测试学报,2012,31(10):1243.
HE W,DING H,WANG J C,etal.Determination of chain-length distribution of amylopectin by high performance ion exchange chromatography with pulsed amperometric detection [J].JournalofInstrumentalAnalysis,2012,31(10):1243.
[17] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000:268.
BAO S D.Soil and agro-chemistry analysis [M].Beijing:China Agriculture Press,2000:268.
[18] DOUGLAS C D,KUO T M,FELKER F C.Enzymes of sucrose and hexose metabolism in developing kernels of two inbreds of maize [J].PlantPhysiol,1988,86:1014.
[19] 赵法茂,齐 霞,肖 军,等.测定淀粉分支酶活性方法的改进[J].植物生理学通讯,2007,43(6):1168.
ZHAO F M,QI X,XIAO J,etal.Improved method for determining starch branching enzyme activity [J].PlantPhysiologyJournal,2007,43(6):1168.
[20] 刘 霞,谭胜兵,田纪春,等.不同淀粉组分含量小麦品种灌浆过程中淀粉去分支酶的活性及类型[J].中国农业科学,2010,43(4):851.
LIU X,TAN S B,TIAN J C,etal.Activity and types of starch debranching enzyme in wheat cultivars containing different starch contents during the grain filling period [J].ScientiaAgriculturaSinica,2010,43(4):851.
[21] 陈 静.小麦籽粒淀粉合成关键酶表达对磷素的响应[D].扬州:扬州大学,2010:33.
CHEN J.Expressions starch synthase response in wheat grain to phosphorus [D].Yangzhou:Yangzhou University,2010:33.
[22] WANG Z B,LI W H,QI J C,etal.Starch accumulation,activities of key enzyme and gene expression in starch synthesis of wheat endosperm with different starch contents [J].JournalofFoodScienceandTechnology,2011,51(3):419.
[23] 王树亮,田奇卓,李娜娜,等.不同小麦品种对磷素吸收利用的差异[J].麦类作物学报,2008,28(3):476.
WANG S L,TIAN Q Z,LI N N,etal.Differences of phosphorus utilization efficiency among different wheat varieties [J].JournalofTriticeaeCrops,2008,28(3):476.
[24] 姜宗庆.磷素对小麦产量和品质的调控效应及其生理机制[D].扬州:扬州大学,2006:5.
JIANG Z Q.Effects of phosphorus on yield and quality formation and physiological mechanism in wheat [D].Yangzhou:Yangzhou University,2006:5.
[25] 陈淑琴,路香彩,张耀泉,等.适宜两熟制种植的早熟冬小麦新品种冀麦37号的选育及应用[J].河北农业科学,1998,2(2):1.
CHEN S Q,LU X C,ZHANG Y Q,etal.Breeding and application of new early-variety Jimai 37 which fit for double-crop system area [J].JournalofHebeiAgriculturalScience,1998,2(2):1.
[26] 姚翠琴,苏敬国,李新杰,等.优质面包小麦新冬 23号品质分析及应用前景[J].种子,2010,29(5):92.
YAO C Q,SU J G,LI X J,etal.Quality analysis and application prospect of superior bread wheat Xindong 23 [J].Seed,2010,29(5):92.
[27] 李春艳,马 龙,张 宏,等.新疆冬小麦苗期耐低磷指标的筛选[J].麦类作物学报.2013,33(1):137.
LI C Y,MA L,ZHANG H,etal.Screening on indexes for tolerance to phosphorus stress at seedling stage of winter wheat [J].JournalofTriticeaeCrops,2013,33(1):137.
[28] 李春燕,封超年,王亚雷,等.不同小麦品种支链淀粉链长分配及其与淀粉理化特性的关系[J].作物学报,2007,33(8):1240.
LI C Y,FENG C N,WANG Y L,etal.Chain length distribution of debranched amylopectin and its relationship with physicochemical properties of starch in different wheat cultivars [J].ActaAgronomicaSinica,2007,33(8):1240.
