咸水灌溉棉田休耕期土壤胞外酶活性和微生物多样性研究
2018-05-02梁悦萍李科江张俊鹏王广帅孙景生段爱旺
梁悦萍,李科江,张俊鹏,王广帅,高 阳,孙景生,段爱旺*
(1.中国农业科学院农田灌溉研究所/农业部作物需水与调控重点实验室,河南 新乡453002;2.河北省农林科学院旱作农业研究所,河北 衡水 053000;3.山东农业大学水利土木工程学院,山东 泰安 271000;4.中国农业科学院研究生院,北京 100081)
土壤是陆地生态系统的核心[1]。微生物是土壤生物化学过程的重要参与者[2-3],其通过降解土壤中动物残体、植物凋落物以及自身呼吸作用,直接影响土壤关键化学元素循环。微生物群落多样性是表征土壤质量变化的敏感指标,对土壤管理具有重要指示作用[4-5]。另外,由微生物、动植物活体分泌及由动植物残体、遗骸分解释放于土壤中的土壤胞外酶是一类具有催化能力的生物活性物质,其活性反映了土壤生物化学过程的方向与强度[6-8]。土壤胞外酶活性强弱直接影响土壤相关养分元素的可利用性,是土壤健康状况的“指示器”[9]。农田是陆地生态系统的重要组成部分,全球耕地面积约占到陆地总面积的37.67%[10]。因此,研究农田土壤微生物群落多样性以及土壤胞外酶活性对于评价农田土壤健康状况、实现农田土壤科学管理至关重要。
灌溉是保障农田作物获得高产、稳产的重要农业管理措施。我国75%以上的粮食产自于灌溉农田,可以说农田灌溉是满足日益增长的人口口粮需求的基础保障[11-12]。然而,我国也是一个淡水资源短缺的国家,淡水资源短缺已经对许多地区农业生产形成严重威胁。在淡水资源短缺不断加剧的背景下,合理利用地下咸水资源进行灌溉已经成为农业生产中缓解淡水资源不足的较为经济、有效的手段[13]。咸水灌溉一方面提供了作物生长所需要的水分,缓解旱情,适度维持大田作物产量;另一方面其也将多种盐分带入土壤,影响土壤渗透性[14],造成土壤盐渍化[15],进而影响土壤的理化性质,造成土壤微生物活性发生改变[16-19]。例如:Wichern等[20]研究发现随着土壤盐分含量增加,土壤微生物活性指标值均下降;然而Wang等[21]却发现盐分处理能够促进土壤微生物量的增加。虽然普遍认为盐分胁迫是抑制微生物活性的主要非生物因素之一,但是部分耐受极端环境的微生物在盐胁迫环境下反而被底物诱导,表现出较高活性[22],从而使得不同盐分浓度胁迫下土壤微生物群落多样性以及活性表现出较高的不确定性。土壤胞外酶在关键化学元素循环转化过程中扮演着不可替代的关键角色,多数研究认为其活性随土壤盐分含量升高而降低[23-25],但王国栋等[26]却发现碱性磷酸酶和多酚氧化酶活性随土壤含盐量的增加而升高。因此,不同种类土壤胞外酶对盐分含量高低响应差异也很大。咸水灌溉引入农田土壤大量盐分,然而不同矿化度灌溉水导致土壤盐分含量各异,因此,开展不同矿化度灌溉水处理条件下土壤微生物群落多样性和胞外酶活性的研究对于准确评价咸水灌溉生态安全性和土壤健康状况显得尤为必要。
河北低平原是我国淡水资源严重短缺的地区之一。棉花作为耐盐抗旱能力较强的作物,在该区域种植面积非常大。翟红梅[27]和冯棣等[23]分别对该区域咸水灌溉土壤微生物群落特征和胞外酶活性做了比较系统的研究,但是集中于作物生长季,缺乏关注休耕期状况。休耕期土壤生物化学反应并未停止,为作物生长季储备大量养分物质,对于农田地力恢复十分重要。因此,本研究通过测定不同矿化度咸水灌溉棉田休耕期土壤微生物群落状况和胞外酶活性,分析盐分浓度对土壤微生物群落特征和胞外酶活性的影响,为区域咸水灌溉安全性评价提供数据支持。
1 材料与方法
1.1 试验地概括
试验平台位于河北省深州市的河北省农林科学院旱作节水农业试验站(37°44′N,115°47′E)内,海拔21 m。该试验站地处河北平原中部,属暖温带大陆性季风气候,多年平均气温12.8℃,无霜期188 d,日照时数2 509.4 h,多年平均蒸发量1 785.4 mm,降雨量500.3 mm且70%集中于夏季(6—8月)。试验区土壤质地为壤土,地下水埋深大于5 m。土壤1 m土层平均田间持水率为28%,土壤容重为1.44 g·cm-3;土壤表层 0~20 cm 有机质含量为 11.5 g·kg-1,速效氮含量76 mg·kg-1,速效磷含量 15 mg·kg-1,速效钾含量 112 mg·kg-1[28]。
1.2 试验设计
试验平台灌溉方式为畦灌,依据灌溉水的矿化度设置了6个处理,每个处理3次重复,共18个小区,每个试验小区面积37.62 m2(5.7 m×6.6 m),采用完全随机区组设计。6个处理的灌溉水矿化度分别为淡水、2、4、6、8 g·L-1和 10 g·L-1,其中,淡水取自当地深层地下水,其他矿化度灌溉水由深层地下水掺兑海盐配制而成。灌水下限均控制在田间持水率的65%。试验采用棉花-休耕的种植模式,棉花栽培管理参见冯棣等[23]。用于培养与分析的土壤样品采集于2015年11月9日,处于农田休耕期。按照“Z”字形用直径2.5 cm土钻取表层土(0~20 cm),共10个点,混匀后以四分法取样。由于采样时气温较低,故采集的土壤样品在田间过2 mm网筛,手工去除可见的植物根系和有机质残余,运送至实验室,放置在4℃冰箱以备测定使用。
1.3 主要测定指标与方法
1.3.1 土壤微生物群落多样性
采用含有31种碳源的Biolog ECO微平板分析微生物群落特征。首先,将待测土壤样品在25℃培养箱培养活化1周;配制0.85%的氯化钠溶液并在121℃高温灭菌锅中灭菌30 min;称取1 g土壤样品置于150 mL三角瓶中,加入99 mL 0.85%的氯化钠溶液,将三角瓶以200 r·min-1的速度振荡30 min。待前一过程制备的土壤溶液静置20 min后,无菌条件下,吸取1 mL土壤悬浊液加入9 mL 0.85%的氯化钠溶液,继续吸取上述稀释10倍的土壤悬浊液1 mL,加入9 mL 0.85%的氯化钠溶液使土壤溶液稀释100倍,测定其OD590值,在0.13±0.02范围内(若大于0.13±0.02,则继续稀释10倍,使其稀释倍数为1000倍)。最后,将稀释土壤溶液摇匀,无菌条件下使用电动排枪吸取上清液150 μL加入Biolog ECO微平板,读板。前3 d每隔12 h读一次板,4 d后每隔24 h读一次板。本试验采用BIOLOG公司的EL808微孔板读数仪在590 nm波长下对ECO板进行数据读取,读板时间分别为 12、24、36、48、72、96、120、144、168 h 和192 h。平均颜色变化率是反映土壤微生物代谢活性,即利用单一碳源能力的指标,相关计算公式参见邹春娇等[29]。
1.3.2 土壤胞外酶活性
称取新鲜土壤样品2.00 g,将土样装入120 mL振荡瓶并加入100 mL Tris碱性缓冲溶液,以200 r·min-1的速度振荡30 min,之后静止30 min,用8通道排枪取100 μL土壤溶液加入96孔板,并加待测土壤胞外酶所对应的荧光标记底物100 μL,放入25℃培养箱黑暗培养3 h;将96孔板从培养箱取出,加50 μL的0.5 mol·L-1的NaOH溶液终止反应。利用酶标仪(帝肯,Infinite-200Pro,瑞士)在激发波长 365 nm 和发射波长450 nm下读数,部分试验步骤参见Jing等[30]。土壤胞外酶活性以nmol·g-1·h-1(DW)来表示,性质见表1。

表1 本试验涉及土壤胞外酶性质Table 1 Fluorescent substrates of soil extracellular enzymes used in this experiment
1.4 数据处理
采用SPSS 22.0软件进行方差分析(ANOVA)和多重比较(LSD),α=0.05;利用 Data Reduction工具对96 h的平均吸光度(AWCD)值进行主成分分析。采用SigmaPlot 12.5软件进行作图。
2 结果与分析
2.1 咸水灌溉对土壤胞外酶活性的影响
由图1 a可以看出,不同矿化度咸水灌溉对土壤α葡萄糖苷酶活性存在显著影响。与淡水灌溉处理相比,高矿化度(6 g·L-1和 10 g·L-1)灌溉水处理能够显著降低α葡萄糖苷酶的活性,但低矿化度(2 g·L-1和4 g·L-1)和高矿化度(8 g·L-1)灌溉水处理仅有降低 α葡萄糖苷酶活性的趋势,统计学上不显著。不同矿化度咸水灌溉处理之间也存在显著差异,其中在2、4 g·L-1和 8 g·L-1处理下,土壤 α 葡萄糖苷酶活性显著高于 10 g·L-1处理,同时,4 g·L-1处理也显著高于 6 g·L-1处理。通过图1b、图1c可以发现不同矿化度灌溉水处理并没有对土壤β葡萄糖苷酶活性和乙酰氨基葡萄糖苷酶活性产生显著影响(P>0.05)。通过图1d可以看出,纤维素二糖酶对咸水灌溉响应强烈,且纤维素二糖酶活性随着灌溉水矿化度升高而显著降低。与淡水灌溉处理相比,高矿化度(6、8 g·L-1和 10 g·L-1)灌溉水处理显著降低了土壤纤维素二糖酶活性,降幅分别达到了53.25%、56.50%和81.03%;而低矿化度(2 g·L-1和 4 g·L-1)灌溉水处理条件下仅有降低的趋势,统计学上不显著;对不同矿化度咸水灌溉处理之间比较后发现,土壤纤维素二糖酶活性在2 g·L-1处理下显著高于高矿化度(6、8 g·L-1和 10 g·L-1)灌溉水处理,而4 g·L-1灌溉水处理则显著高于高矿化度(8 g·L-1和 10 g·L-1)灌溉水处理。由图 1e 可以看出,随着灌溉水矿化度的升高,土壤磷酸酶的活性逐步减弱。与淡水灌溉处理相比,灌溉水矿化度增加均能显著降低土壤磷酸酶的活性;并且不同矿化度灌溉水处理之间同样存在显著性差异,其中2 g·L-1灌溉水处理显著高于高矿化度(6、8 g·L-1和 10 g·L-1)灌溉水处理,4 g·L-1灌溉水处理则显著高于高矿化度(8 g·L-1和 10 g·L-1)灌溉水处理,而 6 g·L-1灌溉水处理仅高于 10 g·L-1灌溉水处理下的土壤磷酸酶活性,8 g·L-1处理和10 g·L-1处理之间无显著差异。由图1f可以看出,β木糖苷酶活性显著受到灌溉水矿化度的影响。与淡水灌溉处理相比,虽然灌溉水矿化度增加能够降低土壤β木糖苷酶活性,但仅有高矿化度(8 g·L-1和10 g·L-1)灌溉水处理达到了显著水平,随着灌溉水矿化度增加,咸水灌溉处理之间土壤β木糖苷酶活性差异逐渐显著。

图1 不同矿化度灌溉水处理对土壤胞外酶活性的影响Figure 1 Effects of saline water irrigation on soil enzyme activity
2.2 咸水灌溉对土壤微生物群落的影响
2.2.1 平均颜色变化率
平均吸光度(AWCD)随时间的变化可以用来表征土壤微生物对碳源利用水平的高低,能够直观地体现土壤微生物群落对不同碳源的响应速度和最终达到的程度。不同矿化度咸水灌溉处理下AWCD随培养时间的动态变化曲线如图2所示。可以看出AWCD在培养初期(12 h)均较低,但随着时间的推移,低矿化度(淡水、2 g·L-1和 4 g·L-1)灌溉水处理 AWCD 随着时间的推移逐步增大,到136 h后微生物活性趋于平稳;但是高矿化度(6、8 g·L-1和 10 g·L-1)灌溉水处理下,AWCD随着时间的推移无明显变化趋势。与淡水灌溉处理相比,2 g·L-1处理显著增加了AWCD值,增幅近80%。而其他矿化度灌溉水处理均显著降低土壤微生物AWCD值,并且减弱作用随灌溉水矿化度的增加而加强。
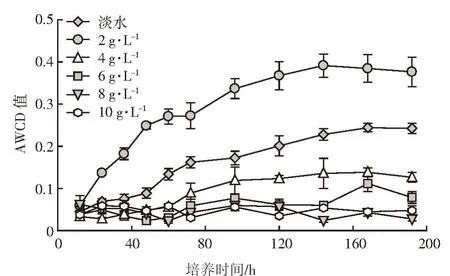
图2 不同咸水灌溉处理下土壤微生物平均吸光度(AWCD)变化特征Figure 2 Average well color development(AWCD)of Biolog ECO plates under different saline water irrigation treatments
2.2.2 群落多样性分析
根据Biolog ECO培养板所有微孔光密度数据,参考相关计算公式对不同矿化度咸水灌溉条件下的土壤微生物多样性进行计算,结果见表2。从表2可以看出,在不同咸水灌溉处理下,4种土壤微生物多样性指数均存在显著差异。在8 g·L-1处理下Shannon-wiener指数(H′)值显著高于低矿化度咸水灌溉(2 g·L-1和 4 g·L-1)处理和淡水灌溉处理;其总体上呈现从淡水处理到咸水灌溉处理先降低再增高而后又降低的趋势。2 g·L-1处理的 Mcintosh指数(U)值显著高于其他处理,其次是淡水处理高于高矿化度(6、8 g·L-1和 10 g·L-1)处理,总体上呈现从淡水处理到咸水灌溉处理先增高后下降的趋势。对于Simpson指数(D),最大值出现在 8 g·L-1处理,其次是 10 g·L-1处理,淡水灌溉与各个咸水灌溉处理均无显著性差异,但是咸水灌溉处理之间的D值存在显著差异。从淡水到咸水灌溉处理来看,总体呈现先下降后增高而后又下降的趋势。2 g·L-1处理下丰富度指数最高,其次是淡水灌溉处理,随着灌溉水的矿化度增加丰富度指数急剧下降。
2.2.3 碳源利用性特点分析
为研究不同矿化度咸水灌溉对土壤微生物群落碳源利用影响的特点,选择96 h作为取样分析点进行主成分分析。分别提取两个主成分,第一主成分(PC1)和第二主成分(PC2)。样方间距离的长短表征处理之间的相似程度,距离越接近表示相似度越高。图3是不同矿化度咸水灌溉处理条件下土壤微生物群落多样性主成分分析结果。可以看出,2 g·L-1处理分布在第一象限,与其他咸水灌溉(4、6、8 g·L-1和 10 g·L-1)处理分布在第四象限的土壤微生物群落多样性存在显著区别;淡水处理分布在第三象限,也和2 g·L-1处理之间存在明显差异。相比淡灌溉处理,咸水灌溉对土壤微生物群落产生的影响存在差异,并且不同矿化度处理之间也存在显著差异。
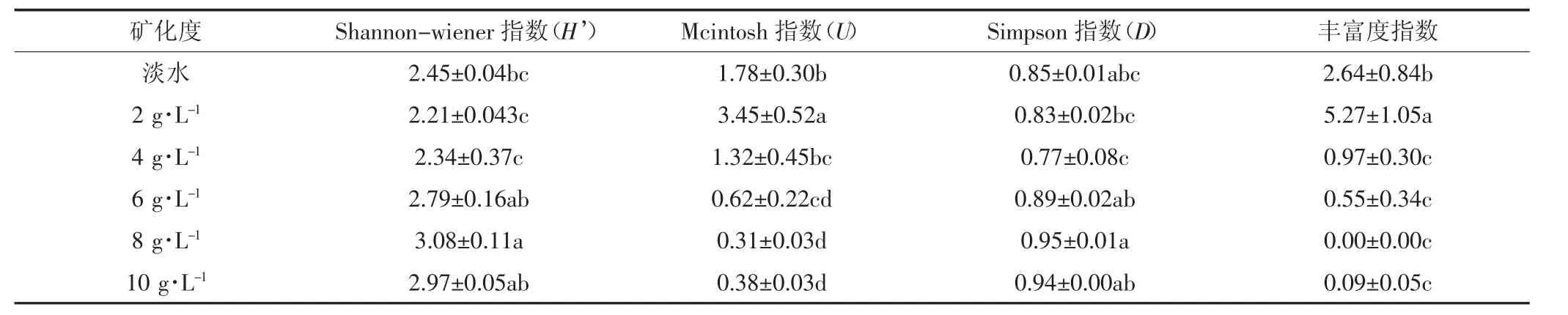
表2 不同矿化度咸水灌溉模式下微生物群落多样性指数Table 2 Microorganism diversity under different saline water irrigation treatments
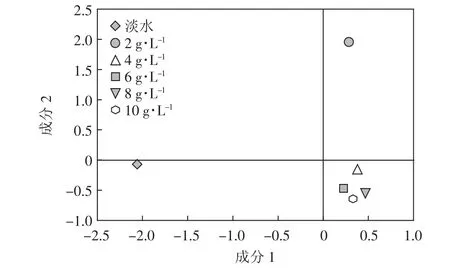
图3 不同盐分梯度咸水灌溉下土壤微生物群落多样性主成分分析Figure 3 Principal analysis of the function diversity of saline-irrigated soil
按照底物碳源化学基团的性质,Biolog Eco板上的31种碳源划分为6大类:糖类、氨基酸类、胺类、羧酸类、酯类以及醇类。将不同矿化度咸水灌溉处理下的6种碳源分别计算AWCD值见图4。就醇类来讲,低矿化度咸水灌溉(2 g·L-1和 4 g·L-1)能够显著促进微生物对醇类的利用,并且利用率随时间的推移而增大,到120 h时处于稳定状态,其中2 g·L-1处理下的AWCD值也显著高于4 g·L-1处理。淡水处理的醇类底物利用能力显著低于低矿化度咸水灌溉(2 g·L-1和4 g·L-1)处理,但却显著高于高矿化度咸水灌溉(6、8 g·L-1和 10 g·L-1)处理,6 g·L-1处理下的 AWCD 值高于 10 g·L-1处理(图 4a)。就酯类来讲,2 g·L-1处理下AWCD值最高,并且也具有随时间延续而增大的趋势,到144 h后趋于稳定,其次是淡水下的AWCD值,该值显著低于2 g·L-1处理,但显著高于其他矿化度处理(4、6、8 g·L-1和 10 g·L-1)。对于糖类来讲,咸水灌溉随着矿化度的增加,有先促进后减弱微生物对糖类碳源利用的趋势。其中,相对于淡水处理,2 g·L-1处理能够显著促进土壤微生物对糖类碳源的利用(图4c),而其余矿化度咸水灌溉均显著减弱了土壤微生物对糖类碳源的利用。对于羧酸类来讲,2 g·L-1处理下的微生物AWCD依然最高,且随时间的推移而增大,到达72 h时保持稳定。通过统计发现,除2 g·L-1的微生物AWCD最高外,淡水处理下土壤微生物AWCD依然高于4 g·L-1处理和高矿化度咸水灌溉处理(6、8 g·L-1和 10 g·L-1),并且 4 g·L-1处理下的AWCD也高于高矿化度咸水灌溉处理。对于氨基酸类来讲,淡水灌溉和2 g·L-1咸水灌溉处理下土壤微生物AWCD值最高且之间无差异,其余咸水灌溉处理显著低于两者,但是6 g·L-1处理下土壤微生物AWCD最低。对于胺类来讲,2 g·L-1和 6 g·L-1处理下土壤微生物AWCD值明显高于其他处理,并且2 g·L-1处理显著高于6 g·L-1处理下的土壤微生物AWCD值,其余处理AWCD值均较小,仅有淡水处理下的AWCD超过了0.05。不同盐分梯度咸水灌溉条件下,土壤微生物对6种碳源的利用由强到弱依次为醇类>氨基酸类>羧酸类>糖类>胺类>酯类。
3 讨论
3.1 咸水灌溉对土壤胞外酶活性的影响
土壤盐渍化是影响土壤胞外酶活性的主要因素之一。本研究结果表明多种土壤胞外酶活性随着咸水灌溉矿化度的增加而减弱,这可能是由于咸水灌溉将大量盐分引入土壤,抑制了土壤微生物活性,减少了微生物数量,从而致使活体微生物分泌酶的数量减少。长期咸水灌溉对土壤微环境产生的负面影响不容忽视,应是将来水资源短缺加剧状况下咸水资源开发利用环境评价的重要内容。虽然咸水灌溉对多种土壤胞外酶都有抑制作用,但是抑制程度随胞外酶种类的变化有很大差异。咸水灌溉矿化度对土壤纤维素二糖酶活性、磷酸酶活性以及β木糖苷酶活性的抑制作用较强,而对α葡萄糖苷酶活性和β葡萄糖苷酶活性的抑制作用相对较弱。另外,本研究还发现咸水灌溉对乙酰氨基葡萄糖苷酶活性有促进作用。这可能是由于产生不同胞外酶所对应的微生物种类不同,而这些微生物对土壤盐分的耐受性也不同。土壤理化性质是土壤微生物活性的直接影响因素,咸水灌溉也可通过改变土壤理化性质而间接地影响微生物活性,乃至胞外酶活性。例如:Min等[33]发现微咸水和咸水灌溉处理下土壤脲酶活性显著高于淡水处理,就是因为微咸水和咸水灌溉显著改变了土壤理化性质,从而导致脲酶活性增加。Zahran[34]指出耐盐型细菌所产生的土壤酶活性高于不耐盐型细菌所产生的相应土壤酶的活性,这表明胞外酶活性强弱和相应微生物的耐盐性有关。休耕期土壤胞外酶活性与生长季测定数据(未列出)相比,没有显著性差异。冯棣等[23]在该区域相关研究也发现了类似的结果。这说明休耕期土壤胞外酶仍然处于高度活化状态,对土壤关键养分元素的周转并没有停止,休耕期对农田土壤储能、休养生息非常关键。咸水灌溉引入土壤大量盐分离子,对土壤微生物细胞生理和代谢过程产生胁迫[19],影响了微生物活性[35]和微生物数量[36],也可能改变土壤理化性质[37],从而直接或间接地改变土壤胞外酶活性,其中有些酶对土壤盐分含量不敏感,但是土壤α葡萄糖苷酶、纤维素二糖酶、磷酸酶和β木糖苷酶对灌溉水矿化度反应相对敏感,亦可作为咸水灌溉棉田土壤质量评价的灵敏指标。

图4 咸水灌溉土壤对不同碳源的利用Figure 4 Utilization of carbon sources by soil microorganisms under different saline water irrigation treatments
3.2 咸水灌溉对土壤微生物群落的影响
较低水平盐分咸水灌溉对微生物活性有促进作用,而较高水平的盐分则显著抑制土壤微生物的活性[38]。本研究发现土壤微生物代谢水平(AWCD值)在矿化度较低的灌溉处理下较高,而随着灌溉水矿化度的增加而下降。与本研究同一站点的研究也发现相似结果,相比对照处理,2.5 dS·m-1矿化度灌溉水处理并没有降低土壤细菌丰度,而随着灌溉水矿化度升高,土壤细菌数量显著降低,10 dS·m-1矿化度灌溉水处理相比对照下降高达50.2%[27]。这主要是因为较低矿化度咸水灌溉能够促进土壤微生物产生应激反应,刺激微生物数量的增加和活性的加强。另外,有关咸水灌溉棉田土壤微生物群落特征的研究却发现,土壤微生物群落AWCD值、土壤微生物群落多样性Shannon指数和丰富度指数均随盐浓度的升高而逐渐降低[39],而本研究结果却发现棉田土壤微生物群落多样性Shannon指数和丰富度指数随盐浓度的升高有先增高后降低的趋势,可能是由两个实验土壤性质差别较大所致。这也说明低矿化度咸水灌溉或微咸水灌溉对土壤微生物群落多样性影响存在较大不确定性。今后,应该加强微咸水灌溉对土壤理化性质以及微生物环境的影响方面的研究。翟红梅[27]在与本研究同一站点开展的冬小麦咸水灌溉试验发现,长期咸水灌溉对土壤细菌多样性影响结果与本研究结果类似,这可能是由于小尺度环境下土壤微生物群落组成较为相近。本研究发现较高矿化度咸水灌溉显著降低了土壤微生物群落组成,但低矿化度咸水灌溉对土壤微生物群落组成的影响随地域等因素变化而不同。
4 结论
(1)不同矿化度咸水灌溉显著改变了土壤胞外酶活性。土壤盐分含量状况对土壤胞外酶活性有促进或抑制作用,说明咸水灌溉对土壤健康状况的影响较为复杂。休耕期土壤胞外酶活性仍然保持较高水平,农田实行冬季休耕,仍能保持较高的养分元素转化效率,促进土壤养分元素积累,对于提升土壤质量十分有利。
(2)不同矿化度咸水灌溉对土壤微生物群落多样性影响不同。低矿化度灌溉水可以促进土壤微生物产生应激效应,促使微生物快速生长并加快土壤营养元素转化速率;而高矿化度灌溉水则会抑制土壤微生物生长和养分元素转化。水分短缺背景下,适当实施微咸水灌溉可能会提高棉田土壤微生物活性和养分元素转化速率。
参考文献:
[1]王绍强,周成虎.中国陆地土壤有机碳库的估算[J].地理研究,1999,18(4):349-356.WANG Shao-qiang,ZHOU Cheng-hu.Estimating soil carbon reservior of terrestrial ecosystem in China[J].Geographical Research,1999,18(4):349-356.
[2]O′donnell A G,Seasman M,Macrae A,et al.Plants and fertilisers as drivers of change in microbial community structure and function in soils[J].Plant and Soil,2001,232(1/2):135-145.
[3]姚槐应,黄昌勇.土壤微生物生态学及其实验技术[M].北京:中国科学出版社,2006.YAO Huai-ying,HUANG Chang-yong.Soil microbial ecology and its experimental techniques[M].Beijing:China Science Press,2006.
[4]吴则焰,林文雄,陈志芳,等.武夷山国家自然保护区不同植被类型土壤微生物群落特征[J].应用生态学报,2013,24(8):2301-2309.WU Ze-yan,LIN Wen-xiong,CHEN Zhi-fang,et al.Characteristics of soil microbial community under different vegetation types in Wuyishan National Nature Reserve,East China[J].Chinese Journal of Applied E-cology,2013,24(8):2301-2309.
[5]侯晓杰,汪景宽,李世朋.不同施肥处理与地膜覆盖对土壤微生物群落功能多样性的影响[J].生态学报,2007,27(2):655-661.HOU Xiao-jie,WANG Jing-kuan,LI Shi-peng.Effects of different fertilization and plastic-mulching on functional diversity of soil microbial community[J].Acta Ecologica Sinica,2007,27(2):655-661.
[6]张咏梅,周国逸,吴 宁.土壤酶学的研究进展[J].热带亚热带植物学报,2004,1(1):83-90.ZHANG Yong-mei,ZHOU Guo-yi,WU Ning.A review of studies on soil enzymology[J].Journal of Tropical and Subtropical Botany,2004,1(1):83-90.
[7]刘 远,潘根兴,张 辉,等.大气CO2浓度和温度升高对麦田土壤呼吸和酶活性的影响[J].农业环境科学学报,2017,36(8):1484-1491.LIU Yuan,PAN Gen-xing,ZHANG Hui,et al.Effects of elevated atmospheric CO2concentration and temperature on soil respiration and enzyme activity in a wheat field[J].Journal of Agro-Environment Science,2017,36(8):1484-1491.
[8]王 进,杨明凤,褚贵新.外源砷在石灰性土壤中的形态与土壤酶活性研究[J].农业机械学报,2016,47(11):179-184.WANG Jin,YANG Ming-feng,CHU Gui-xin.Form and soil enzyme activities of exogenous arsenic in calcareous soil[J].Transactions of the Chinese Society for Agricultural Machinery,2016,47(11):179-184.
[9]Nannipieri P,Giagnoni L,Renella G,et al.Soil enzymology:Classical and molecular approaches[J].Biology and Fertility of Soils,2012,48(7):743-762.
[10]Food and Agricultural Organization of the United Nations.FAOSTAT Data[EB/OL].[2017-11-01].http://www.fao.org/faostat/en/#data.
[11]Du T S,Kang S Z,Zhang J H,et al.Deficit irrigation and sustainable water-resource strategies in agriculture for China′s food security[J].Journal of Experimental Botany,2015,66(8):2253-2269.
[12]Wang G S,Liang Y P,Zhang Q,et al.Mitigated CH4and N2O emissions and improved irrigation water use efficiency in winter wheat field with surface drip irrigation in the North China Plain[J].Agricultural Water Management,2016,163:403-407.
[13]王艳娜,侯振安,龚 江,等.咸水资源农业灌溉应用研究进展与展望[J].中国农学通报,2007,23(2):393-397.WANG Yan-na,HOU Zhen-an,GONG Jiang,et al.Development and expectation of utilization of saline water resources in agriculture irrigation[J].Chinese Agricultural Science Bulletin,2007,23(2):393-397.
[14]张余良,陆文龙,张 伟,等.长期微咸水灌溉对耕地土壤理化性状的影响[J].农业环境科学学报,2006,25(4):969-973.ZHANG Yu-liang,LU Wen-long,ZHANG Wei,et al.Effects of long term brackish water irrigation on characteristics of agrarian soil[J].Journal of Agro-Environment Science,2006,25(4):969-973.
[15]Pereira L S,Oweis T,Zairi A.Irrigation management under water scarcity[J].Agricultural Water Management,2002,57(3):175-206.
[16]Jin Z Z,Li J Q,Xu S Y,et al.Characteristics of sandy soil microbial metabolisms in the forests drip irrigation with saline water[J].Journal of Desert Research,2014,34:363-370.
[17]Elgharably A,Marschner P.Microbial activity and biomass and N and P availability in a saline sandy loam amended with inorganic N and lupin residues[J].European Journal of Soil Biology,2011,47(5):310-315.
[18]Ghollarata M,Raiesi F.The adverse effects of soil salinization on the growth of Trifolium alexandrinum L.and associated microbial and biochemical properties in a soil from Iran[J].Soil Biology and Biochemistry,2007,39(7):1699-1702.
[19]Sardinha M,Muller T,Schmeisky H,et al.Microbial performance in soils along a salinity gradient under acidic conditions[J].Applied Soil Ecology,2003,23(3):237-244.
[20]Wichern J,Wichren F,Joergensen R G.Impact of salinity on soil microbial communities and the decomposition of maize in acidic soils[J].Geoderma,2006,137(1/2):100-108.
[21]Wang V N L,Dalal R C,Greene R S B.Salinity and sodicity effects on respirationandmicrobialbiomassofsoil[J].BiologyandFertilityofSoils,2008,44(7):943-953.
[22]贺纪正,张丽梅.氨氧化微生物生态学与氮循环研究进展[J].生态学报,2009,29(1):406-415.HE Ji-zheng,ZHANG Li-mei.Advances in ammonia-oxidizing microorganisms and global nitrogen cycle[J].Acta Ecologica Sinica,2009,29(1):406-415.
[23]冯 棣,张俊鹏,孙池涛,等.长期咸水灌溉对土壤理化性质和土壤酶活性的影响[J].水土保持学报,2014,28(3):171-176.FENG Di,ZHANG Jun-peng,SUN Chi-tao,et al.Effects of long-term irrigation with saline water on soil physical-chemical properties and activitiesofsoilenzyme[J].JournalofSoilandWaterConservation,2014,28(3):171-176.
[24]Tripathi S,Chakraborty A,Chakrabarti K,et al.Enzyme activities and microbial biomass in coastal soils of India[J].Soil Biology and Biochemistry,2007,39(11):2840-2848.
[25]窦超银,康跃虎,万书勤,等.覆膜滴灌对地下水浅埋区重度盐碱地土壤酶活性的影响[J].农业工程学报,2010,26(3):44-51.DOU Chao-yin,KANG Yue-hu,WAN Shu-qin,et al.Effect of mulchdrip irrigation on soil enzyme activities of saline-sodic soil with shallow water table[J].Transactions of the Chinese Society of Agricultural Engineering,2010,26(3):44-51.
[26]王国栋,褚贵新,刘 瑜,等.干旱绿洲长期微咸地下水灌溉对棉田土壤微生物量影响[J].农业工程学报,2009,25(11):44-48.WANG Guo-dong,CHU Gui-xin,LIU Yu,et al.Effects of long-term irrigation with brackish groundwater on soil microbial biomass in cotton field in arid oasis[J].Transactions of the Chinese Society of Agricultural Engineering,2009,25(11):44-48.
[27]翟红梅.咸水灌溉对土壤微生物活性和多样性的影响[D].北京:中国科学院,2012.ZHAI Hong-mei.Impact of irrigation with saline water on the activities and diversity of microorganism in soil[D].Beijing:Chinese Academy of Sciences,2012.
[28]张俊鹏.咸水灌溉覆膜棉田水盐运移规律及耦合模拟[D].北京:中国农业科学院,2015.ZHANG Jun-peng.Coupling simulation of soil water-salt movement in plastic film mulched cotton field under saline water irrigation[D].Beijing:Chinese Academy of Agricultural Sciences,2015.
[29]邹春娇,齐明芳,马 建,等.Biolog-ECO解析黄瓜连作营养基质中微生物群落结构多样性特征[J].中国农业科学,2016,49(5):942-951.ZOU Chun-jiao,QI Ming-fang,MA Jian,et al.Analysis of soil microbial community structure and diversity in cucumber continuous cropping nutrition medium by Biolog-ECO[J].Scientia Agricultura Sinica,2016,49(5):942-951.
[30]Jing X,Wang Y H,Chung H,et al.No temperature acclimation of soil extracellular enzymes to experimental warming in an alpine grassland ecosystem on the Tibetan Plateau[J].Biogeochemistry,2013,117(1):39-54.
[31]Amirjani M R.Effect of salinity stress on growth,mineral composition,proline content,antioxidant enzymes of soybean[J].American Journal of Plant Physiology,2010,5(6):350-360.
[32]张前前,王 飞,刘 涛,等.微咸水滴灌对土壤酶活性、CO2通量及有机碳降解的影响[J].应用生态学报,2015,26(9):2743-2750.ZHANG Qian-qian,WANG Fei,LIU Tao,et al.Effects of brackish water irrigation on soil enzyme activity,soil CO2flux and organic matter decomposition[J].Chinese Journal of Applied Ecology,2015,26(9):2743-2750.
[33]Min W,Guo H J,Zhang W,et al.Irrigation water salinity and N fertilization:Effects on ammonia oxidizer abundance,enzyme activity and cotton growth in a drip irrigated cotton field[J].Journal of Integrative Agriculture,2016,15(5):1121-1131.
[34]Zahran H H.Diversity,adaptation and activity of the bacterial flora in saline environments[J].Biology and Fertility of Soils,1997,25(3):211-223.
[35]Nan Y,Maschner P.Microbial activity and biomass recover rapidly after leaching of saline soils[J].Biology and Fertility of Soils,2013,49(3):367-371.
[36]Tripathi S,Kumari S,Chakraborty A,et al.Microbial biomass and its activities in salt-affected coastal soils[J].Biology and Fertility of Soils,2006,42(3):273-277.
[37]时唯伟,支月娥,王 景,等.土壤次生盐渍化与微生物数量及土壤理化性质研究[J].水土保持学报,2009,23(6):166-170.SHI Wei-wei,ZHI Yue-e,WANG Jing,et al.Secondary salinization of greenhouse soil and its effects on microbe number and soil physicochemical properties[J].Journal of Soil and Water Conservation,2009,23(6):166-170.
[38]郑丽萍,龙 涛,林玉锁,等.Biolog-ECO解析有机氯农药污染场地土壤微生物群落功能多样性特征[J].应用与环境生物学报,2013,19(5):759-765.ZHENG Li-ping,LONG Tao,LIN Yu-suo,et al.Biolog-ECO analysis of microbial community functional diversity in organochlorine contaminated soil[J].Chinese Journal of Applied and Environmental Biology,2013,19(5):759-765.
[39]路海玲.土壤盐分对棉田土壤微生物活性和土壤肥力的影响[D].南京:南京农业大学,2011.LU Hai-ling.Effects of salinity on soil microbial activity and soil fertility in cotton field[D].Nanjing:Nanjing Agricultural University,2011.
