外施Ca2+对盐胁迫下珠美海棠组培苗光合特性的影响
2018-04-24秦家慧李颖华范航伟孙敏彭立新
秦家慧 李颖华 范航伟 孙敏 彭立新


摘 要:为研究Ca2+对盐胁迫下植株伤害的缓解作用,以珠美海棠组培苗为材料,采用盆栽法研究外施Ca2+对盐胁迫下珠美海棠光合特性的影响。采用双因素随机区组设计,NaCl设置0,100,200,300 mmol·L-14个浓度梯度,Ca2+设置0,5,10,15, 20 mmol·L-1 5个浓度梯度,对各处理珠美海棠色素和光合指标进行测定和分析。结果表明:(1)单一NaCl胁迫和外施Ca2+处理均可使珠美海棠叶绿素a和b含量呈先升高后下降的趋势,其中在NaCl和Ca2+浓度分别达到200 mmol·L-1和15 mmol·L-1时达到最高值;(2)单一NaCl胁迫可使珠美海棠Pn、Gs和Tr呈先升高后下降的趋势,Ci趋势与三者相反,其中在NaCl浓度达到100 mmol·L-1时Pn、Gs和Tr达到最高值而Ci达到最低值;(3)在NaCl高浓度即200 mmol·L-1和300 mmol·L-1处理,外施适宜浓度Ca2+对盐胁迫下珠美海棠光合指标有缓解作用,并分别以外施15,10 mmol·L-1 Ca2+效果最佳。研究可为珠美海棠在滨海盐碱地的引种栽培提供参考和借鉴。
关键词:珠美海棠;盐胁迫;外施Ca2+;光合特性
中图分类号:S661.4 文献标志码:A DOI 编码:10.3969/j.issn.1006-6500.2018.04..001
Effects of Exogenous Ca2+ on Photosynthetic Characteristics of Tissue Cultured Seedlings of Malus zumi under Salt Stress
QIN Jiahui, LI Yinghua, FAN Hangwei, SUN Min, PENG Lixin
(College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300384, China)
Abstract: To study the mitigation effect of Ca2+ on plant injury under salt stress, the effects of additional Ca2+ on photosynthetic characteristics of Malus zumi under salt stress were studied by pot culture method. Two factors randomized block design was used, which included 4 NaCl concentration gradient (0,100,200,300 mmol·L-1), and 5 Ca2+ concentration gradient (0,5,10,15,20 mmol·L-1), the pigment and photosynthetic indices of M. zumi were measured and analyzed. The results showed as follows: (1) under single NaCl stress or Ca2+ treatments, the chlorophyll a and b contents of Malus zumi were increased and then decreased and the maximum was at 200 mmol·L-1 and 15 mmol·L-1, respectively; (2) Under single NaCl stress, the Pn, Gs and Tr of Malus zumi were also increased and then decreased while the trend of Ci was contrary, the maximum of Pn, Gs and Tr or the minimum of Ci was at NaCl 100 mmol·L-1; (3) When NaCl concentration was 200 mmol·L-1 and 300 mmol·L-1, the suitable Ca2+ could relieve the injury of salt on photosynthetic indices of Malus zumi, and the optimum Ca2+ were 15 and 10 mmol·L-1 respectively. The results could provide reference for introduction and cultivation of Malus zumi in coastal saline-alkali land.
Key words: Malus zumi; salt stress; additional Ca2+; photosynthetic characteristics
土壤鹽渍化是当今世界耕地退化和土地荒漠化的主要因素之一,目前,全球盐碱地面积已达9.5×108 hm2[1],潜在的盐渍化土壤约0.17×108 hm2,如何充分开发和合理利用或改良盐碱地,成为当代农业面临的主要问题之一。盐胁迫会影响植物生长发育过程[2],MUNNS[3]将盐胁迫影响植物生长发育主要概括为3个方面:盐胁迫条件下的低水势是影响植物多种生理生化过程的根本原因,引起植物叶片水势下降,导致气孔导度下降;盐胁迫通过降低光合速率、减少能量和同化物的供给,限制植物生长发育;盐胁迫还会影响某些特定的酶及其代谢过程,从而影响植物生长发育。
Ca2+是偶联细胞外信号的第二信使,不仅可以维持细胞膜结构及细胞膜结合蛋白的稳定性,而且在调控多种酶活性方面具有重要作用[4-5]。在营养液中施加Ca2+能起到降低盐胁迫所导致的氧化损伤,从而保持较高的酶活性,保护光系统Ⅱ并维持较高的光合效率[6]。盐胁迫会通过影响光合作用碳同化的进程降低碳同化效率,减少植物有机物的积累。因此,施加一定浓度的Ca2+可调节因Ca2+缺失导致的植物非正常性生长发育,维持钙信号系统的正常发生及传递,从而改善植物光合能力。因此, Ca2+在植物抗逆过程中的应用受到普遍关注[7]。
珠美海棠(Malus zumi)是一种落叶小乔木或灌木,原产于日本北部山区,属于毛山荆子和三叶海棠的野生杂交种,1976年引入我国[8],1981年在天津农学院组培取得成功,因其抗逆性强,耐盐碱,可用作砧木嫁接苹果,在盐碱地区苹果的生产中具有重要意义。
通过研究珠美海棠在盐胁迫下,外施不同浓度Ca2+对其光合特性的影响,旨在为珠美海棠在盐碱地的应用和推广提供理论依据。
1 材料和方法
1.1 试验材料
以天津农学院果树学重点实验室提供的珠美海棠组培生根苗为试验材料。
1.2 试验设计
将珠美海棠组培生根苗练苗7 d左右(培养箱3 d,室内4 d),移栽到盛有基质(蛭石∶珍珠岩∶草炭土=1∶1∶1)的直径约为10 cm的花盆中,待幼苗在室温下生长45 d左右,长出8~10片新叶,并且茎部开始木质化时,进行盐胁迫,同时外施Ca2+。试验采用盆栽法,双因素随机区组设计,NaCl设置0,100,200,300 mmol·L-1 4个浓度梯度,营养液中的Ca2+由Ca(NO3)2提供,设置0,5,10,15,20 mmol·L-1 5个浓度梯度,共20个处理,每个处理3株珠美海棠幼苗,即3个重复。营养液的摄入采用浇灌法,对处理10 d后的珠美海棠幼苗进行光合指标的测定。
1.3 光合指标测定方法
采用CIRAS-2便携式全自动光合仪对珠美海棠叶片的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等进行测定。按不同处理分别随机选取叶片,用95%乙醇提取的方法进行叶绿体色素测定。
1.4 数据处理
采用Microsoft Excel 2007和SPSS 17.0统计软件对试验数据进行处理和统计分析。
2 结果与分析
2.1 外施Ca2+对盐胁迫下珠美海棠组培苗叶片色素含量的影响
由图1可知,在未施加Ca2+处理,珠美海棠叶绿素a的含量随着NaCl浓度的升高呈先升后降的趋势,其中NaCl 200 mmol·L-1时达到最大值,与NaCl 100 mmol·L-1和300 mmol·L-1处理差异不显著,三者均显著高于对照组;在未经NaCl胁迫处理,珠美海棠叶绿素a的含量随着Ca2+浓度的增加呈先上升后下降的趋势,在Ca2+浓度15 mmol·L-1时含量最高,可达到3.192 mg·g-1,与Ca2+浓度5,10,20 mmol·L-1差异不显著,均显著高于未施加Ca2+处理;在NaCl 100 mmol·L-1和200 mmol·L-1处理,叶绿素a含量在不同Ca2+浓度处理间差异均不显著;在NaCl浓度为300 mmol·L-1时(除去Ca2+缺失的处理),叶绿素a的含量随着Ca2+浓度的增加呈先升高再降低的趋势,至15 mmol·L-1时达到最高,显著高于其他Ca2+浓度处理,略高于未施加Ca2+处理,但差异不显著。
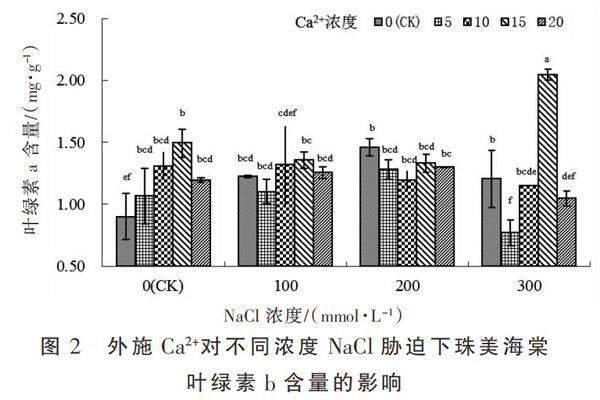
由图2可知,珠美海棠叶绿素b含量变化趋势与叶绿素a基本一致,在未施加Ca2+处理,珠美海棠叶绿素b含量随NaCl浓度升高呈先升后降的趋势,其中NaCl 200 mmol·L-1时达到最大值,与NaCl 100 mmol·L-1和300 mmol·L-1处理差异不显著,三者均显著高于对照组;在未经NaCl胁迫处理,珠美海棠叶绿素b含量随着Ca2+浓度增加亦呈先上升后下降的趋势,在Ca2+浓度15 mmol·L-1时含量最高值1.494 mg·g-1,且未施加Ca2+处理差异显著;在NaCl 浓度为100 mmol·L-1和200 mmol·L-1处理,叶绿素b含量在不同Ca2+浓度处理间差异均不显著;在NaCl浓度为300 mmol·L-1时(除去Ca2+缺失的处理),叶绿素b含量随着Ca2+浓度的增加呈先升高再降低的趋势,至15 mmol·L-1时达到最高,显著高于其他Ca2+浓度处理,是未施加Ca2+处理的1.7倍。
由图3可知,珠美海棠类胡萝卜素含量变化趋势与叶绿素a和b基本一致,在未施加Ca2+处理,珠美海棠类胡萝卜素含量随着NaCl浓度升高呈先升后降的趋势,其中NaCl 200 mmol·L-1时达到最大值,与NaCl 100 mmol·L-1和300 mmol·L-1处理差异不显著,三者均显著高于对照组;在未经NaCl胁迫处理,类胡萝卜素含量随Ca2+浓度增加呈先上升后下降的趋势,在Ca2+浓度10 mmol·L-1时达到最大值,显著高于未施加Ca2+和20 mmol·L-1 Ca2+处理;在NaCl浓度为100和200 mmol·L-1时,类胡萝卜素含量在不同Ca2+浓度处理间差异均不显著;在NaCl浓度为300 mmol·L-1时,类胡萝卜素含量表现为15 mmol·L-1Ca2+浓度处理显著低于其他处理。
2.2 外施Ca2+对盐胁迫下珠美海棠光合特性的影响
由图4可知,在未施加Ca2+处理,珠美海棠净光合速率(Pn)在不同NaCl处理表现为:100 mmol·L-1顯著高于对照组,二者均显著高于200,300 mmol·L-1处理,300 mmol·L-1较200 mmol·L-1略有升高,但差异未达显著水平。在不同浓度NaCl处理下珠美海棠的Pn随着Ca2+浓度的增加(除去Ca2+缺失组)呈先上升后下降的变化趋势,未经盐胁迫处理下,不同浓度Ca2+处理间差异显著,在外施Ca2+浓度为15 mmol·L-1时达到最大值,为5.8 μmol·m-2·s-1,是Ca2+浓度为0 mmol·L-1时的2.23倍;在NaCl浓度为100 mmol·L-1处理,未施加Ca2+处理Pn显著高于各其他Ca2+浓度处理;在NaCl浓度为200 mmol·L-1处理,施加Ca2+浓度为5,10,15 mmol·L-1 Pn显著优于未施加Ca2+处理,三者之间差异不显著;在NaCl浓度为300 mmol·L-1处理下,Pn在Ca2+浓度10 mmol·L-1 时显著高于未施加Ca2+处理。
由图5可知,在未施加Ca2+处理,珠美海棠气孔导度(Gs)在不同NaCl处理变化趋势与Pn基本一致,但对照组、100,300 mmol·L-1处理之间差异不显著,三者均显著高于200 mmol·L-1;在未经NaCl胁迫处理,珠美海棠叶片的Gs随着Ca2+浓度的增加呈先上升后下降的趋势,当Ca2+浓度为15 mmol·L-1时,最高值达到145 mmol·m-2·s-1,显著高于其他处理组,说明适宜浓度Ca2+可提高植物气孔的开度;在NaCl 100 mmol·L-1 时,未施加Ca2+与施加Ca2+各处理之间Gs差异不显著;在NaCl 200 mmol·L-1 时,施加Ca2+各处理之间Gs差异不显著,但均显著高于未施加Ca2+各处理;在NaCl 300 mmol·L-1 时, Ca2+ 在10 mmol·L-1Gs显著高于其他浓度处理和未施加Ca2+处理。珠美海棠蒸腾速率(Tr)的变化趋势与Gs基本一致(图6)。
由图7可知,在未施加Ca2+处理,珠美海棠胞间CO2浓度(Ci)随着NaCl浓度增加呈增加趋势,其中对照组与100 mmol·L-1处理之间差异不显著,二者显著低于200 mmol·L-1,后者又显著低于300 mmol·L-1处理;在未经NaCl胁迫处理,珠美海棠叶片的Ci在施加Ca2+处理均显著高于未施加Ca2+处理,除去Ca2+缺失组Ci随着Ca2+浓度的增加呈先降低后升高的趋势;在Ca2+浓度为15 mmol·L-1时,Ci值最低,结合Pn和Gs在该处理具有最高值,说明适宜浓度的Ca2+可通过提高珠美海棠对CO2的利用促进光合作用;在NaCl处理浓度为100 mmol·L-1时,仅10 mmol·L-1 Ca2+处理Ci与未施加Ca2+处理差异不显著,其他浓度均显著高于未施加Ca2+处理;在NaCl浓度200 mmol·L-1时,外施10和15 mmol·L-1的Ca2+处理Ci显著低于未施加Ca2+和Ca2+为20 mmol·L-1的处理;在NaCl为300 mmol·L-1时,Ca2+浓度为10 mmol·L-1处理显著低于其他Ca2+浓度和未施加Ca2+处理。
3 结论与讨论
光合作用是植物有机物合成的主要途径,光合能力的大小与植物对有机物的积累量直接相关。叶绿体是植物进行光合作用的主要场所,而叶绿素则是参与光合作用中光能吸收、传递和转化的重要色素,叶绿素含量与光合作用存在十分密切的关系,其含量在一定程度上可以反映出植物同化物质的能力大小,属于反应植物光合作用强度大小的生理指标。盐胁迫下一方面使植物吸收不到足够的水分和矿质营养,造成营养不良,降低叶绿素含量[9];另一方面还会使叶绿素合成受到抑制,从而提高叶绿素酸的活性,以至加速了叶绿素的降解。本研究结果表明,单一NaCl胁迫和外施Ca2+处理均可使珠美海棠叶绿素a和b含量呈先升高后下降的趋势,其中在NaCl和Ca2+浓度分别达到200 mmol·L-1和15 mmol·L-1时达到最高值,说明适宜浓度盐分可以在一定程度上提高珠美海棠叶绿素含量,但超过其阈值即表现为盐胁迫导致的下降。据报道,盐胁迫时外施一定浓度的Ca2+有助于保持水稻[10]、玉米[11]叶片较高的叶绿素含量。本研究结果中,在NaCl胁迫下施加Ca2+,仅在高盐(300 mmol·L-1)胁迫时可表现出15 mmol·L-1 Ca2+在叶绿素a和b含量上有一定的缓解作用。这可能是由于珠美海棠耐盐碱,仅在其阈值范围外的NaCl 300 mmol·L-1处理表现出盐胁迫伤害,此时Ca2+可通过改善细胞内外环境减轻盐胁迫中Na+对植物带来的毒害作用[12],但若Ca2+不足,会导致盐胁迫下植物对Ca2+的吸收和利用被抑制[13],故其仅在外施15 mmol·L-1 Ca2+处理中表现出Ca2+在叶绿素含量上的缓解作用,这种缓解作用可能與Ca2+在植物胁迫条件下发挥的作用有关,即一方面维持细胞壁和膜结合蛋白的稳定性及参与细胞内稳态(homeostasis)和生长发育的调节过程[14],另一方面可作为膜的保护物质,通过连接膜表面的磷酸盐和磷酸脂及蛋白质的羟基桥,维持盐胁迫下细胞质膜、叶绿体膜、液泡膜的结构稳定性和ATPare的活性[15]。
净光合速率直接反映了单位叶面积的同化能力[16-17],可以作为反映植物对盐胁迫的生理响应和鉴定植物抗盐能力的指标。一般认为,造成光合速率下降的因素主要有气孔限制因素和非气孔限制因素,其指标主要是气孔导度和胞间CO2浓度[18]。气孔部分关闭导致的气孔限制因素使Ci降低;而叶肉细胞光合性作用下降导致的非气孔限制因素使Ci增高。若两个因素同时存在,则占优势的因素成为Ci变化方式的主要原因。本研究结果表明,单一NaCl胁迫可使珠美海棠Pn、Gs和Tr呈先升高后下降的趋势,Ci趋势与三者相反,其中在NaCl浓度分别达到100 mmol·L-1时Pn、Gs和Tr达到最高值而Ci达到最低值,说明适宜浓度盐分可以在一定程度上提高珠美海棠的光合能力,表现为Gs略有增加而Ci略有降低,但Pn显著提高,但在超过其阈值的NaCl 200,300 mmol·L-1处理表现为盐胁迫导致的抑制作用,即Gs下降而Ci增加,导致Pn降低,此阶段(盐处理10 d)的状态应该示非气孔限制因素占主导。
薛延丰等[6]在营养液中施加Ca2+能起到降低盐胁迫所导致的氧化损伤,保持较高的酶活性,保护光系统Ⅱ并维持较高的光合效率;张宗申等[19]研究表明,盐胁迫下补充适宜浓度的Ca2+可以改善植物幼苗的抗逆性。在本试验中,仅在NaCl 200,300 mmol·L-1胁迫时可表现出外施Ca2+的优越性,即可在一定程度上通过提高Gs降低Ci,提高Pn,进而缓解高NaCl对珠美海棠的胁迫,其中NaCl 200,300 mmol·L-1处理分别以外施15, 10 mmol·L-1 Ca2+效果最佳。其可能原因与其对叶绿素含量的影响相同,即珠美海棠耐盐碱,仅在其阈值范围(NaCl 100 mmol·L-1)外表现出盐胁迫伤害,而仅在此时外施适宜浓度Ca2+改善植物抗盐性的作用才得以显现。另外,结合叶绿素含量结果发现,本试验中珠美海棠在NaCl 200 mmol·L-1时即出现了光合效率显著下降,而叶绿素含量仅在NaCl 300 mmol·L-1处理表现为下降但不显著,说明判断珠美海棠NaCl胁迫效应时,光合效率相关指标较叶绿素指标更为敏感。
由本试验研究可知,盐渍土面积不断扩大、耕地面积日益减少的今天,在引种经济盐生植物时,根据土壤盐碱性质和植物耐盐状况,适量增施钙肥,可起到提高植物抗性、促进植物生长的作用,可为珠美海棠在滨海盐碱地的引种栽培提供参考和借鉴。
参考文献:
[1]KOVDA V A. Loss of productive land due to salinazation[J]. Ambin XII, 1983, 2: 9-93.
[2]PARIDA A K, DAS A B. Salt tolerance and salinity effects on plants[J]. Ecotoxicol environ safe, 2005, 60: 324-349.
[3]MUNNS R. Physiological and hypotheses[J]. Plant cell environ, 1993, 16: 15-24.
[4]Bush D S. Calcium regulation in plant cells and its role in signaling[J]. Annual review of plant physiology and molecular biology, 1995, 46: 95-112.
[5]余叔文, 汤章城. 植物生理与分子生物学[M].2版.北京: 科学出版社, 1998: 123-133.
[6]薛延丰,刘兆普.钙离子对盐胁迫下菊芋幼苗的生长、生理反应和光合能力的影响[J]. 农业工程学报, 2006(9): 44-47.
[7]佟友丽, 冯君伟, 李玉花. 植物抗盐胁迫研究进展[J]. 生物技术通讯, 2008, 19(1): 138- 140.
[8]王玉珍, 冯学赞, 罗景兰. 多用途耐盐碱经济树种——珠美海棠[J]. 林业科技, 1999, 24(2): 53-54.
[9]刘国花. 植物抗盐机理研究进展[J].安徽农业科学, 2006, 34(23): 6111-6112.
[10]朱小军. 钙对盐胁迫下水稻幼苗盐害缓解的效应及机理研究[D]. 南京: 南京农业大学, 2004: 16-17.
[11]王玉鳳.玉米苗期对NaCl胁迫的响应与耐盐性调控机理的研究[D].沈阳: 沈阳农业大学, 2008.
[12]SHABALA S, DEMIDCHIK V. Extracellular Ca2+ ameliorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+- permeable channels[J]. Plant physiology, 2006(8): 1653-1665.
[13]EPSTEIN E, RAINS D W. Advances in salt tolerance[J]. Plant soil, 1987, 99: 17-29.
[14]BUSH D S. Calcium regulation in plant cells and its role in signalling[J]. Annual review of plant physiology and plant molecular biology, 1995, 46: 95-112.
[15]刘学平, 陶建敏, 高福新. 南京地区葡萄避雨栽培“H”形整形及根域限制栽培技术[J]. 中国南方果树, 2012, 41(6): 86-88.
[16]KAO W Y, TSAI T T, SHIH C N. Photosynthetic gas exchange and chlorophyll a, fluorescence of three wild soybean species in response to NaCl treatments[J]. Photosynthetica, 2003, 41(3): 415-419.
[17]YANG Y L, GUO J K, ZHANG F, et al. NaCl induced changes of the H+-ATPase in root plasma membrane of two wheat cultivars[J]. Plant science, 2004, 166(4): 913-918.
[18]FARQUHAR G D, VON CAEMMERER S, BERRY J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species[J]. Plant, 1980, 149 (1) : 78-90.
[19]张宗申, 利容千, 王建波. Ca2+预处理对热胁迫下辣椒叶肉细胞中Ca2+-ATP酶活性的影响[J]. 分子植物(英文版), 2001, 27(6): 451-454.
