高浓度乙烯利刺激诱导橡胶树死皮发生过程中的胶乳生理研究
2018-04-23冯成天郭秀丽胡义钰王真辉
何 晶,冯成天,郭秀丽,胡义钰,孙 亮,王真辉,袁 坤*
(1.海南大学 热带农林学院,海南 海口 570100;2.中国热带农业科学院 橡胶研究所,农业部橡胶树生物学与遗传资源利用重点实验室,海南 儋州 571737)
橡胶树是重要的热带经济作物,由其产生的天然橡胶是一种战略物资,在国民经济和国防建设中发挥着巨大作用。天然橡胶主要是来源于巴西橡胶树(HeveabrasiliensisMuell.Arg.)乳管细胞中的胶乳。橡胶树死皮(Tapping Panel Dryness,TPD,即割面干涸),是指乳管细胞丧失部分或全部产胶能力的现象,其症状为割线排胶减少甚至是完全停止排胶[1],严重影响胶乳产量[2-3],其发生的生理与分子机制是我国天然橡胶生产急需解决的重要问题。研究死皮发生动态变化过程中胶乳生理参数的变化,对橡胶树死皮发生机理的阐明具有一定的指导意义。
乙烯利广泛应用于天然橡胶割胶生产,具有增产的效果,但随着乙烯利的过度使用,其副作用也日益体现,主要表现为排胶线内缩、树皮肥肿和死皮等[4]。有研究认为,橡胶树死皮发生后,胶乳黄色体稳定性下降,黄色体破裂指数增加[5],死皮树中胶乳的硫醇含量明显下降,其作为自由基清除剂不能有效地清除有毒氧对黄色体膜的危害,所以导致黄色体膜受损,乳管停止排胶而逐渐死皮[6]。Putranto[7]等的研究表明,与正常树相比,死皮树胶乳中蔗糖、硫醇和无机磷含量均下降,暗示了死皮树乳管产胶能力下降。郭秀丽[8]等对橡胶树不同死皮程度植株的胶乳生理参数进行分析,结果显示,胶乳产量、硫醇和无机磷含量等随死皮程度的增加而降低,而蔗糖、镁离子含量及黄色体破裂指数等随死皮程度的增加而增加。
现有关于橡胶树死皮发生的生理基础的研究,主要是在胶园中直接选取健康树和死皮树进行比较分析,这种研究很难反映死皮发生的动态变化过程中生理指标的变化。本研究采用高浓度乙烯利刺激的方法,逐步诱导橡胶树死皮的发生:乙烯利刺激过程中,健康树割线症状由排胶线正常,逐渐转变为排胶线内缩(指割胶后排胶线比正常情况下明显变窄的现象,简称“内缩”)、严重内缩(指割胶后在割线上只能看到紧挨割面处有一条很窄的排胶线)[9],直至割线部分不排胶,即发生死皮。研究死皮发生动态变化过程中植株胶乳各生理参数的变化,从而为阐明橡胶树死皮发生机理提供一定的理论依据。
1 材料与方法
1.1 试验材料
试验材料来源于海南省儋州市西庆民营胶园,品系为橡胶树无性系热研7-33-97,2006年定植,2015年开割,采用s/2 d/3割制(割线长度为树围的1/2,每3 d割1刀)。试验前,该林段每年施用2次橡胶树专用复合肥,主要包括有机质、氮、磷、钾等。
1.2 试验方法
1.2.1 试验材料的选择与处理 选择树围大小相对一致、树皮完好、割面排胶正常的健康橡胶树作为试验材料。 2016年9-11月,用5%乙烯利浓度[10](一般生产上使用的乙烯利浓度为0.5%~1%,随着割龄增加,乙烯利浓度会适当增加,但一般不超过3%,而且涂乙烯利的周期一般是2次/月,涂施浓度太大或频率提高,都会对橡胶树造成伤害,导致死皮的发生)刺激所选取的健康树来诱导死皮的发生,即每10 d沿割线在割面2 cm宽处均匀涂刷乙烯利1次,共涂刷7次。健康树作为对照,健康树经5%乙烯利处理1个半月后产生割线内缩症状、2个月后产生严重内缩症状,2个半月后产生部分不排胶(死皮)症状。每种症状选取9株树,分成3组,每组3株树作为一个生物学重复,即每个处理3个生物学重复。取割胶后5~25 min排出的胶乳,每个生物学重复3株树各取10 mL胶乳等量混合,置于冰上后迅速带回实验室进行胶乳各生理参数的测定。
1.2.2 胶乳生理参数测定 胶乳产量、干胶含量及pH值的测定参照郭秀丽[8]等的方法进行;胶乳硫醇含量参考魏芳[11]等的DTNB试剂法进行测定;无机磷含量的测定采用钼酸铵比色法进行[12];蔗糖含量的测定采用蒽铜试剂法进行[13];粗酶液蛋白质含量及黄色体破裂指数分别参照袁坤[2]等、程成[14]等的方法进行测定。
1.3 数据统计分析
用Excel 2007、SAS 9.0以及Origin Pro 2015软件进行数据的处理、统计分析及作图。
2 结果与分析
2.1 乙烯利刺激诱导死皮发生过程中胶乳产量和干胶含量的变化
乙烯利刺激诱导橡胶树死皮发生的动态变化过程中,健康树割线症状由排胶线正常逐渐转变为内缩、严重内缩,直至部分死皮的发生。在这一动态变化过程中,平均单株胶乳产量呈现先上升后下降的趋势。未涂乙烯利之前,即健康树(对照)的胶乳产量为97.50 mL,涂高浓度乙烯利后割线出现内缩症状时,胶乳产量显著提高(P<0.05,下同),达最大值228.20 mL,是对照的2倍多。随着乙烯利刺激时间的增加,当割线出现严重内缩时,胶乳产量显著下降至86.60 mL。当出现部分死皮时,产量显著下降至最低值30.20 mL,约为对照健康树胶乳产量的31%(图1)。

注:图柱上方不同小写字母表示处理间差异显著(P<0.05)。下同。
图1乙烯利刺激诱导死皮发生过程中胶乳产量的变化
Fig.1 The change of latex yield during TPD occurrence induced by ethrel stimulation
干胶含量的变化趋势与胶乳产量的变化趋势基本一致,也呈先上升后下降的趋势。健康树的干胶含量为23.59 %,内缩症状时植株干胶含量显著上升至最大值39.09 %,随后干胶含量显著下降,发生部分死皮植株的干胶含量降至最低值22.11 %(图2)。
2.2 乙烯利刺激诱导死皮发生过程中胶乳各生理参数的变化
乙烯利刺激后,胶乳硫醇、无机磷及蔗糖含量均发生显著变化,同未刺激前植株相比,差异均达显著水平,且三者变化趋势一致,均表现出先上升后下降的趋势。乙烯利刺激前,胶乳硫醇、无机磷和蔗糖含量最低,分别为0.48、4.11 mmol/L和11.79 mmol/L;刺激后割线出现内缩症状时,胶乳硫醇、无机磷和蔗糖含量显著提高,达到最高值,分别为1.56、 32.76 mmol/L和58.10 mmol/L,约为对照的3倍多、8倍和4倍;随后胶乳硫醇、无机磷和蔗糖含量又显著下降,部分死皮时,三者分别为0.66、23.28 mmol/L和25.41 mmol/L。不同症状之间胶乳硫醇、无机磷和蔗糖含量均达显著性差异。
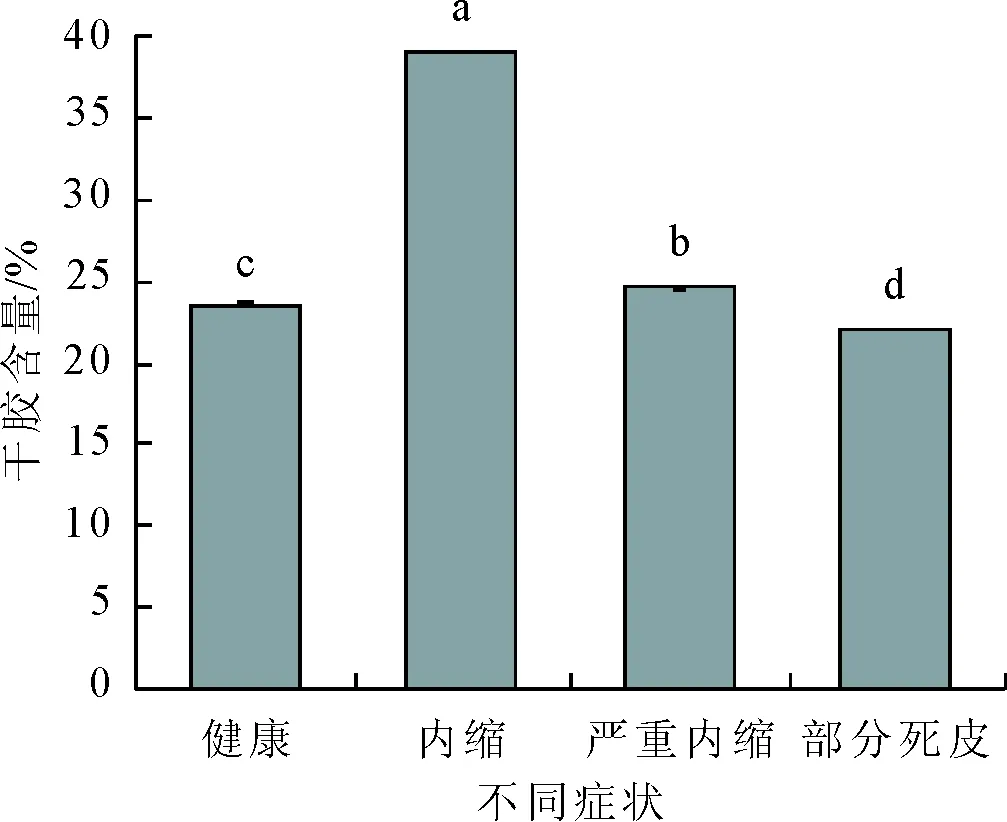
图2 乙烯利刺激诱导死皮发生过程中干胶含量的变化

图3 乙烯利刺激诱导死皮发生过程中硫醇含量的变化
乙烯利刺激前后胶乳pH值和粗酶液蛋白含量也均呈先上升后下降的趋势。严重内缩症状出现之前,健康及内缩植株间胶乳pH值无明显差异,内缩时胶乳pH值最高,为7.06。随后pH值下降,部分死皮出现后,胶乳pH值显著下降至最低值6.49;而粗酶液蛋白含量在植株割线症状由正常向严重内缩变化的过程中,呈显著上升趋势,严重内缩时上升至最大值15.85 mg/L,随后显著下降,部分死皮出现时降至最低值15.22 mg/L。
值得注意的是,乙烯利刺激后,黄色体破裂指数的变化趋势与上述各生理指标变化趋势不同,呈先下降后显著上升的趋势。健康植株黄色体破裂指数为18.72%,当割线出现内缩时,黄色体破裂指数显著下降,达最低值14.27%。随后又显著上升,出现严重内缩时,破裂指数上升至19.15%,当部分死皮出现后,破裂指数显著增加到最高值33.69%,约为健康植株的1.8倍。

图4 乙烯利刺激诱导死皮发生过程中无机磷含量的变化
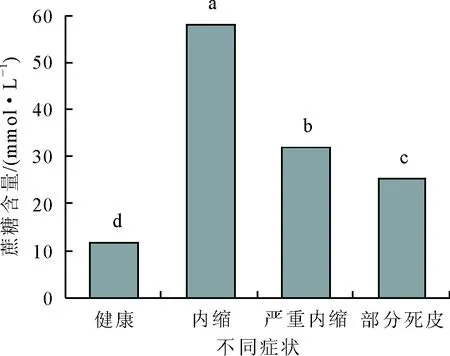
图5 乙烯利刺激诱导死皮发生过程中蔗糖含量的变化
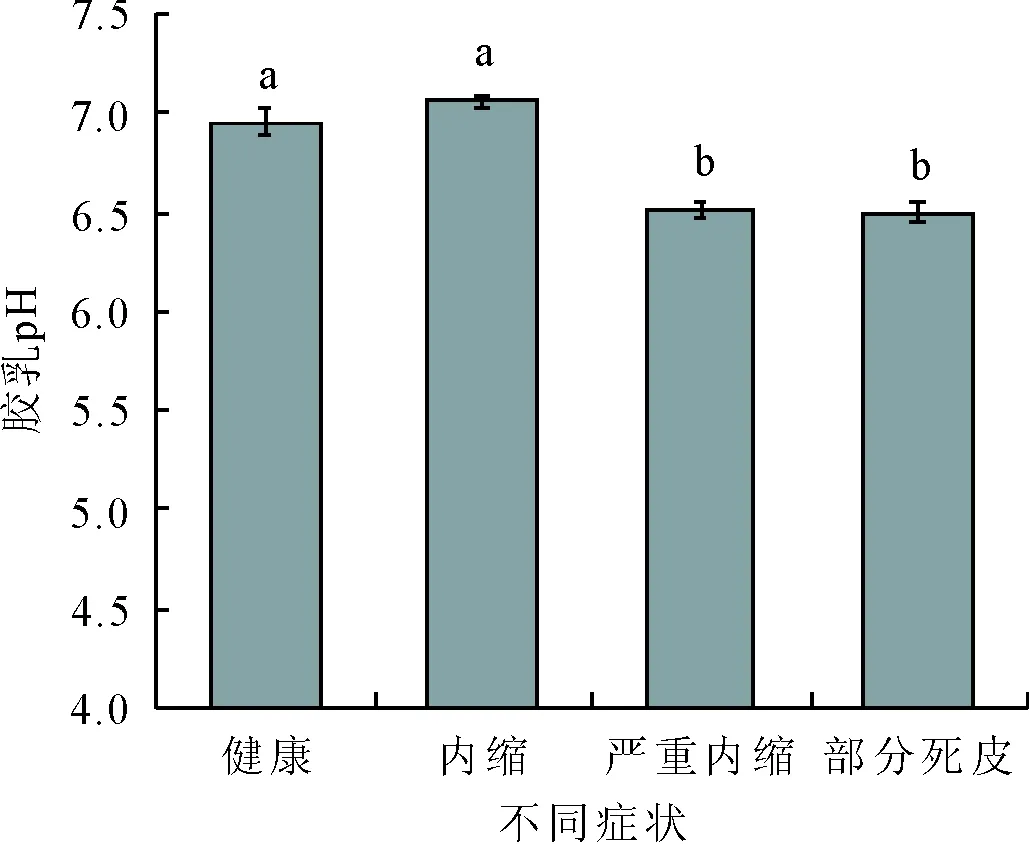
图6 乙烯利刺激诱导死皮发生过程中pH值的变化
3 结论与讨论
采用乙烯利刺激已成为提高橡胶树产量的重要手段,并在天然橡胶生产中大面积推广应用[15-16],涂施乙烯利可促使橡胶树胶乳产量增加1.5~2倍[17]。但是过度的乙烯利刺激会导致橡胶树死皮的发生。乙烯利过度刺激诱导橡胶树死皮的组织学、生物化学等方面前人已开展过一些研究[4,18-20],但关于生理方面的研究还未见报道。

图7 乙烯利刺激诱导死皮发生过程中粗酶液蛋白含量的变化
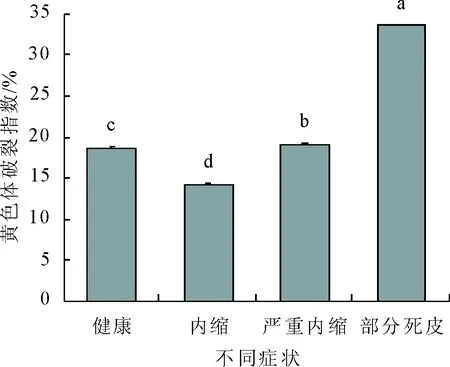
图8 乙烯利刺激诱导死皮发生过程中黄色体破裂指数的变化
本研究结果表明,高浓度乙烯利刺激后,割线由正常排胶逐渐发展为排胶线内缩、严重内缩,最后部分割线不排胶,即死皮发生。吴继林[4]等通过对过度乙烯利刺激后割线树皮结构变化的研究中发现,长时间过度刺激采胶导致失去排胶功能的乳管列从树皮外层逐渐扩展到树皮内层,直至死皮的发生,这与本研究的观测结果一致。
胶乳产量和干胶含量反映了胶乳的合成和再生能力,而黄色体破裂指数则反映了胶乳中黄色体的稳定性、完整性。破裂指数愈大,胶乳愈不稳定,愈容易凝固堵塞,使胶乳停排[5,14]。本研究结果显示,高浓度的乙烯利刺激诱导橡胶树死皮发生的过程中,胶乳产量和干胶含量均呈现先上升后下降的趋势。在刺激的早期出现割线内缩症状时,胶乳产量和干胶含量均达最大值,而此时黄色体破裂指数最低。表明刺激早期,乙烯利刺激可能通过提高黄色体的稳定性来延长排胶时间,进而增加胶乳产量,这与目前认为的乙烯利刺激增产的机制是一致的[15,21]。随着刺激时间的延长,胶乳产量和干胶含量明显下降,当出现部分死皮时,二者均降至最低值,而此时黄色体破裂指数达最大值,这说明乙烯利的过度刺激导致黄色体的稳定性受到破坏,黄色体破裂后释放出凝固因子,胶乳原位凝固致使排胶受阻而死皮发生[21]。吴继林[4]等的组织学研究结果也显示,过度刺激后树皮乳管内的黄色体破裂并与橡胶粒子一起形成絮状物,这进一步验证了本研究的结果。上述研究结果说明,黄色体在高浓度乙烯利刺激诱导橡胶树死皮发生过程中发挥重要作用。
本研究的结果可以看出,乙烯利刺激诱导死皮发生过程中,除黄色体破裂指数外,其余各生理参数的变化趋势均呈先上升后下降的趋势。硫醇是乳管细胞中重要的还原剂,具有清除乳管细胞代谢过程中所产生的有毒氧的功能,从而减少有毒氧对乳管细胞的伤害[11,22]。校现周和蔡磊[20]等的研究显示,高浓度的乙烯利刺激可提高乳管细胞中活性氧的产生速率和含量。据此推测,在本研究中高浓度的乙烯利刺激诱导橡胶树死皮发生过程中,活性氧含量可能也是逐渐增加的。在刺激早期出现内缩症状时,硫醇含量达最大值,暗示此时硫醇对活性氧的清除能力较强。随着刺激时间的延长,硫醇含量逐渐降低至出现部分死皮,说明随着活性氧的大量产生,硫醇对活性氧的清除能力逐渐下降,导致有毒氧的过多积累对乳管细胞造成伤害,进而可能导致死皮的发生。
无机磷是反映乳管能量代谢的重要指标。胶乳中无机磷含量高,说明乳管能量代谢活跃[23]。本研究结果显示,乙烯利刺激诱导死皮发生过程中,乳管系统代谢能力先增强后逐渐减弱。
蔗糖是橡胶生物合成的原料,其含量是综合反映乳管蔗糖供给和利用效率的指标[24]。本研究结果显示,在诱导死皮发生过程中,蔗糖含量先升高随后降低,说明乙烯利刺激后的早期刚出现内缩症状时,蔗糖含量显著上升,即用于合成橡胶的前体充足,胶乳的合成和再生能力较强,因此胶乳产量很高。随着严重内缩及部分死皮的出现,蔗糖含量逐渐下降,进而导致胶乳产量的降低。此外,胶乳pH值与胶乳再生相关[25],而粗酶液蛋白含量与乳管系统合成代谢相关[26],二者在乙烯利刺激诱导死皮发生过程中也发生显著变化,表明它们与橡胶树死皮发生也是紧密相关的。
橡胶树死皮发生是一个动态变化的过程。本研究利用高浓度乙烯利刺激诱导橡胶树死皮的发生,动态监测了死皮发生过程中胶乳各生理指标的变化,为后续进一步开展死皮发生动态变化过程中相关基因或蛋白的表达变化奠定了基础,也为阐明橡胶树死皮发生机理提供一定的理论依据。
参考文献:
[1] 袁坤,徐智娟,王真辉,等.橡胶树胶乳死皮相关蛋白的鉴定及分析 [J].西北林学院学报,2012,27(6):105-109.
YUAN K,XU Z J,WANG Z H,etal.Identification and analysis of latex proteins related with tapping panel dryness (TPD) inHeveabrasiliensis[J].Journal of Northwest Forestry University,2012,27(6):105-109.(in Chinese)
[2] 袁坤,周雪梅,李建辉,等.死皮防治剂对死皮橡胶树胶乳生理的影响 [J].湖北农业科学,2011,50(17):3570-3572.
YUAN K,ZHOU X M,LI J H,etal.The influence of medicament on latex physiology of rubber tree with tapping panel dryness [J].Hubei Agricultural Sciences,2011,50(17):3570-3572.(in Chinese)
[3] 袁坤,杨礼富,陈帮乾,等.海南植胶区橡胶树死皮发生现状分析 [J].西北林学院学报,2016,31(1):176-179.
YUAN K,YANG L F,CHEN B Q,etal.The investigation on the current situation ofHeveabrasiliensisTPD occurring in Hainan [J].Journal of Northwest Forestry University,2016,31(1):176-179.(in Chinese)
[4] 吴继林,谭海燕,郝秉中.乙烯利过度刺激采胶诱导巴西橡胶树割面干涸病的研究 [J].热带作物学报,2008,29(1):1-9.
WU J L,TAN H Y,HAO B J.Aspects of the development of tapping panel dryness induced by overstimulation with ethrel inHeveabrasiliensis[J].Chinese Journal of Tropical Crops,2008,29(1):1-9.(in Chinese)
[5] 杨少琼,熊涓涓.橡胶树乳管系统功能的胶乳诊断Ⅱ.黄色体破裂指数的测定 [J].热带作物研究,1989(1):68-71.
[6] 范思伟,杨少琼.强割和排胶过度引起的死皮是一种特殊的局部衰老病害 [J].热带作物学报,1995,16(2):15-22.
FAN S W,YANG S Q.A special and localized senescence disease-bark dryness due to over exploitation and over-drainage [J].Chinese Journal of Tropical Crops,1995,16(2):15-22.(in Chinese)
[7] PUTRANTO R A,HERLINAWATI E,RIO M,etal.Involvement of ethylene in the latex metabolism and tapping panel dryness ofHeveabrasiliensis[J].International Journal of Molecular Sciences,2015,16(8):17885-17908.
[8] 郭秀丽,孙亮,胡义钰,等.巴西橡胶树不同死皮程度植株的胶乳生理参数分析 [J].南方农业学报,2016,47(9):1553-1557.
GUO X L,SUN L,HU Y Y,etal.Analysis on latex physiological parameters ofHeveabrasiliensisplants at different tapping panel dryness extents [J].Journal of Southern Agriculture,2016,47(9):1553-1557.(in Chinese)
[9] 郝秉中,吴继林,云翠英.乙烯利刺激割胶对巴西橡胶树树皮结构的影响 [J].热带作物学报,1980,1(1):61-65.
HAO B Z,WU J L,YUN C Y.The effect of stimulation tapping with ethrel on the structure of the bark ofHeveabrasiliensisMuell.Arg.[J].Chinese Journal of Tropical Crops,1980,1(1):61-65.(in Chinese)
[10] 闫洁,陈守才,夏志辉.橡胶树死皮病胶乳C-乳清差异表达蛋白质的筛选与鉴定 [J].中国生物工程杂志,2008,28(6):28-36.
YAN J,CHEN S C,XIA Z H.Screening and identification of differential expressed proteins in C-serum on latex of tapping panel dryness (TPD) in rubber tree (HeveabrasiliensisMuell.Arg.) [J].China Biotechnology,2008,28 (6):28-36.(in Chinese)
[11] 魏芳,罗世巧,校现周.橡胶树胶乳中硫醇功能以及模式植物中硫醇合成途径研究进展 [J].热带农业科学,2012,32(8):12-17.
WEI F,LUO S Q,XIAO X Z.Function of thiols in rubber tree latex and recent researches on thiols anabolism pathway in model plant [J].Chinese Journal of Tropical Agriculture,2012,32(8):12-17.(in Chinese)
[12] TAUSSKY H H,SHORR E A.Microcolorimetric method for the determination of inorganic phosphorus [J].Journal of Biological Chemistry,1953,202:675-685.
[13] ASHWELL G.Colorimetric analysis of sugars [J].Methods in Enzymology,1957,3:73-105.
[14] 程成,史敏晶,田维敏.巴西橡胶树胶乳中黄色体破裂指数测定方法的优化 [J].热带作物学报,2012,33(7):1197-1203.
CHENG C,SHI M J,TIAN W M.An improved method for measuring the lutoid bursting index inHeveabrasiliensis[J].Chinese Journal of Tropical Crops,2012,33(7):1197-1203.(in Chinese)
[15] 庄海燕,安锋,张硕新,等.乙烯利刺激橡胶树增产机制研究进展 [J].林业科学,2010,46(7):120-125.
ZHUANG H Y,AN F,ZHANG S X,etal.Progress in study on the mechanisms to increase latex yield ofHeveabrasiliensisby ethephon stimula [J].Scientia Silvae Sinicae,2010,46(7):120-125.(in Chinese)
[16] 位明明,李维国,高新生,等.巴西橡胶树响应乙烯利刺激的生理及其分子调控机制研究进展 [J].生物技术通报,2016,32(3):1-11.
WEI M M,LI W G,GAO X S,etal.Research progress of the physiological and molecular regulation mechanism ofHeveabrasiliensisin response to ethephon stimulation [J].Biotechnology Bulletin,2016,32(3):1-11.(in Chinese)
[17] PUJADE-RENAUD V,CLEMENT A,PERROT-RECBENMANN C,etal.Ethylene induced increase in glutamine synthetase activity and mRNA levels inHeveabrasiliensislatex cells [J].Plant Physiology,1994,105(1):127-132.
[18] HAO B Z,WU J L.Ultrastructure of laticifers in drying bark induced by overexploitation of with ethephon [J].Journal of Natural Rubber Research,1993,8:286-292.
[19] CHRESTIN H.Biochemical aspects of bark dryness induced by overstimulation of rubber trees with ethrel [C].In:d’Auzac J,Jacob J L,Chrestin Heds.Physiology of Rubber Tree Latex.Boca:CRC Press,1989,431-441.
[20] 校现周,蔡磊.乙烯利刺激对橡胶树乳管细胞活性氧代谢的影响 [J].热带作物学报,2003,24(1):1-7.
XIAO X Z,CAI L.Effect of ethrel stimulation on metabolism of active oxygen in laticiferous cell [J].Chinese Journal of Tropical Crops,2003,24(1):1-7.(in Chinese)
[21] 朱家红,张全琪,张治礼.乙烯利刺激橡胶树增产及其分子生物学基础 [J].植物生理学通讯,2010,46(1):87-93.
ZHU J H,ZHANG Q Q,ZHANG Z L.Ethephon stimulation on latex production and its molecular biological basis inHeveabrasiliensis[J].Plant Physiology Communications,2010,46(1):87-93.(in Chinese)
[22] 校现周.橡胶胶乳中R-SH的生理作用 [J].热带作物研究,1996(3):5-9.
[23] 谭德冠,姚庆收,张伟算,等.10个橡胶树新品系幼龄试割期间生理参数的分析与比较 [J].热带农业科学,2004,24(1):1-6.
TAN D G,YAO Q S,ZHANG W S,etal.Comparison and evaluation of latex physiology of ten new rubber clones during the test tapping period [J].Chinese Journal of Tropical Agriculture,2004,24(1):1-6.(in Chinese)
[24] 黄德宝,秦云霞,唐朝荣.橡胶树三个品系(热研8-79、热研7-33-97和PR107)胶乳生理参数的比较研究 [J].热带亚热带植物学报,2010,18(2):170-175.
HUANG D B,QIN Y X,TANG C R.Physiological characters of latex from three hevea clones (Reyan 8-79,Reyan 7-33-97 and PR 107) [J].Journal of Tropical and Subtropical Botany,2010,18(2):170-175.(in Chinese)
[25] 王岳坤,阳江华,秦云霞.PR107两种割胶制度胶乳生理参数的季节变化 [J].热带作物学报,2014,35(3):419-424.
WANG Y K,YANG J H,QIN Y X.Seasonal variation of latex physiological parameters in hevea clone PR107 exploited by two tapping system [J].Chinese Journal of Tropical Crops,2014,35(3):419-424.(in Chinese)
[26] 敖硕昌,赵淑娟,何长贵.橡胶树高产生理基础研究Ⅰ:胶乳生理和排胶特性的研究 [J].云南热作科技,1994,17(3):6-10.
