妊娠后期饲粮中添加乳酸菌与酵母菌的复合菌对母猪繁殖性能、血浆脂质代谢和抗氧化能力的影响
2018-04-20任红立汪晶晶金三俊董佳琦刁新平
任红立 汪晶晶 金三俊 董佳琦 刁新平
(东北农业大学动物科学技术学院,哈尔滨 150030)
益生菌替代抗生素已成为养猪业的一大研究热点,益生菌的作用广泛,在猪生产中有良好的应用效果。乳酸菌是一种能够利用碳源产生乳酸的细菌[1],具有增强动物机体免疫力、改善胃肠道菌群平衡、抗氧化、抗肿瘤、抗高血压和降低胆固醇含量等作用[2]。酵母菌是一种应用较广泛的益生菌,具有增强动物机体免疫力,改善胃肠道菌群平衡,调节机体代谢,提供营养源等作用[3]。乳酸菌和酵母菌是对母猪有益的2种益生菌,可以在同一体系中共存,研究发现乳酸菌的代谢产物可为酵母菌提供碳源[4],酵母菌发酵产生的营养因子可被乳酸菌利用[5],二者的代谢产物可以互相被利用[6]。益生菌能提高母猪的繁殖性能,增强免疫力,提高后代仔猪的生长性能。李秋艳等[7]研究发现,饲粮中添加乳酸菌、酵母菌和枯草芽孢杆菌复合菌可提高瘦肉型母猪的繁殖性能。闫俊书等[8]研究发现,饲粮中添加植酸酶、复合酶、枯草芽孢杆菌、酵母菌、乳酸菌复合制剂可提高发酵床育肥后期猪的生产性能。妊娠后期是胎儿生长发育的最快时期,胎儿体重增长快,母猪的营养水平直接影响胎儿的生长、仔猪的初生窝重和个体重以及哺乳期的哺乳情况。妊娠后期在母猪饲粮中添加益生菌可改善母猪胃肠道菌群平衡,提高饲料利用率,增强母猪免疫力,减少母猪患病率,但因试验因素不同,研究结果有所差异。本试验旨在研究妊娠后期饲粮中添加不同剂量的乳酸菌与酵母菌的复合菌对母猪繁殖性能、血浆脂质代谢和抗氧化能力的影响,确定复合菌的最佳添加剂量,为益生菌在母猪生产中的合理应用提供科学依据。
1 材料与方法
1.1 试验材料
复合菌制剂:乳酸菌、酵母菌及其共用复合培养基,均购于深圳市百澳飞生物公司,产品为奶黄色粉末状,酸奶香味,用发酵罐发酵24 h后,以生物发酵液的形式饲喂母猪。发酵液的pH为3.59,含乳酸菌1.0×108CFU/mL,酵母菌8×107CFU/mL。
1.2 试验设计
饲养试验于2017年2—4月份在河南省鹿邑县天种猪场进行。选取体况相近、妊娠第80天的3胎“大×长”杂交母猪48头,随机分为4组,每组12个重复,每个重复1头母猪。对照组饲喂基础饲粮,Ⅰ组、Ⅱ组和Ⅲ组饲粮分别在基础饲粮中添加150、300和450 mL/d的乳酸菌与酵母菌的复合菌发酵液。预试期5 d,正试期37 d。正式试验从妊娠第85天开始,产后第7天结束,其中产后7 d内饲粮中不添加复合菌发酵液。基础饲粮参照NRC(2012)母猪营养需要配制,其组成及营养水平见表1。
1.3 饲养管理
妊娠母猪按照猪场常规程序管理,妊娠后期母猪大圈饲养,预产期前3 d转舍到母猪产房,进行单栏饲养。对照组和试验组妊娠后期母猪在同一妊娠舍、同一产房进行饲养,妊娠后期和哺乳期前7 d均为限制饲喂,妊娠后期喂料量为1.8 kg/(头·次),每天饲喂2次,分娩当天不饲喂,分娩后第1天0.5 kg/(头·d),随后每天增加0.5 kg,7 d后自由采食。各组母猪饲粮在基础饲粮中加入适量水,以湿拌料的方式饲喂,Ⅰ组、Ⅱ组和Ⅲ组饲粮中添加不同剂量的复合菌发酵液。充足饮水,环境相同,按照猪场常规免疫程序进行免疫接种。

表1 基础饲粮组成及营养水平(风干基础)
1)预混料为每千克饲粮提供 The premix provided the following per kg of diets:VA 25 000 IU,VB11 mg,VB28 mg,VB63 mg,VB120.015 mg,VD35 000 IU,VK 2.5 mg,叶酸 folic acid 0.25 mg,D-泛酸D-pantothenic acid 12.5 mg,烟酸 nicotinic acid 17.5 mg,Cu (as copper sulfate) 30 mg,Fe (as ferrous sulfate) 100 mg,Mn (as manganese sulfate) 40 mg,Zn (as zinc sulfate) 100 mg,I (as potassium iodide) 0.25 mg,Se (as sodium selenite) 0.25 mg,Cr (chromium sulfate)。
2)计算值 Calculated value。
1.4 检测指标
1.4.1 繁殖性能
记录每头试验母猪的繁殖性能指标,包括总产仔数、活仔数、木乃伊仔数、死胎数以及仔猪初生重。计算出母猪的初生活仔率、木乃伊率、死胎率及仔猪初生窝重。
1.4.2 血浆脂质代谢和抗氧化指标
于母猪妊娠第85天(空腹)和分娩当天,采用肝素钠抗凝真空采血管进行耳缘静脉采血,每头母猪采集10 mL新鲜血液,静置30 min后,离心机3 000 r/min离心20 min,取上层血浆保存于-20 ℃冰箱中。
血浆脂质代谢指标包括总胆固醇(TC)、甘油三酯(TG)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)含量,采用全自动生化仪进行测定。血浆抗氧化指标包括丙二醛(MDA)含量和超氧化物歧化酶(SOD)活性,采用比色法测定,试剂盒购自北京华英生物技术研究所。
1.5 数据处理
试验数据用Excel 2007软件进行初步处理后,采用SPSS 19.0进行单因素方差分析(one-way ANOVA),差异显著者进行Duncan氏多重比较法检验,结果以“平均值±标准差”表示,P<0.05表示差异显著。
2 结 果
2.1 乳酸菌与酵母菌的复合菌对母猪繁殖性能的影响
由表2可知,各组母猪的总产仔数、活仔数、木乃伊仔数、死胎数、初生活仔率、木乃伊率和死胎率均无显著差异(P>0.05)。与对照组相比,Ⅱ组的仔猪初生窝重显著增加(P<0.05),Ⅰ组和Ⅲ组的仔猪初生窝重无显著差异(P>0.05);Ⅰ组、Ⅱ组和Ⅲ组的仔猪初生重显著增加(P<0.05)。

表2 饲粮添加乳酸菌与酵母菌的复合菌对母猪繁殖性能的影响
同行数据肩标相同小写字母或无字母表示差异不显著(P>0.05),不同小写字母表示差异显著(P<0.05)。下表同。
In the same row, values with the same small or no letter superscripts mean no significant difference (P>0.05), while with different small letter superscripts mean significant difference (P<0.05). The same as below.
2.2 乳酸菌与酵母菌的复合菌对母猪血浆脂质代谢指标的影响
由表3可知,各组妊娠第85天母猪的血浆TC、TG、HDL-C和LDL-C含量均无显著差异(P>0.05)。由表4可知,各组分娩母猪的血浆LDL-C含量无显著差异(P>0.05);与对照组相比,Ⅰ组和Ⅱ组的血浆TC含量显著增加(P<0.05),Ⅲ组无显著差异(P>0.05);Ⅱ组的血浆TG含量显著增加(P<0.05),Ⅰ组和Ⅲ组无显著差异(P>0.05);Ⅰ组和Ⅱ组的血浆HDL-C含量显著增加(P<0.05),Ⅲ组无显著差异(P>0.05)。
2.3 乳酸菌与酵母菌的复合菌对母猪血浆抗氧化指标的影响
由表5可知,各组妊娠第85天母猪的血浆MDA含量和SOD活性均无显著差异(P>0.05)。与对照组相比,Ⅰ组和Ⅲ组分娩母猪的血浆MDA含量显著降低(P<0.05),Ⅱ组无显著差异(P>0.05);各组分娩母猪的血浆SOD活性无显著差异(P>0.05)。
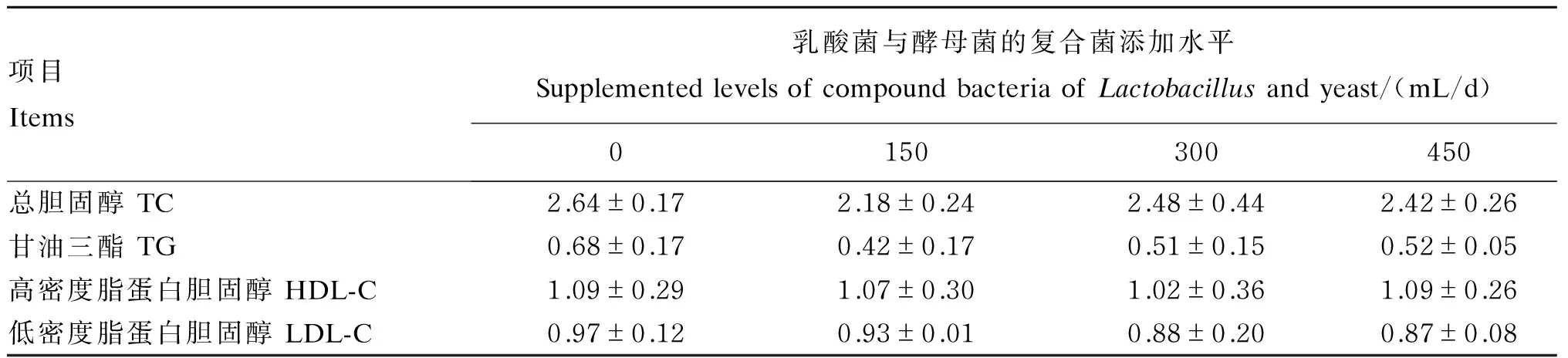
表3 饲粮添加乳酸菌与酵母菌的复合菌对妊娠第85天母猪血浆脂质代谢指标的影响

表4 饲粮添加乳酸菌与酵母菌的复合菌对分娩母猪血浆脂质代谢指标的影响
3 讨 论
3.1 乳酸菌与酵母菌的复合菌对母猪繁殖性能的影响
妊娠后期母猪的繁殖性能主要表现在窝产活仔数、死胎数、初生仔猪窝重、初生仔猪个体重、初生活仔率和死胎率。母猪的窝产仔数与母猪的品种、妊娠前期的营养水平相关[9]。母猪的营养和健康直接影响其繁殖性能和后代仔猪的生长发育[10]。妊娠后期母猪饲粮添加益生菌能增强母猪体质,提高母猪的饲料利用率,进而提高母猪的繁殖性能。但益生菌对妊娠母猪繁殖性能影响的研究结果有所差异,这可能是由于母猪的品种、妊娠时期、饲养环境、饲养管理、营养水平以及益生菌的种类、数量、比例的不同所导致。刘晴等[11]研究发现,分娩前30 d在母猪饲粮中添加芽孢杆菌、乳酸菌和酵母菌的复合益生菌能显著提高初生仔猪个体重,但产仔数无显著差异。霍军等[12]研究饲粮中添加微生态制剂对繁殖母猪生产性能的影响时发现,分娩前45 d在母猪饲粮中添加微生态制剂能显著提高窝产仔数、窝产活仔数和仔猪初生重,显著降低死胎率。付云超等[13]研究复合菌制剂对母猪繁殖机能的影响时发现,分娩前45 d在纯种长白经产母猪饲粮中添加复合菌制剂能显著提高仔猪初生窝重和仔猪初生重,但对初生仔猪数、初生活仔数和初生仔猪成活率无显著影响。本试验研究发现,妊娠后期母猪饲粮中添加乳酸菌与酵母菌的复合菌能显著提高初生仔猪窝重和初生仔猪重,这可能是由于复合菌制剂增强了母猪对饲粮中营养物质的吸收和消化利用,优化了肠道环境。本试验研究也发现,复合菌对妊娠后期母猪的窝产仔数、活仔数、木乃伊仔数、死胎数、初生活仔率、木乃伊率、死胎率等繁殖性能指标无显著影响,这与张国权等[14]和龙广[15]的研究结果相一致。
3.2 乳酸菌与酵母菌的复合菌对母猪血浆脂质代谢的影响
TC是动物体必需的营养成分[16],是细胞构成的重要成分,是性激素等类固醇激素、维生素D3和胆酸合成的原料[17];可维持白细胞的生理功能和血管壁的完整性,降低癌变率;可通过启动T细胞生成免疫分子白细胞介素 2(IL-2),诱导T淋巴细胞增殖发挥免疫应答,促进B淋巴细胞的免疫应答[18]。TC在血液中常通过高密度脂蛋白(HDL)和低密度脂蛋白(LDL)进行转运[19]。LDL-C含量超标,说明存在于血液循环中的TC过多,血脂含量过高,研究显示LDL-C含量的增加是动脉硬化的危险信号[20]。HDL-C可清除血管壁多余的LDL-C,对血管有保护作用,降低动脉硬化的发生率[21]。妊娠母体的血脂含量在一定范围内升高对母体和胎儿是有益的[22]。妊娠后期母体的脂肪代谢增强,母体血浆中游离脂肪酸和甘油含量增加,可以作为母体合成葡萄糖的优先有效物质,作为能量储存;当妊娠后期母体营养缺乏时,保证能向胎儿不断地输入所需营养物质,同时母体可利用自身的脂肪代谢产物,维持自身的代谢平衡[23]。但血脂含量过高,脂代谢异常,会导致胎盘早剥及妊娠高血压综合征的发生[22]。益生菌具有降血脂的作用[24],研究发现乳酸菌可以降低血液中TC含量。有关乳酸菌对TC的降解机制尚不明确,还在持续研究中,主要有以下几种理论:1)吸收理论,乳酸菌对TC的降解是通过菌体吸收介质中的TC,不同胆盐浓度使得菌体对TC的吸收速度不同[25-27];2)沉淀理论,研究发现乳酸菌产生的胆盐水解酶是降低TC含量的关键性因素,它能改变胆盐结构,使胆盐与TC形成沉淀,排出体外,降低体内TC含量[28-29];3)掺入细胞膜理论,研究发现菌体吸收TC,但是吸收进菌体的TC并没有完全进入细胞内,有一部分进入了细胞膜,在细胞膜表面,还有一些其他的降解TC的机制理论[30-31]。本试验研究结果表明,与对照组相比,Ⅱ组分娩母猪的血浆TC、TG和HDL-C含量显著增加,但各组血浆LDL-C含量无显著差异。说明复合菌不能降低分娩母猪的血脂含量,但能促进血脂的代谢,保护动脉血管的完整性。研究显示,妊娠妇女血清TG含量越高,胎儿出生时体重增加越明显;也有研究显示胎儿初生重与母体血脂含量无相关性,目前没有统一的结论[22]。本试验发现,初生仔猪窝重为Ⅱ组>Ⅰ组>Ⅲ组>对照组,分娩母猪的血浆TG含量也为Ⅱ组>Ⅰ组>Ⅲ组>对照组,与血清TG含量越高胎儿体重越大结果相一致。
3.3 乳酸菌与酵母菌的复合菌对母猪血浆抗氧化能力的影响
氧化应激是机体内氧化物的生成能力超过抗氧化防御系统清理的能力或抗氧化防御系统能力减弱时发生的抗氧化防御系统和氧化物生成的不平衡状态,也是活性氧的生成和降解的不平衡状态[32-33]。适量的活性氧含量是机体进行正常的生理活动所必需的,但过量的活性氧则会损伤机体的正常生理机能。能反映机体氧化损伤和抗氧化损伤的主要指标分别是MDA和SOD[34]。MDA是血浆中脂质与自由基结合的过氧化脂质的降解产物,可作为间接反映机体内自由基含量的指标,血浆MDA含量降低说明自由基含量降低,体内抗氧化系统的防御能力升高,抗氧化能力增强。SOD是清除动物体内自由基的主要内源性抗氧化酶之一,SOD活性升高,说明抗氧化防御系统能力升高,抗氧化能力增强;反之,说明抗氧化能力减弱[34]。研究证明乳酸菌具有抗氧化活性[35],乳酸菌的抗氧化物质主要包括抗氧化酶、锰离子和巯基化合物等[36],但乳酸菌抗氧化的具体机制还不十分明确,可能有以下几种:1)抗氧化酶机制,乳酸菌内主要的抗氧化酶有SOD、过氧化氢酶(H2O2)、烟酰胺腺嘌呤二核苷酸(NADH)氧化酶/NADH过氧化物酶等;2)锰离子抗氧化机制,研究发现乳酸菌中的高浓度锰离子抗氧化作用类似于SOD的抗氧化作用[37];3)巯基化合物抗氧化机制,乳酸菌具有蛋白巯基和非蛋白巯基化合物,它们能够清除羟自由基,起到抗氧化的作用[36];4)适应性机制,乳酸菌经过一定的环境胁迫,会增强一些胁迫应激反应的防御能力,如在一定环境下,产生的大量氧自由基就会诱导乳酸菌的抗氧化反应[37]。本试验研究发现,妊娠后期母猪饲粮添加乳酸菌与酵母菌的复合菌对分娩母猪的血浆SOD活性无显著影响,但Ⅰ组和Ⅲ组分娩母猪的血浆MDA含量显著低于对照组。说明复合菌能降低自由基的含量,减少了氧化物的生成,增强了抗氧化防御系统的能力。
4 结 论
妊娠后期母猪饲粮添加乳酸菌与酵母菌的复合菌能改善母猪的繁殖性能、血浆脂质代谢和抗氧化能力,其中对仔猪初生窝重和仔猪初生重、分娩母猪的血浆TC、TG、HLD-C和MDA含量有显著影响。在本试验条件下,从繁殖性能和血浆脂质代谢来看,复合菌发酵液的最适添加量为300 mL/d;从抗氧化能力和经济效益来看,复合菌发酵液的最适添加量为150 mL/d。
参考文献:
[1] HALASZ A.Food quality and standards-vol.Ⅲ-Latic acid bacteria[M].Budapest:EOLSS,2009:70-83.
[2] 湛剑龙,胡萍,陈韵,等.功能性乳酸菌的应用研究[J].食品安全质量检测学报,2014,5(4):1002-1009.
[3] 李磊. 布拉氏酵母菌抗腹泻研究[D].硕士学位论文.济南:齐鲁工业大学, 2013.
[4] 闫彬,贺银凤.乳酸菌与酵母菌共生机理综述[J].食品科学,2012,33(3):277-281.
[5] ROOSTITA R,FLEET G H.The occurrence and growth of yeast in Camembert and blue-veined cheeses[J].International Journal of Food Microbiology,1996,28(3):293-404.
[6] TAMIME A Y,MARSHALL V M E.Microbiology and technology of fermented milks[M]//LAW B A.Microbiology and Biochemistry of Cheese and Fermented Milk.Boston,MA:Springer,1997.
[7] 李秋艳,夏先林,黄伟.日粮中添加微生态制剂对瘦肉型繁殖母猪生产性能的影响[J].贵州农业科学,2012,40(8):145-147,150.
[8] 闫俊书,宦海琳,周维仁,等.日粮中添加生物制剂对发酵床育肥后期猪生长性能、血清生化指标及内分泌激素的影响[J].中国畜牧兽医,2014,41(12):126-130.
[9] 唐明红,凌华云,王启军.微生态制剂对繁殖母猪生产性能和初生仔猪黄白痢发生率的影响[J].饲料工业,2014,35(17):114-117.
[10] 张德晓.复合益生菌的培养优化及其制剂在养猪生产中的应用[D].硕士学位论文.哈尔滨:东北农业大学,2014.
[11] 刘晴,黄华,唐景春,等.复合益生菌菌剂对母猪生产性能和肠道菌群生态的影响[J].家畜生态学报,2016,37(2):72-76.
[12] 霍军,宋予震,董青,等.日粮中添加微生态制剂对繁殖母猪生产性能的影响[J].江苏农业科学,2013,41(12):220-222.
[13] 付云超,薛琳琳,南楠.复合菌制剂对母猪繁殖机能影响的研究[J].黑龙江畜牧兽医,2015(18):89-90.
[14] 张国权,王国艳,乔国峰.复合微生态制剂对经产母猪繁殖性能及粗蛋白质表观消化率的影响[J].甘肃畜牧兽医,2016,46(23):99,101.
[15] 龙广.妊娠和泌乳日粮中添加布拉迪酵母菌对母猪及仔猪性能的影响[D].硕士学位论文.武汉:华中农业大学,2015.
[16] 于守洋,崔洪斌.中国保健食品的进展[M].北京:人民卫生出版社,2001.
[17] 郑建仙.功能性食品第三卷[M].中国轻工业出版社,1999:408-435.
[18] PEREYRA B S,LEMONNIER D.Induction of human cytokines by bacteria used in dairy foods[J].Nutrition Research,1993,13(10):1127-1140.
[19] 刘秉文,曾成林.高密度脂蛋白抗动脉粥样硬化作用[J].中国动脉硬化杂志,1994,2(1):44-50.
[20] STEINBERG D,PARTHASARATHY S,CAREW T E,et al.Beyond cholesterol.Modifications of low-density lipoprotein that increase its atherogenicity[J].New England Journal of Medicine,1989,320(14):915-924.
[21] 姜殿林,董桂虎,徐霞.正常妊娠妇女血脂及载脂蛋白水平的变化[J].实用临床医学,2006,7(4):16-17.
[22] 史艳明.妊娠期代谢综合征对母亲及胎儿的影响及其机制研究[D].硕士学位论文.长沙:湖南师范大学,2011.
[23] HERRERA E,樊中孟.妊娠期的脂肪代谢[J].国际儿科学杂志,1988(1):30-32.
[24] AGERHOLM-LARSEN L,BELL M L,GRUNWALD G K,et al.The effect of a probiotic milk product on plasma cholesterol:a meta-analysis of short-term intervention stuidies[J].European Journal of Clinical Nutrition,2000,54(11):856-860.
[26] GILLAND S E,NELSON C R,MAXWELL C.Assimilation of cholesterol byLactobacillusacidophilus[J].Applied & Environmental Microbiology,1985,49(2):377-381.
[27] CRILL J P,CAYUELA C,ANTOINE J M,et al.Effects ofLactobacillusamylovorusandBifidobacteriumbreveon cholesterol[J].Letters in Applied Microbiology,2000,31(2):154-156.
[28] TANNOCK G W,TANGERMAN A,VAN SCHAIK A,et al.Deconjugation of bile acids by lactobacilli in the mouse small bowel[J].Applied and Environmental Microbiology,1994,60(9):3419-3420.
[29] NOH D O,KIM S H,GILLILAND S E.Incorporation of cholesterol into the cellular membrane ofLactobacillusacidophilusATCC43121[J].Journal of Dairy Science,1997,80(12):3107-3113.
[30] FUKUSHIMA M,NAKANO M.Effects of a mixture of organisms,LactobacillusacidophilusorStreptococcusfaecalison cholesterol metabolism in rats fed on a fat-and cholesterol-enriched diet[J].British Journal of Nutrition,1996,76(6):857-867.
[31] 肖琳琳,董明盛.西藏干酪乳酸菌降胆固醇特性研究[J].食品科学,2003,24(10):142-145.
[32] 徐又先,濮德敏.氧化应激在妊娠方面的研究进展[J].中国优生与遗传杂志,2012,20(12):7-9.
[33] 肖文霞,马秀菊.氧化应激与妊娠[J].中国妇幼健康研究,2005,16(3):142-144.
[34] 李映雪,张琦,尚小明,等.参芍口服液对动脉粥样硬化大鼠血脂和氧化损伤的影响[J].河北联合大学学报(医学版),2017,19(6):457-461.
[35] 张江巍.乳酸菌抗氧化活性的研究[D].硕士学位论文.南昌:南昌大学,2006.
[36] 杨郁荭.白明.乳酸菌抗氧化机理的初步探讨[J].中国乳业,2011(7):68-73.
[37] 段希宇,叶陵,刘成国,等.乳酸菌的抗氧化作用机制[J].微生物学杂志,2017,37(3):111-115.
*Contributed equally
