猕猴桃属33份种质资源的AFLP遗传多样性分析
2018-04-17张世鑫吴绍华田维敏彭小列刘世彪
张 慧, 张世鑫, 吴绍华, 田维敏, 彭小列, 刘世彪
(1. 吉首大学 武陵山猕猴桃研究中心, 吉首 416000; 2. 中国热带农业科学院 橡胶研究所 农业部橡胶树生物学与遗传资源利用重点实验室, 儋州 571737)
猕猴桃(Actinidia)为猕猴桃科猕猴桃属的多年生木质藤本植物,其浆果营养丰富并具有药用价值。自20世纪初以来,通过对野生资源的人工选择驯化,猕猴桃已成为大规模商业化栽培最为成功的果树之一。中国是猕猴桃属植物起源、进化及分布的中心,中国原产的猕猴桃有62种(全世界66种),丰富的遗传资源是研究猕猴桃属植物演化、系统发育和分类的基础,也为猕猴桃品种选育和产业可持续发展提供了丰富的原材料[1-2]。由于猕猴桃属植物的种间杂交现象明显,尤其是不同变种间的杂交产生出的一系列过渡类型,以及复杂的染色体倍性,使得某些种间及变种间的分类界定不清楚,仅凭果实外观形态、生理特征来进行遗传多样性评价,有时难度较大。分子生物学手段可有效地运用于植物的遗传多样性和遗传差异分析,在猕猴桃的品种亲缘关系和性别鉴定方面已有应用[3-6]。其中扩增片断长度多态性(Amplified fragment length polymorphism, AFLP) 是一种基于PCR和限制性内切酶酶切的DNA分子标记技术[7],结合了RFLP技术的可靠性和PCR技术的高效性,且不受环境条件的限制,具有反应灵敏、快速高效、指纹多态性丰富、重复性好等优点。AFLP 标记已广泛应用于柑桔[8]、苹果[9]等果树的资源评价和遗传多样性分析,在猕猴桃方面也有报道[10-13]。
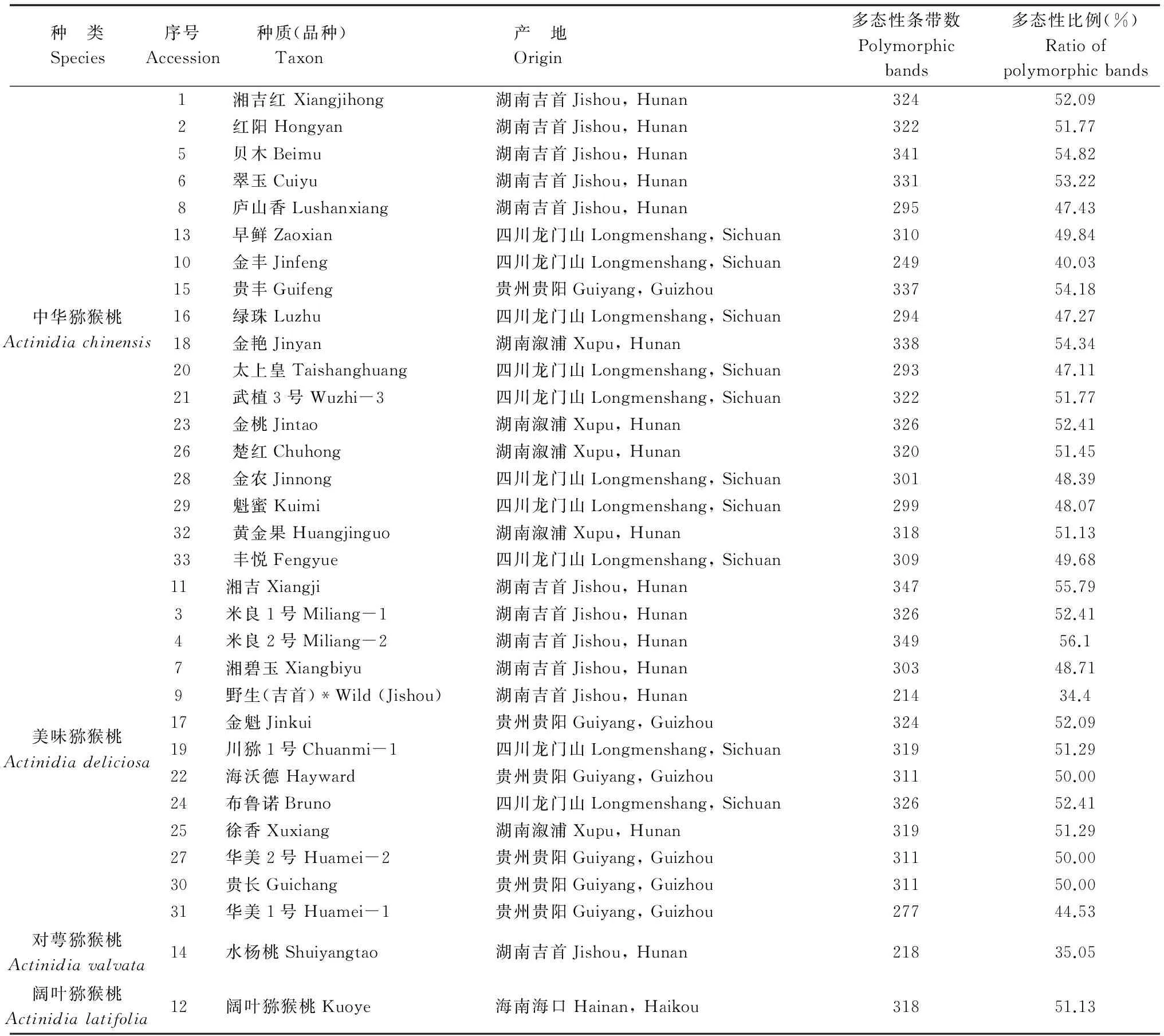
表1 猕猴桃属33份种质信息和AFLP分析结果
*为优良株系;其余为栽培品种或植物新品种
武陵山区是中国重要的猕猴桃自然分布区和果品生产区,其中的湘西土家族苗族自治州地处武陵山腹地,野生猕猴桃资源丰富,已选育了6个优良的猕猴桃品种和植物新品种(米良1号,湘吉,湘吉红,米良2号,湘碧玉,贝木)以及优良株系野生(吉首)猕猴桃,还引种栽培了数个国内优良品种,而有关该地区猕猴桃类群分布与遗传多样性的研究还未见报道。本研究利用AFLP标记技术对采自湖南湘西、贵州贵阳、四川都江堰和海南海口的33份猕猴桃种质资源进行了遗传多样性和遗传差异分析,以进一步了解其亲缘关系,并为猕猴桃遗传资源的保存和优良品种的筛选提供依据。
1 材料与方法
1.1 植物材料
2014年9月于湖南湘西的吉首、溆浦,四川都江堰龙门山,贵州贵阳,海南海口采集了33份猕猴桃种质资源,分属于4个物种:中华猕猴桃(Actinidiachinensis)18份,美味猕猴桃(A.deliciosa)13份,对萼猕猴桃(A.valvata)1份,阔叶猕猴桃(A.latifolia)1份,具体信息如表1所示。
1.2 样品处理和基因组DNA制备
每株取枝蔓顶部10 cm幼茎及初展的2~3片幼叶,每3株的材料做混合样,硅胶干燥保存备用。取1 g混合样品于研砵中加液氮磨成粉末,使用TaKaRa MiniBEST Plant Genomic DNA Extraction Kit [宝生物(大连)工程有限公司]提取和纯化样品的基因组DNA,以Thermo Gene Company Limited ND2000(Thermo Fisher Scientific, Inc., USA)测定DNA的含量,用1 %琼脂糖凝胶电泳检测DNA的完整性。
1.3 AFLP 引物对筛选和AFLP 分析
根据EcoR I和MseI酶切和接头序列,各设计筛选扩增引物8条(表2),并从64对引物中随机匹配23对引物进行AFLP实验(表3)。引物由英潍捷基(上海)贸易有限公司进行合成。AFLP分析参照Vos等[7]和秦小波等[14]方法进行,相关酶切反应及检测试剂和仪器购自New England Biolabs (Beijing) LTD.、Bio-Rad Laboratories, Inc.(USA)以及宝生物(大连)工程有限公司。
1.4 数据分析
用数字1和0,分别表示实验猕猴桃材料的某一等位变异的“有”和“无”,“有”带的值为1,“无”带的值为0。利用NTSYS-pc2.1[15]软件计算遗传相似系数和遗传距离,每两个种质之间的遗传距离GD=1-2Nij/(Ni+Nj),其中Ni和Nj分别为样品i和j具有带的条数,Nij为两样品共有带的条数。根据所得遗传距离,利用非加权配对算术平均法(unweighed pair group method arithmetic average, UPGMA)进行聚类分析,构建树状聚类图。
2 结果与分析
2.1 DNA纯化和AFLP 扩增
从图1可见,1%琼脂糖凝胶电泳检测使用TaKaRa MiniBEST Plant Genomic DNA Extraction Kit提取和纯化得到材料的DNA浓度和质量均较好(图1-A),使用Thermo Gene company Limited ND2000测定其含量,且A260/A280值范围为1.94~2.06,证明所得 DNA 样品的质量较好,可以用于AFLP分析猕猴桃种质的遗传差异性。对PCR扩增产物进行检测,发现PCR扩增的条带弥散且清晰,条带大小约为100~1500 bp(图1-B),可以用于变性聚丙烯酰胺凝胶电泳。选用23对AFLP选择性引物对33份猕猴桃属植物的材料进行AFLP分析,均得到清晰的指纹(图1-C),且各引物组合的指纹图谱反映出所有种质均表现出多态性,说明AFLP技术在猕猴桃种质鉴定中是可行的。

图1 猕猴桃DNA 和AFLP 选择性扩增产物的聚丙烯酰胺凝胶电泳图Fig 1 DNA and polyacrylamide gel electrophoresis of AFLP selective amplification for kiwifruits
A:部分猕猴桃品种的DNA;B:部分PCR 扩增产物;C:EcoR I-3/MseI-1扩增的PCR产物

表2 AFLP所用接头和引物序列
2.2 猕猴桃属33份种质的遗传多样性
表1为33份猕猴桃种质资源的产地及AFLP的分析结果,它显示各种质的扩增条带分布均匀。在23对引物所获得的622个多态性位点中,最多的米良2号的多态性带数为349条,多态性比例达56.1%,显示其杂合程度高;最少的是野生猕猴桃(吉首),多态性带仅有214条,多态性比例34.4%。同时,对萼猕猴桃的多态性带条偏少(249条),多态性比例40.03%,而阔叶猕猴桃的多态性带为318条,多态性比例51.13%,处于中等水平。
表3为23对AFLP引物序列及扩增条带信息。23对引物共获得了683个遗传位点,即33份种质共获得683条扩增谱带,平均每对引物的遗传位点为29.7个,其中多态性位点共622个,多态性比例平均为91.07%,对33个种质材料的区分率达到100%。上述数据表明猕猴桃属植物在分子水平上存在丰富的遗传多样性,每对引物的多态性位点都较丰富,最高的多态性比例高达100%,最低的有50.0%,平均条带/种质16.1,说明上述方法完全可以区分这些种质。

表3 AFLP引物组合及扩增条带
2.3 猕猴桃属33份种质的遗传距离和聚类分析
根据UPGMA软件对33份猕猴桃种质遗传距离的分析,各猕猴桃种质间均存在一定的遗传距离,其值在0.05与0.83之间,平均为0.2325,遗传距离越小,亲缘关系越近,反之越远。如有些品种间的遗传距离很小,例如1号(湘吉红)与2号(红阳)的遗传距离最小仅为0.05,5号(贝木)与6号(翠玉)的遗传距离为0.07,它们的相似性很高,说明在进化过程中由于自然选择,其生物性状和遗传物质趋于一致。与其他种质遗传距离较大的有10号金丰(中华猕猴桃),平均值为0.31,9号野生美味猕猴桃(平均0.367),12号阔叶猕猴桃(平均值0.37),阔叶猕猴桃与吉首野生美味猕猴桃的遗传距离为0.48,显示了较远的亲缘关系。而遗传距离最大的是14号水杨桃(平均值0.70),它与吉首野生猕猴桃之间的遗传距离系数达0.83,说明两个物种间的亲缘关系更远,这与各自的花、果实和叶等植物学性状的差异是相符合的。
利用UPGMA对23对引物组合检测出的683个遗传位点相关性进行聚类,如图2所示。33份供试猕猴桃属种质之间相似系数为0.51~0.95,显示猕猴桃种质间的遗传关系存在一定距离。在相似系数0.70的水平,可以将供试的33个材料分为3个类群,Ⅰ类:包括所有的中华猕猴桃和美味猕猴桃物种;Ⅱ类:阔叶猕猴桃;Ⅲ类:水杨桃(对萼猕猴桃)。阔叶猕猴桃与中华猕猴桃和美味猕猴桃的相似系数较小,亲缘关系较远,而水杨桃(对萼猕猴桃)与前两者的相似系数最小,亲缘关系最远。在最大的第I类分支中,共包括31份样品,在0.82水平上又可以将其主要再分为Ⅳ类、Ⅴ类和其他细支,Ⅳ类包括湘吉红、红阳、米良1号、米良2号、湘碧玉、庐山香、贝木、翠玉和湘吉,Ⅴ类包括早鲜、金艳、川猕1号、武植3号、金桃、海沃德、布鲁诺、徐香、华美2号、魁蜜、黄金果、金农、楚红、贵长、华美1号、丰悦、贵丰和金魁。与Ⅳ、Ⅴ类遗传距离较远的分支有太上皇、绿珠、野生(吉首)和金丰。
3 讨论
AFLP标记的对象是双酶切处理的基因组DNA,获得的指纹图谱多态性丰富。该技术因其高效而稳定,重复性强,被广泛应用在作物资源评价和遗传多样性分析。本研究用23对引物共获得了33份猕猴桃种质资源的683个遗传位点,其中多态性位点622个,多态性比例平均为91.07%,区分率达100%。证明AFLP标记扩增效率高,是分析猕猴桃品种遗传差异的一种有效的手段。
这些猕猴桃种质资源的遗传相似系数在0.51~0.95之间,说明猕猴桃种质的多样性水平较高。图2表明,4个种33份猕猴桃种质资源基本上按照种分别聚在一起,尤其是阔叶猕猴桃和对萼猕猴桃(水杨桃)按组系关系聚类现象明显,与徐小彪[11]的研究结果相似,显示出华南四省猕猴桃的遗传多样性差异较大。种质的亲缘关系在一定程度上与地理分布有关。例如,6个湖南吉首地区选育的品种中,米良1号和米良2号的相似系数为0.89,湘碧玉与之关系较近,黄肉品种贝木和无籽猕猴桃品种湘吉距离较远,而红肉型的湘吉红品种距离最远,但所有这些种质都聚在Ⅳ类中,相似系数达到了0.8(图2),反映了在相同的环境中长期的进化选择使其亲缘关系更近。又如海沃德、华美2号、贵长、华美1号、贵丰、金魁等几个品种被归为一类,这批材料均采自贵阳,其相似系数达0.83,而采自湖南溆浦县的金艳、金桃、徐香、黄金果和楚红都聚于V类的上半部分,相似系数0.86,显示出很近的亲缘关系,证明猕猴桃具有按照地理来源聚类的趋势[16]。通常同一物种不同品种间的亲缘关系较近,如中华猕猴桃的红阳与湘吉红,贝木和翠玉,美味猕猴桃的米良1号和米良2号,贵长和华美1号。但中华猕猴桃和美味猕猴桃之间具有较高的基因流、存在共祖多态和遗传渐渗现象等,两者存在着密切的亲缘关系和聚类交叉关系[17-20],这种现象在陈华等[21]对江西31份猕猴桃种质和秦小波等[14]对西南地区10种猕猴桃50个样本的遗传多样性研究中得到支持,本文中的金艳与川猕1号,魁蜜与华美2号,湘碧玉和庐山香,金丰与野生美味猕猴桃等也是如此,这也与我国猕猴桃育种实践中常采用亲缘关系较近的亲本杂交有关,从而使育成品种之间的遗传距离逐渐下降。尤其是吉首野生的美味猕猴桃,单果较大,果形圆柱形,与原产于湘西凤凰县的米良1号和米良2号差别很大,现证明其间的遗传差异也很大,该优株有进一步研发的前景,也可以作为一个新的杂交种质资源。
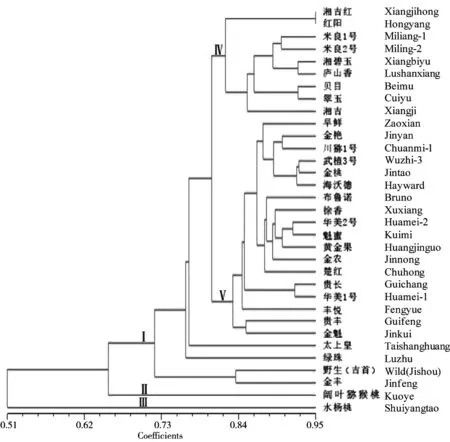
图2 基于AFLP 标记的猕猴桃属33份种质遗传距离的UPGMA 聚类分析Fig 2 UPGMA dendrogramof 33 kiwifruit germplasms based on AFLPs
采自海南海口的阔叶猕猴桃(斑果组)和采自湖南吉首的水杨桃(对萼猕猴桃,净果组),均为单独的物种,他们彼此间及与中华猕猴桃和美味猕猴桃(同属星毛组)品种间的相似系数小,遗传距离大,亲缘关系也最远,符合传统的形态分类结论。从猕猴桃亲缘关系聚类图上可以看出各种质的亲缘关系远近及其在分类上的系统地位。因此,传统的形态分类结合分子标记,从分子水平上反映猕猴桃属种质之间的亲缘关系,可以作为猕猴桃种质分类及选育种的评价依据。
致谢:特别感谢为本研究提供实验材料的四川龙门山猕猴桃基因库、贵阳市乌当区开元猕猴桃研究所、海南省农业科学院、湖南永鑫特色农业开发专业合作社。
[1]HUANG H W, WANG Y, ZHANG Z H, et al. Actinidia germplasm resources and kiwifruit industry in China[J]. Hort Science, 2004, 39(6): 1165-1172.
[2]FERGUSON A R, HUANG H W. Genetic resources of kiwifruit: domestication and breeding[J]. Horticultural Reviews, 2007, 33:1-121.
[3]井赵斌, 徐 明, 雷玉山. 猕猴桃SRAP-PCR体系的建立及品种资源亲缘关系研究[J]. 园艺学报,2016,43 (2): 337-346.
[4]刘 娟, 廖明安, 谢 玥, 等. 猕猴桃属16个雄性材料遗传多样性的ISSR分析[J]. 植物遗传资源学报, 2015, 16(3): 618-623.
[5]ZHANG Q, LIU C Y, LIU Y F, et al. High-density interspecific genetic maps of kiwifruit and the identification of sex-specific markers[J]. DNA Research, 2015, 22(5), 367-375.
[6]HILARIO E, BARRON L, DENG C H, et al. Random tagging genotyping by sequencing (rtGBS), an unbiased approach to locate restriction enzyme sites across the target genome [J]. Plos One, 2015, 10(12):e0143193.
[7]VOS P, HOGERS R, BLEEKER M, et al. AFLP: a new technique for DNA fingerprinting[J]. Nucleic Acids Research, 1995, 23(21): 4407-4414.
[8]OLIVEIRA A C, BASTIANEL M, CRISTOFANI-YALY M, et al. Development of genetic maps of the citrus varieties Murcott′ tangor and Pera′ sweet orange by using fluorescent AFLP markers [J]. Journal of Applied Genetics, 2007, 48(3): 219-231.
[9]PARIS R, DONDINI L, ZANNINI G, et al. dHPLC efficiency for semiautomated cDNA-AFLP analyses and fragment collection in the apple scab-resistance gene model [J]. Planta, 2012, 235(5): 1065-1080.
[10]张潞生, 李传友, 贾建航,等. 猕猴桃雌雄性别的AFLP鉴别中DNA模板的制备[J]. 果树科学, 1999, 16 (3):171-175.
[11]徐小彪. 猕猴桃属植物的遗传多样性及种质超低温保存研究[D]. 长沙:湖南农业大学,2004.
[12]陈 华, 易干军, 徐小彪,等. 猕猴桃AFLP分析体系的建立[J]. 西北植物学报, 2005, 25(8):1528-1535.
[13]吕天雯, 栗望薇, 张太奎, 等. 猕猴桃AFLP分析体系的建立及其组培变异苗的动态检测[J]. 浙江农业学报, 2016, 28(4): 618-623.
[14]秦小波,高继海.利用AFLP分析西南特色猕猴桃的遗传多样性[J]. 热带亚热带植物学报,2013,21(4):315-322.
[15]Rohlf F J. NTSYSpc: numerical taxonomy and multivariate analysis system[CP]. New York: Exeter Software, Applied Biostatistics Inc, 2000:16-29.
[16]岁立云, 刘义飞, 黄宏文. 红肉猕猴桃种质资源果实性状及AFLP遗传多样性分析[J].园艺学报, 2013, 40(5): 859-868.
[18]刘亚令, 李作洲, 张鹏飞, 等. 猕猴桃自然居群SSR遗传变异的空间自相关分析[J].生物多样性,2006,14(5):421-434.
[19]LIU Y F, LIU Y L, HUANG H W. Genetic variation and natural hybridization among sympatricActinidiaspecies and the implications for introgression breeding of kiwifruit [J]. Tree Genetics & Genomes, 2010, 6(5): 801-813.
[20]饶静云, 刘义飞, 黄宏文. 中华猕猴桃不同倍性间杂交后代倍性分离和遗传变异分析[J]. 园艺学报, 2012, 39(8): 1447-1456.
[21]CHEN H , YI G J, XU X B. Identification of the kiwifruit germplasms in Jiangxi province by AFLP and its classification significance [J]. Chinese Journal of Biochemistry and Molecular Biology, 2007, 23(2):122-129.
