酿酒酵母在传代过程中生理变化研究进展
2018-04-12丁华建王金晶朱佳琪郑梅莹郑飞云李永仙钮成拓刘春凤李崎
丁华建,王金晶,朱佳琪,郑梅莹,郑飞云,李永仙,钮成拓,刘春凤,李崎
1 江南大学 生物工程学院 工业生物技术教育部重点实验室,江苏 无锡 214122
2 江南大学 酿酒科学与工程研究室,江苏 无锡 214122
酿酒酵母Saccharomyces cerevisiae是人类了解最早、应用最广泛的一种酵母,主要应用于面包、酒类等发酵。酿酒酵母具有生长周期短、发酵能力强、容易进行大规模培养以及含有多种蛋白质、氨基酸、维生素、生物活性物质等丰富的营养成分等优点,一直是基础及应用研究的主要对象,在食品、医药等领域应用广泛。酵母的重复利用在工业生产尤其是酿造行业中被广泛接受并应用,以降低生产成本,使菌株发挥更大的应用价值。然而随着回用次数的增加,酿酒酵母不可避免地出现逐渐衰老的现象,导致其发酵能力发生下降至影响最终产品的质量[1],因此在生产中对酵母的活力以及性能提出了更高的要求。
对细胞衰老机理的研究一直是生命科学领域所关注的问题,而 Botstein等发现酿酒酵母有将近31%编码蛋白质的基因或者开放阅读框与哺乳动物编码蛋白质的基因有高度的同源性[2],因此酿酒酵母被认为是研究细胞生理分子机制以及高等真核生物病理生理过程的重要模型系统,常被用于衰老机制的研究[3]。衡量酿酒酵母的衰老有两种模式,时序衰老 (Chronological aging) 和复制衰老 (Replicative aging)。而与此相应的酿酒酵母的寿命也有两种定义方式,即时序寿命(Chrononical lifespan,CLS) 和复制寿命(Replicative lifespan,RLS),前者指一定条件下单个细胞的存活时间,后者指酵母细胞在死亡之前的分裂次数[4-5]。酿酒酵母在连续传代的情况下,酵母的各项生理指标会因为一些胁迫条件 (如温度高低、乙醇积累、pH上升以及渗透压上升等)的变化,在某种程度上影响其衰老。本文综述了酿酒酵母在连续传代过程中细胞形态结构的变化、细胞的絮凝性、活力以及胞内多种酶活性等的变化趋势。
1 细胞形态
细胞的形态结构决定了细胞的功能,而细胞的功能则影响了细胞的形态结构变化。酵母细胞在连续传代过程中,培养环境以及培养基成分在转接时发生剧烈变化,酵母在感受到外界环境变化时会启动其自身系列调控系统用以维持细胞的基本形态、大小以及出芽的比率等。
正常发酵过程中,酵母细胞一般多呈椭圆形、圆形,表面圆润饱满,有明显的出芽现象和芽痕。通过对不同代数的酵母细胞进行观察,发现第 1代和第2代的酵母细胞表面圆润饱满、光滑无褶皱,且大部分为椭圆形的单细胞状态,细胞之间没有相互附着现象。但是到了第15代之后,细胞表面就会布满褶皱,形态近似圆形,有许多细胞聚集成团,只有个别的细胞还处于单细胞状态[6]。但是当这些酵母细胞处于营养匮乏的环境中时,通过扫描电镜 (Scanning electronic microscopy,SEM) 和透射电镜 (Transmission electron microscope,TEM) 对细胞进行观察,发现一般会在第5代左右就出现严重的褶皱变化,细胞的内部结构会逐渐水解,同时细胞的大小也发生一定的变化(图 1、2)。说明随着传代次数的增加,细胞衰老过程在细胞形态尤其是细胞壁上的反应愈加明显。同时研究还表明经过多次分裂后,细胞壁表面的芽痕[7]增多,细胞的形态趋向于圆形,这可能是胞内抗氧化酶在经过多次传代后活性降低,引起的胞内自由基积累,而自由基的存在对细胞产生了毒害作用,从而引起细胞形态的变化[8]。再者,在传代的过程中由于环境变化的影响,酿酒酵母不可避免地发生自溶[9],其细胞形态也发生了大幅度的变化。
2 细胞壁
酿酒酵母细胞壁是一个动态的且被调控得非常有序的细胞结构,其结构和组成被严格调控并能通过全局应答系统对环境变化作出广泛地响应[10]。

图1 第1代与第5代酿酒酵母在扫描电镜下的形态变化Fig. 1 The SEM image of S. cerevisiae of first and fifth generation. (A) Morphology of the G1 yeast cells under the condition of nutrient deficiency. (B) Morphology of the G5 yeast cells under the condition of nutrient deficiency.
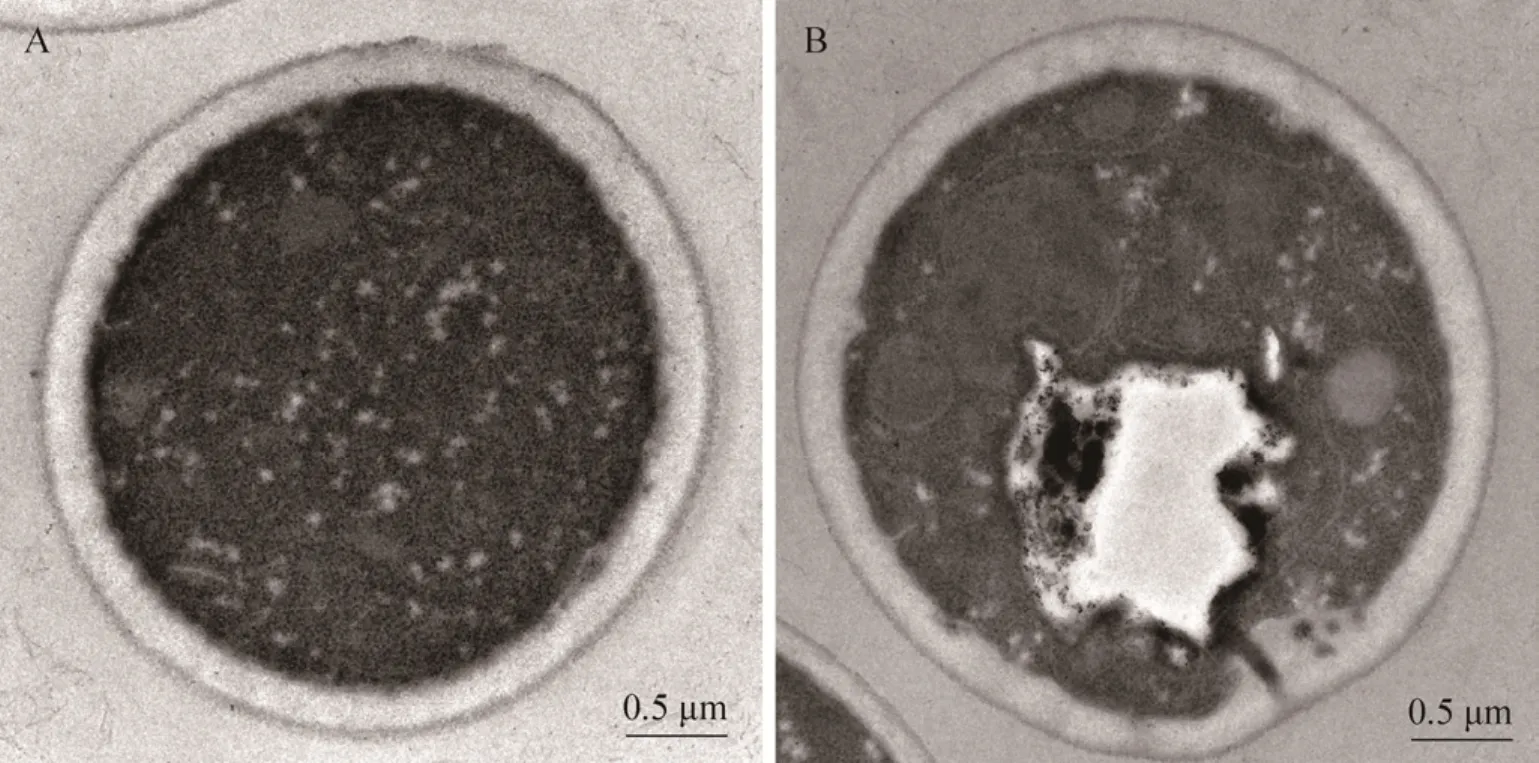
图2 第1代与第5代酿酒酵母在透射电镜下的形态变化Fig. 2 The TEM image of S. cerevisiae of first and fifth generation. (A) Morphology of the G1 yeast cell under the condition of nutrient deficiency. (B) Morphology of the G5 yeast cell under the condition of nutrient deficiency.
酿酒酵母细胞壁为双层结构,内层是由β-1,3-葡聚糖和 β-1,6-葡聚糖组成的葡聚糖层,在出现牙痕的附近还含有一定数量的几丁质。外层是由性质不同的各种甘露糖蛋白组成。在酵母细胞壁中,甘露糖蛋白是酵母细胞壁的重要组成部分,它除了可以参与酵母细胞之间的交配外,还与菌落形态变化以及生物大分子的免疫识别等相关[11]。当酵母细胞的生长环境发生异常变化或细胞壁结构受到破坏时,细胞壁表面感受压力因子将信号传递给细胞,然后再通过细胞壁完整性 (Cell wall integrity,CWI) 途径或高渗透性甘油(High osmolarity glycerol,HOG) 途径等及时进行调节,来抵抗不良环境,减少对细胞的损害[12-13]。
成建国[14]发现酿酒酵母在传代的过程中,酵母细胞壁内层的骨架结构成分葡聚糖所占比例随着代数的增加逐渐积累,而对细胞壁蛋白起修饰作用的甘露糖含量则逐渐减少。研究表明当这些酵母细胞处于营养匮乏的环境中时,细胞壁的变化更加明显,正常发酵条件下酵母细胞壁的厚度增加趋势缓慢,而在营养匮乏的条件下,厚度增加的趋势明显加快,第一代细胞壁厚度维持在0.2 μm左右,到了第5代已经达到0.3 μm左右。分析酵母菌骨架结构层 (葡聚糖) 的聚合度变化时,发现在传代过程中,酵母菌细胞壁内层葡聚糖的聚合度随着代数的增加而逐渐降低,而且葡聚糖组成中的 β-1,6-糖苷键比例降低,β-1,3-糖苷键比例增加。
酵母菌在传代的过程中,由于细胞的代谢能力逐渐下降,使细胞内的甘露糖合成酶活力下降,对细胞壁蛋白质的糖基化修饰就会降低,最终导致甘露糖含量的下降。虽然甘露糖的含量在逐渐减少,但是菌体自身为了维持正常的生理代谢活动,通过CWI以及HOG等途径调节发生快速修复,合成更多的葡聚糖来弥补甘露糖的减少以及其自身聚合度的下降对细胞造成的伤害,因此总体上仍然导致了细胞壁的加厚。由于酵母内壁葡聚糖层逐渐变厚,它的增厚导致菌体外形发生微小变化,由椭圆形逐渐趋于圆形。酵母菌细胞壁出现这些重要变化是酵母菌自我保护的一种代谢调节能力的体现,因此,当酵母菌对环境等不良因素的抵抗能力减弱时就会通过合成更多的葡聚糖加厚细胞壁来阻止外界大分子物质进入细胞内破坏细胞[15]。
3 絮凝性
絮凝性是酵母在发酵工业如啤酒酿造中体现的一个重要特性。目前对酵母絮凝性机理有多种假说,但影响最广泛的就是类外源凝集素 (Lectin-like)假说,即酵母细胞外壁的表面蛋白质或糖蛋白与其他酵母细胞壁的甘露糖残基特异性结合导致絮凝[16]。从分子学的角度来看,酵母絮凝是由整个FLO基因家族编码的絮凝蛋白与邻近细胞的胞壁甘露糖结合形成的多细胞聚集沉降现象,是成千上万个细胞形成的可逆的、无性的、需要Ca2+的过程[17-18]。酵母的絮凝不仅受遗传因子的控制,而且受环境和生理等因素的影响[19]。酵母细胞在连续传代的过程中,细胞会逐渐衰老,而酵母的衰老影响细胞的表面特性,尤其是细胞表面的疏水性,从而影响酵母细胞的絮凝性[20]。
酵母细胞在传代的过程中,其细胞形态及表面特性会逐渐发生变化,与母细胞相比,子细胞的再生殖有所缓慢,絮凝能力有所降低[21]。大多数酿酒酵母在发酵初期丧失絮凝能力,而在指数后期或稳定期时酵母细胞表面的疏水相互作用达到最大,粘附性最强,因此又重新获得絮凝的能力[22]。一般菌龄较长的酵母细胞絮凝能力更强。另外,形态学研究表明,随酵母细胞菌龄的增长,细胞壁出现褶皱状变化,细胞间联系更加紧密,细胞间的粘连更加容易,这可以部分解释酵母在不同生长时期表现出不同絮凝能力的现象。
也有许多研究者认为,酵母菌细胞壁在絮凝过程中起着重要的作用,而其中的细胞壁蛋白质在酵母菌的絮凝中发挥着更加明显的作用[23-24]。安文涛等[25]以及 Potter[26]发现在酵母菌连续传代过程中,酵母菌细胞壁蛋白质的糖基化修饰程度在减弱,而糖基化修饰程度的降低使大量蛋白质在酵母菌细胞表面裸露,这对细胞表面张力的维持带来了一定的负作用,引起细胞壁表面各种力的变化,最终导致酵母菌絮凝性加强。
4 细胞活力
人类利用酵母发酵生产各种食品、化工产品已有非常长的历史,然而酵母发酵产生这些代谢产物也会造成酵母菌生存环境的不断改变,使其面临各种胁迫条件的影响,如温度、乙醇、pH以及渗透压等[27]。因此在进行连续传代的过程中,酵母的活力会因为这些胁迫条件的影响而发生一些改变。
评价细胞的活力有两种方式,一个是活细胞的占比率 (Viability);另一个是细胞的生理能力(细胞存活,但是无分裂能力),也叫细胞活性(Vitality)。通常情况下两种方式需要结合来反映细胞的活力[28]。研究表明,酵母细胞在传代的过程中,其细胞活力逐渐下降,其活力的下降趋势和程度在不同菌株之间又表现不同。对两株lager型啤酒酵母Pilsner和5-2进行研究发现,比较0代和5代酵母,其死亡率都从0上升至5%左右,而菌株的活性下降则有明显的区别。Pilsner和 5-2的活性分别由0代的99%下降到第5代的45%和60%左右,其细胞活性的下降正说明了其不断衰老的过程。另外,酵母细胞的活力除了反映生理功能,还包括了细胞的代谢活力。由此也证明了细胞的死亡率并不能作为单一的评价指标对酵母的活力活性进行评价。
酵母的主要能量来源为葡萄糖,对葡萄糖的代谢能力直接反映了细胞的代谢活力。葡萄糖代谢活力以培养基或发酵液中残留葡萄糖浓度来表示,残留的葡萄糖浓度越低酵母的代谢越快,代谢活力就越强。姚继兵等[6]、季杨杨[29]等研究发现当酿酒酵母连续培养到第 10代时其降糖的速率比第2代略有降低,而当培养到第15代时与第10代相比,降糖速率明显降低。此外,不同代数酵母在指数生长期时降糖速率基本一致,而在生长稳定期时,不同代数酵母降糖速率有明显差异。出现这种现象的原因可能是指数期酵母分解葡萄糖是用于细胞的分裂生长繁殖,而在稳定期表现的现象则说明随着酵母菌的传代次数增多,酵母的细胞活性下降,发酵能力降低,代谢能力越来越弱。
另外,也有一些学者对提高细胞活力和活性进行了研究。海藻糖是一类非常重要的压力保护剂,海藻糖的抗氧化压力主要体现在其保护细胞免受氧自由基破坏。研究发现,与海藻糖降解相关的基因 (NTH1、NTH2、ATH1) 受应激反应元件 (The stress responsive element,core consensus CCCCT,STRE) 的调控[30-31]。在酵母衰老的过程中,与海藻糖降解相关基因NTH1和ATH1的表达水平发生了相应的下调,使海藻糖降解速度减慢,从而保证了海藻糖在酵母细胞中的积累,保护细胞不受氧自由基损伤[32]。因此Hirasawa等通过在酵母细胞的培养中添加适量海藻糖,发现细胞的抗冻性会显著增加,同时细胞的发酵能力和活性会有效改善[33];此外,Magalhães等研究表明丰富的黄腐酚虽然对酵母活性没有显著影响,但对酵母活力却有一定的积极作用,同时黄腐酚对酵母的生理状态也有积极的影响[34]。
5 胞内酶活性变化
酵母细胞内含有多种酶,如抗氧化酶、蛋白酶、脂肪酶、蔗糖酶、淀粉酶等[35-37]。在工业生产中,随着酵母使用代数的增加,酵母酶活力因为各种培养条件或胁迫环境等因素的影响发生变化,而其酶活力高低则直接影响发酵的效率和产物质量。
5.1 抗氧化酶
酵母细胞内的抗氧化酶包括超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT) 和过氧化物酶 (Peroxidase,POD)。SOD、CAT和POD是生物体内主要抗氧化酶系统。在正常情况下,3种酶联合清除活性氧自由基(ROS),保护自身免受自由基伤害[38]。
研究发现[39]随着酵母传代次数的增加,酵母细胞内3种抗氧化酶的活性呈现出先增加后减小的变化趋势,因此导致了胞内活性氧自由基的含量出现了先下降后上升的现象,这与在前期研究中发现的胞内 ROS出现先降低再升高的趋势相一致。抗氧化酶系的作用下使细胞中的自由基被快速清除,ROS出现下降趋势,使得细胞不被自由基破坏;当酶活性上升到最大值后开始下降,细胞内自由基的积累和清除失去平衡。因此,过多的ROS积累氧化细胞膜脂、蛋白质以及DNA等生物大分子,会对细胞产生不可逆的伤害,细胞结构会受到一定程度的损伤[40],这也解释了为什么酵母细胞在经过连续传代后细胞表面布满褶皱,细胞呈现逐渐衰老的趋势。
5.2 其他酶类
在酵母细胞内除了有保护细胞免受活性氧自由基伤害的抗氧化酶外,还有在酵母的生理代谢活动中起着重要作用的蛋白酶、脂肪酶、蔗糖酶等一些酶类。
张利等[41]通过对不同代数的酵母进行蛋白酶、脂肪酶和蔗糖酶等酶活力测定,发现酵母蛋白酶活力随着酵母代数的增加而升高,而脂肪酶以及蔗糖酶活力都在逐渐下降。在酵母连续传代过程中,当细胞内蛋白酶活力得到明显增加时,就会加速催化水解酵母细胞壁和细胞膜上的蛋白质,蛋白质分子水解成游离氨基酸,使细胞失去稳定的结构和活性。脂肪酶、蔗糖酶在酵母细胞中的作用主要是通过水解大分子的脂肪、蔗糖等物质以供给细胞代谢所需要的能量,酵母细胞内的脂肪酶和蔗糖酶的活性在传代的过程中逐渐降低,说明酵母在传代的过程中对营养物质的利用率逐渐降低,反映了酵母的代谢能力在逐渐减弱,细胞在逐渐衰老。
6 胞外多糖的代谢
胞外多糖 (Extracellular polysaccharides,EPS)[42]由微生物产生后分泌到细胞外,一般由单糖和一些非糖基 (如乙酸盐、丙酮酸、琥珀酸等)所组成,是易与菌体分离且可以通过深层发酵实现工业化生产的一类代谢物。由于胞外多糖的组成具有多样性,因此胞外多糖在各种食品和制药行业有着广泛应用。酿酒酵母胞外多糖作为多糖中的一种,具有多种生理活性,除了能够提高机体免疫活性外,还具有抗病毒、抗氧化和抑制肿瘤生长等作用[43]。而酵母多糖除了甘露寡聚糖、β-葡聚糖等细胞壁多糖外,更多的是与酵母菌生理密切相关的,由葡萄糖在酵母菌细胞内合成,然后通过某种机制分泌到细胞外的胞外多糖[44]。
研究发现[45],在进行酵母菌传代培养的过程中,随着传代次数的增加,酵母菌的代谢能力逐渐下降,胞外多糖的分泌速度和含量也都在降低。酵母菌胞外多糖含量的降低与其代谢糖能力降低是密切相关的,在这个层面上多糖含量的变化与残糖消耗的减少表现出了一致性,然而其占葡萄糖代谢总量的比例却随着传代次数的增加有了小幅度的提升,可以推测,酵母菌胞外多糖与酵母菌抗衰老有一定的关系。
7 展望
酵母的回收利用在工业生产中不可避免,而酵母在传代过程中生理变化及衰老机制的研究并不深入。虽然了解到酵母的时序衰老和复制衰老模式,但是酵母的衰老是一个多因素的现象,其详细的机制仍然未知,所以这一问题的成功解决将为酵母大工业生产中存在的问题起到积极影响,另外也为多细胞真核生物的衰老研究提供研究基础。结合当前研究中遇到的困难,可以尝试从以下两方面进行研究:1) 虽然酿酒酵母的衰老有两种模式,但是两者之间也有许多相同点,如氧化压力的增加、细胞凋亡的发生等。可以尝试跟踪两者共有特征来研究酵母的衰老。2) 酵母在传代过程中各种组学信息包括转录组学、蛋白组学以及代谢组学的分析将为酵母细胞在传代中的生理代谢变化提供遗传信息支持,有助于对酵母的生理变化及对外界环境压力应答有一个更加全面系统的了解。通过对酵母细胞衰老机制的不断探究,能够更全面地认识酵母在工业应用中的衰老,将为酵母在工业生产中更广泛的应用提供理论基础。
[1]Gibson BR,Lawrence SJ,Boulton CA,et al. The oxidative stress response of a lager brewing yeast strain during industrial propagation and fermentation.FEMS Yeast Res,2008,8(4): 574–585.
[2]Botstein D,Chervitz SA,Cherry JM. Yeast as a model organism. Science,1997,277(5330):1259–1260.
[3]Gershon H,Gershon D. The budding yeast,Saccharomyces cerevisiae,as a model for aging research: a critical review. Mech Ageing Dev,2000,120(1/3): 1–22.
[4]Longo VD,Shadel GS,Kaeberlein M,et al.Replicative and chronological aging inSaccharomyces cerevisiae. Cell Metab,2012,16(1):18–31.
[5]Fabrizio P,Longo VD. The chronological life span ofSaccharomyces cerevisiae. Aging Cell,2003,2(2):73–81.
[6]Yao JB,Zu GR,Piao YZ,et al. Proteomic analysis of cell wall proteins fromSaccharomyces cerevisiaewith different generation numbers. Microbiol China,2013,40(11): 1962–1969 (in Chinese).姚继兵,祖国仁,朴永哲,等. 不同传代次数的酿酒酵母细胞壁蛋白组学分析. 微生物学通报,2013,40(11): 1962–1969.
[7]Wang XX,Wang X,Wang LZ,et al. Influencing factors of self-flocculation of yeast. China Brew,2011,30(4): 5–10 (in Chinese).王肖肖,王侠,王立志,等. 酵母自絮凝特性的影响因素. 中国酿造,2011,30(4): 5–10.
[8]Ayer A,Gourlay CW,Dawes IW. Cellular redox homeostasis,reactive oxygen species and replicative ageing inSaccharomyces cerevisiae. FEMS Yeast Res,2014,14(1): 60–72.
[9]Schiavone M,Sieczkowski N,Castex M,et al.Effects of the strain background and autolysis process on the composition and biophysical properties of the cell wall from two different industrial yeasts. FEMS Yeast Res,2015,15(2):fou012.
[10]Smits GJ,van den Ende H,Klis FM. Differential regulation of cell wall biogenesis during growth and development in yeast. Microbiology,2001,147(4):781–794.
[11]Aguilar-Uscanga B,Francois JM. A study of the yeast cell wall composition and structure in response to growth conditions and mode of cultivation. Lett Appl Microbiol,2003,37(3): 268–274.
[12]Rodríguez-Peña JM,García R,Nombela C,et al. The high-osmolarity glycerol (HOG) and cell wall integrity (CWI) signalling pathways interplay: a yeast dialogue between MAPK routes. Yeast,2010,27(8): 495–502.
[13]Klis FM,Boorsma A,de Groot PW. Cell wall construction inSaccharomyces cerevisiae. Yeast,2006,23(3): 185–202.
[14]Cheng JG. The research of cell wall composition and related enzymatic properties of saccharomyces cerevisiae aging process [D]. Dalian: Dalian Polytechnic University,2012 (in Chinese).成建国. 酿酒酵母衰老过程中细胞壁组成及相关酶学性质研究[D]. 大连: 大连工业大学,2012.
[15]Kwiatkowski S,Thielen U,Glenney P,et al. A study ofSaccharomyces cerevisiaecell wall glucans. J Inst Brew,2009,115(2): 151–158.
[16]Ngondi-Ekome J,Thiebault F,Strub JM,et al. Study on agglutinating factors from flocculentSaccharomyces cerevisiaestrains. Biochimie,2003,85(1/2): 133–143.
[17]Javadekar VS,Sivaraman H,Sainkar SR,et al. A mannose-binding protein from the cell surface of flocculentSaccharomyces cerevisiae(NCIM 3528):its role in flocculation. Yeast,2000,16(2): 99–110.
[18]Bidard F,Bony M,Blondin B,et al. TheSaccharomyces cerevisiae FLO1flocculation gene encodes for a cell surface protein. Yeast,1995,11(9): 809–822.
[19]Verstrepen KJ,Derdelinckx G,Verachtert H,et al.Yeast flocculation: what brewers should know. Appl Microbiol Biotechnol,2003,61(3): 197–205.
[20]Vichi S,Gallardo-Chacón JJ,Pradelles R,et al.Surface properties ofSaccharomyces cerevisiaelees during sparkling wine ageing and their effect on flocculation. Int J Food Microbiol,2010,140(2/3):125–130.
[21]Powell CD,Quain DE,Smart KA. The impact of brewing yeast cell age on fermentation performance,attenuation and flocculation. FEMS Yeast Res,2003,3(2): 149–157.
[22]Chang YI,Shih LH,Chen SW. Effects of glucose and mannose on the flocculation behavior ofSaccharomyces cerevisiaeat different life stages.Colloids Surf B Biointerfaces,2005,44(1): 6–14.
[23]Speers RA,Wan YQ,Jin YL,et al. Effects of fermentation parameters and cell wall properties on yeast flocculation. J Inst Brew,2006,112(3):246–254.
[24]Moradas-Ferreira P,Fernandes PA,Costa MJ. Yeast flocculation—the role of cell wall proteins. Colloids Surf B Biointerfaces,1994,2(1/3): 159–164.
[25]An WT,Yang D,Yin YH,et al. Mechanism analysis of yeast flocculation in the process of replicative aging. Microbiol China,2013,40(12): 2189–2195 (in Chinese).安文涛,杨迪,尹亚辉,等. 酵母菌复制衰老过程中絮凝机理分析. 微生物学通报,2013,40(12):2189–2195.
[26]Potter G. An examination of cell wall properties affecting brewing yeast flocculation[D]. Dalhousie:Dalhousie University,2014.
[27]Zhang M,Mao J,Huang GD,et al. Research progress in mechanism of alcohol tolerance of sake yeast. Sci Technol Food Ind,2012,33(20): 342–345,350 (in Chinese).张敏,毛健,黄桂东,等. 清酒酿酒酵母酒精耐受机理的研究进展. 食品工业科技,2012,33(20):342–345,350.
[28]Kwolek-Mirek M,Zadrag-Tecza R. Comparison of methods used for assessing the viability and vitality of yeast cells. FEMS Yeast Res,2014,14(7):1068–1079.
[29]Ji YY,Piao YZ,Yuan F,et al. Proteomics analysis ofSaccharomycescerevisiaewith differential generations. Food Sci Technol,2014,39(7): 26–30(in Chinese).季杨杨,朴永哲,袁方,等. 不同传代次数酿酒酵母细胞内蛋白质组学分析. 食品科技,2014,39(7):26–30.
[30]François J,Parrou JL. Reserve carbohydrates metabolism in the yeastSaccharomyces cerevisiae.FEMS Microbiol Rev,2001,25(1): 125–145.
[31]Herdeiro RS,Pereira MD,Panek AD,et al. Trehalose protectsSaccharomyces cerevisiaefrom lipid peroxidation during oxidative stress. Biochim Biophys Acta,2006,1760(3): 340–346.
[32]Xu WN. Study on the evaluation and mechanism of autolysis in lager brewer’s yeasts [D]. Wuxi:Jiangnan University,2014 (in Chinese).许维娜. 啤酒酵母自溶评价及机理研究[D]. 无锡:江南大学,2014.
[33]Hirasawa R,Yokoigawa K,Isobe Y,et al. Improving the freeze tolerance of bakers' yeast by loading with trehalose. Biosci Biotechnol Biochem,2001,65(3):522–526.
[34]Magalhães PJ,Barbosa AR,Gonçalves LM,et al.The impact of xanthohumol on a brewing yeast's viability,vitality and metabolite formation. J Inst Brew,2016,122(2): 363.
[35]Jermsuntiea W,Aki T,Toyoura R,et al. Purification and characterization of intracellular lipase from the polyunsaturated fatty acid-producing fungusMortierellaalliacea. New Biotechnol,2011,28(2):158–164.
[36]Treichel H,de Oliveira D,Mazutti MA,et al. A review on microbial lipases production. Food Bioprocess Technol,2010,3(2): 182–196.
[37]Zhang L,Li X,Luan N,et al. Study on the change of the activity of enzymes produced bySaccharomyces cerevisiaeof different generation. Liquor-Making Sci Technol,2012,(7): 53–55 (in Chinese).张利,李欣,栾南,等. 不同代数酿酒酵母发酵产酶酶活力变化研究. 酿酒科技,2012,(7): 53–55.
[38]Du J,Li HL,Li H,et al. Oxidative stress of wine yeasts under copper exposure. Sci Agric Sin,2011,44(2): 369–378 (in Chinese).杜君,李海兰,李慧,等. 铜对葡萄酒酿酒酵母的氧化胁迫机制. 中国农业科学,2011,44(2):369–378.
[39]Yin YH,An WT,Dong L,et al. Changes of antioxidant enzyme activities in different algebraic ofSaccharomyces cerevisiae. Food Sci Technol,2013,38(8): 38–41,47 (in Chinese).尹亚辉,安文涛,董亮,等. 不同世代酿酒酵母胞内抗氧化酶活性变化. 食品科技,2013,38(8):38–41,47.
[40]de Freitas J,Wintz H,Kim JH,et al. Yeast,a model organism for iron and copper metabolism studies.Biometals,2003,16(1): 185–197.
[41]Zhang L,Wang S,Liu BX,et al. Changes in enzyme activity duringSaccharomyces cerevisiaeaging.Food Sci Technol,2012,37(7): 5–9 (in Chinese).张利,王硕,刘宝祥,等. 酿酒酵母老化过程中酶活力变化研究. 食品科技,2012,37(7): 5–9.
[42]Roberts GCK. Encyclopedia of Biophysics. Berlin Heidelberg: Springer,2013: 921–931.
[43]Yang TX,Wu KY,Chen JB,et al. Recent advances in fungal exopolysaccharide fermentation. Food Sci,2016,37(5): 265–270 (in Chinese).杨同香,吴孔阳,陈俊亮,等. 真菌发酵胞外多糖的研究进展. 食品科学,2016,37(5): 265–270.
[44]Pavlova K,Koleva L,Kratchanova M,et al.Production and characterization of an exopolysaccharide by yeast. World J Microbiol Biotechnol,2004,20(4): 435–439.
[45]An WT. Study on metabolism for yeast polysaccharide [D]. Dalian: Dalian Polytechnic University,2013 (in Chinese).安文涛. 酵母菌多糖的代谢研究[D]. 大连: 大连工业大学,2013.
