青藏高原两种典型植物根系抗拉特性与其微观结构的关系
2018-04-11朱海丽李国荣谢彬山李本峰
李 可, 朱海丽,2, 宋 路, 李国荣,2, 谢彬山, 李本峰
(1.青海大学 地质工程系, 西宁 810016; 2.青藏高原北缘新生代资源环境重点实验室, 西宁 810016)
青藏高原地区由于特殊的气候、地理条件,滑坡、崩塌等地质灾害频发,水土流失现象也越来越严重。随着水土保持观念的不断更新,国内外学者对植物根系护坡也更加重视,并发现在土壤中穿插植物根系,能有效地遏制土体的大规模变形[1-2]。在以往对青藏高原地区的灌木和草本植物根系护坡的研究中发现,植物根系对边坡浅层土体稳定、增加土壤强度等方面起到显著作用[3-5]。柠条锦鸡儿(CaraganakorshinskiiKom.)和草地早熟禾(PoapratensisL.)作为我国西北地区比较常见的灌木和草本植物,抗寒、抗旱、抗逆性较强。青海东部柠条锦鸡儿群落总盖度为80%,主要由6种植物组成,群落垂直结构分两层,即灌木和底下草本植物。灌木层优势种为柠条锦鸡儿,平均高度为112 cm,分盖度为55%,生物量达到(1 530.99±809.23) g/m2[6],此外西藏沙棘(HippophaethibetanaSchlechtend.)也占有明显优势;草本层优势种为赖草(LeymussecalinusTzvel.),次优势种为草木樨状黄芪(AstragalusmelilotoidesPall.)、油蒿(ArtemisiadesertorumSpreng. Syst. Veg.)、狗尾草[Setariaviridis(L.) Beauv.]等植物;草地早熟禾群落总盖度为85%,优势种为草地早熟禾,平均高度为54.9 cm,分盖度为60%,生物量达到368.64~633.95 g/m2[7],次优势种为垂穗披碱草(ElymusnutansGriseb.)、油蒿(ArtemisiadesertorumSpreng. Syst. Veg.)、针茅(StipacapillataLinn.)、多裂委陵菜(PotentillamultifidaL.)等植物。熊燕梅[8]、胡夏嵩[9]、朱海丽[10]、李臻[11]等学者对该类植物根系的固土作用研究主要集中在植物根系抗拉特性及不同植物种根系抗拉特性的比较等方面,认为植物根系的抗拉特性主要受根系种类和根系直径的影响,根系抗拉特性与根径之间呈幂函数或指数函数关系。近几年的研究已经证实,由于植物根系微观结构的复杂性与特殊性,植物微观结构是影响木材力学性能的决定因素[12-14]。朱海丽等[15]通过对灌木根系进行单根拉伸以及根系解剖等试验,指出根系韧皮纤维、木纤维的百分含量及周皮的木质化程度显著影响根系力学强度。蒋坤云等[16-17]进行了乔木根系拉伸和根系分子离析试验,定量分析出根系木纤维含量与根系抗拉特性呈正相关关系。郭维俊等[18]研究认为小麦根系承载能力取决于机械组织厚度、维管束数量等;唐朝生等[19]提出纤维表面形态学特征及接触面之间力的产生和传递是纤维加筋土的主要力学机理。以往的研究主要针对不同种植物根系的抗拉力、抗拉强度、根系受力应力—应变特征等开展了相关研究,而对在不同速率、不同标距下植物根系固土的力学特性及其与微观结构间的关系研究还较少[20-26]。
因此本文通过对西宁地区草地早熟禾和柠条锦鸡儿两种优势植物根系的抗拉特性进行测定;重点分析不同拉伸速率和不同标距对植物单根抗拉力和抗拉强度的影响;此外通过对根系拉断断面进行电镜扫描,分析两种植物根系受力破坏特点,揭示根系受力变化的影响因素,为植物根系固坡机理的深入研究以及青藏高原区固坡植被的选择提供理论依据。
1 研究区自然概况
研究区位于青海省西宁市城北区青海大学后山顾家岭沟,地处青海东北部湟水谷地中部西宁盆地,海拔约2 275 m,属高原寒冷干旱—半干旱气候。具有寒长暑短、昼夜温差大等特点。年平均气温为5.9℃,年均降水量为345 mm,集中在6—9月份,占全年降水量的60%左右,且多以暴雨和阵雨形式出现[9]。试验区属于低山丘陵区,黄土边坡坡度较大,冲沟发育,冲蚀沟深达十米至几十米,25°以上的坡地占该地面积的47.2%,前缘分布有冲积扇或坡积裙,水土流失严重。每逢暴雨季节,易诱发滑坡、崩塌、泥石流等不同规模的地质灾害,给研究区周边居民的生命财产安全构成了一定威胁。
2 材料与方法
2.1 根系材料
选取研究区耐寒、抗旱的优势植物种柠条锦鸡儿和草地早熟禾两种根系发育典型的植被作为研究对象[11]。采用挖掘剖面壁法挖取植株根系,草地早熟禾挖掘深度为0.4~0.8 m,柠条锦鸡儿挖掘深度为1.0~1.5 m。在开挖过程中,记录并统计植株根系的主根和侧根的分布、延伸方向、分支情况及根径、根量等。然后将挖取的植株地下部分用毛刷将其表面粘附的土体剥离,用保鲜膜包裹起来,再将整个植株植入试验盆中带回实验室。在试验过程中,用环刀取根系周围原状土,测定土体天然密度,并用铝盒取一定量土体带回实验室应用CS101-EBN烘箱测定土体含水率。试验区土体基本物理性质指标测定结果见表1。

表1 试验区土体的基本物理性质指标测定结果
2.2 试验内容与方法
2.2.1单根拉伸试验将取回的植物根系放入LRHS-150-Ⅲ恒湿恒温培养箱保证其根系新鲜度,选取表皮完好、根径通直均匀的根系作为试验材料。根据试验需要将截取根长分别为60,80,100 mm的根段各60~80段。试验选用RWT-6010微机控制电子式万能试验机,参照试验标准[27],分别设定40,60,80 mm共3个标距,3,5,10 mm/min共3种不同速率对两种植物根系进行拉伸试验。试验开始前,将选取的植物根系,剪切至试验要求长度,并将剪断后备用的根段试样用湿毛巾包裹,以减少根段中水分蒸发。在进行单根拉伸试验时,将不同长度的根段夹持在拉伸仪的上、下夹具间,按不同的设计要求进行拉伸试验。试验开始后,按规定的拉伸速率,使受测根段试样处于受拉状态,直至根段断裂,受测根段试样断裂时的抗拉力即为单根最大抗拉力。每组试验重复45~55次。由于在拉伸试验过程中,拉伸夹具为刚性材料,在拉伸过程中存在柔性材料根段被夹具夹断的现象,因此为了提高试验的成功率,即植物根段受到拉伸作用力而产生断裂,而非夹具作用断裂,本项试验采用在夹具两端黏贴薄海绵以增大根系与夹具间的摩擦,并选取试验中受拉根段在夹具中间部位断裂的试验数据作为有效数据。根系力学参数的计算方法[10]为:

(1)

(2)

(3)
式中:P为根系抗拉强度(MPa);Fmax为根系最大抗拉力;F为实时拉力(N);σ为应力(MPa);D为根系平均直径(mm);ε为纵向线应变即延伸率;ΔL为根系延伸时的延长量(mm);L为夹具两端受到拉伸作用的根段长度,即标距(mm)。
2.2.2断裂根系微观结构首先选取表面完好且有活性的柠条锦鸡儿和草地早熟禾两种相同根径的根段进行单根拉伸试验,挑选拉伸标距为40 mm时,拉伸速率为3,5,10 mm/min条件下拉伸断裂位置居中的3段受拉根系作为试验材料,并做好编号和标签,用刀片在不同编号的根系断口处截取横、纵断面根段,并将截取下的小根段固定在膜片上。采用置换干燥法取代根系中的水分,再进行抽真空干燥。在真空条件下进行喷金处理镀膜后,采用JSM-6610 LV型电子显微镜进行取相,并通过Image-Proplus图样分析系统观测两种植物根系微观断面特征以及根系周皮受拉变形量和根系内部管孔直径的变化。
3 结果与分析
3.1 优势植物抗拉特性与根径关系
3.1.1单根抗拉力与根径的关系通过对研究区两种优势植物种柠条锦鸡儿和草地早熟禾进行单根拉伸试验,结果如表2—3所示,最终试验成功测定柠条锦鸡儿根段试样157个,成功率60.38%;草地早熟禾根段试样159个,成功率62.35%。试验成功的柠条锦鸡儿根径平均值为(0.76±0.14) mm,单根平均最大抗拉力为(25.29±9.5) N;草地早熟禾根径平均值为(0.20±0.05) mm,单根平均最大抗拉力为(1.85±0.41) N。2种植物的根系具有不同的抗拉力,灌木植物柠条锦鸡儿的抗拉力明显高于草本植物草地早熟禾。以拉伸速率为5 mm/min拉伸标距为40 mm为例,见图1对2种植物的单根抗拉力与根径之间的关系进行拟合分析表明:单根的抗拉力随根径的增大而增大,且2种植物的单根抗拉力与根径之间均存在幂函数关系。在其他拉伸速率和拉伸标距下试验结果也同样具有此规律。

表2 试验区柠条锦鸡儿单根拉伸试验统计结果
注:相同小写字母表示差异性不显著,有一个相同小写字母表示差异性<0.05,不同小写字母表示差异性<0.01,下表同。

表3 试验区草地早熟禾单根拉伸试验统计结果

图1 试验区2种植物单根最大抗拉力与根径的关系
3.1.2单根抗拉强度与根径的关系由单根抗拉强度与根径之间的拟合方程可知,2种植物单根抗拉强度与根径之间均呈幂函数或指数函数关系。如表2—3所示,柠条锦鸡儿的单根根系抗拉强度范围为36~84 MPa,平均值为(52.15±16.96) MPa,草地早熟禾的单根根系抗拉强度范围为26~140 MPa,平均值为(64.43±20.25) MPa。对试验区2种植物单根抗拉试验以5 mm/min标距40 mm为例进行分析得出,随着根径的增大,两种植物根系的单根抗拉强度逐渐减小,其关系可用乘幂函数公式表示(图2)。

图2 试验区2种植物单根抗拉强度与根径的关系
3.2 不同标距下单根抗拉力及抗拉强度
通过实地考察发现,研究区河流近岸由于水流冲蚀作用下河岸下部砂砾石层蚀退,从而使上部根—土复合体层在自重的作用下产生拉张裂隙,裂缝中穿插的植物根系形成了一张根系网,充分发挥了根系的抗拉作用,从而减缓了边岸滑塌。根据野外测量临界崩塌块体裂隙宽度约宽7 cm,基于根—土复合体层裂隙宽度,本次单根拉伸试验设置拉伸标距为4,6,8 cm,在拉伸速率为5 mm/min下进行拉伸试验。
试验研究发现,以草地早熟禾在标距为40 mm,平均根径为0.21 mm为例,其单根平均最大抗拉力达到2.47 N,较相同条件下标距为60 mm和80 mm的平均最大抗拉力1.27 N和1.18 N,增加了约1倍;以柠条锦鸡儿在标距为80 mm,平均根径为0.68 mm为例,其单根平均最大抗拉力达到31.65 N,较相同条件下标距为40 mm和60 mm的平均最大抗拉力18.59 N和15.54 N,增加了约1倍(图3)。同时由表2—3可看出,在相同速率下草地早熟禾根系平均抗拉力变化趋势依次为标距40 mm>标距60 mm>标距80 mm;柠条锦鸡儿根系平均抗拉力变化趋势为标距80 mm>标距60 mm>标距40 mm。由此可以看得出:标距40 mm为草地早熟禾发挥最大抗拉力的最佳的标距;标距80 mm为柠条锦鸡儿根系发挥最大抗拉力的最佳标距。造成这种情况的原因可能是两种植物根系的微观组成结构不同,而不同的结构在不同标距下才能发挥各自的最大作用,所以两种植物根系的最佳标距不同。
由图4可知,随着根径的增加,抗拉强度降低,根径越细,抗拉强度越大。根径在0.10~0.15 mm,草地早熟禾标距为60 mm时其抗拉强度最大,随着根径的增大,标距为60 mm的植物根系的抗拉强度下降幅度最大,且在根径超过0.2 mm后,标距为60 mm的根系抗拉强度小于标距为40,80 mm的根系抗拉强度;柠条锦鸡儿在标距为80 mm时根系抗拉强度最大,标距为60 mm时最小,标距40 mm的抗拉强度介于两者之间。经SPSS 21.0比较均值中的单因素方差检验,分别定量分析了两种优势植物根系的抗拉力、抗拉强度在不同拉伸标距、拉伸速率下的差异显著性,结果见表4,除柠条锦鸡儿根系抗拉力在不同拉伸标距下差异性不显著外,其余植物种根系抗拉特性与拉伸标距均呈现显著的差异性。
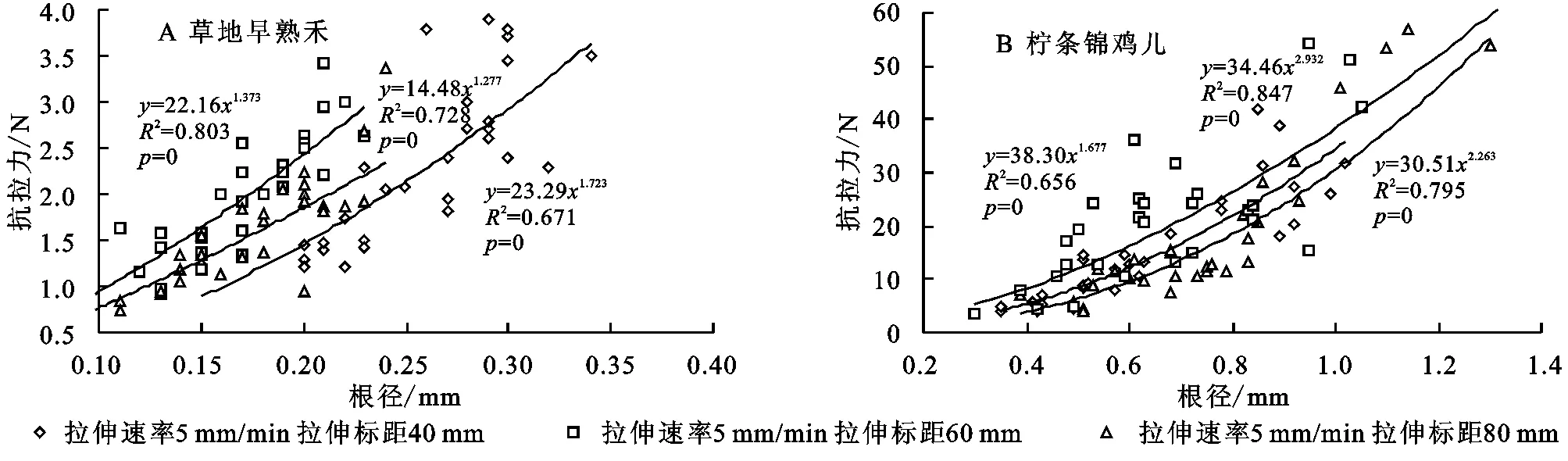
图3 试验区2种植物在拉伸速率5 mm/min不同标距下的根径与抗拉力关系曲线

图4 试验区2种植物在拉伸速率5 mm/min不同标距下的根径与抗拉强度关系曲线
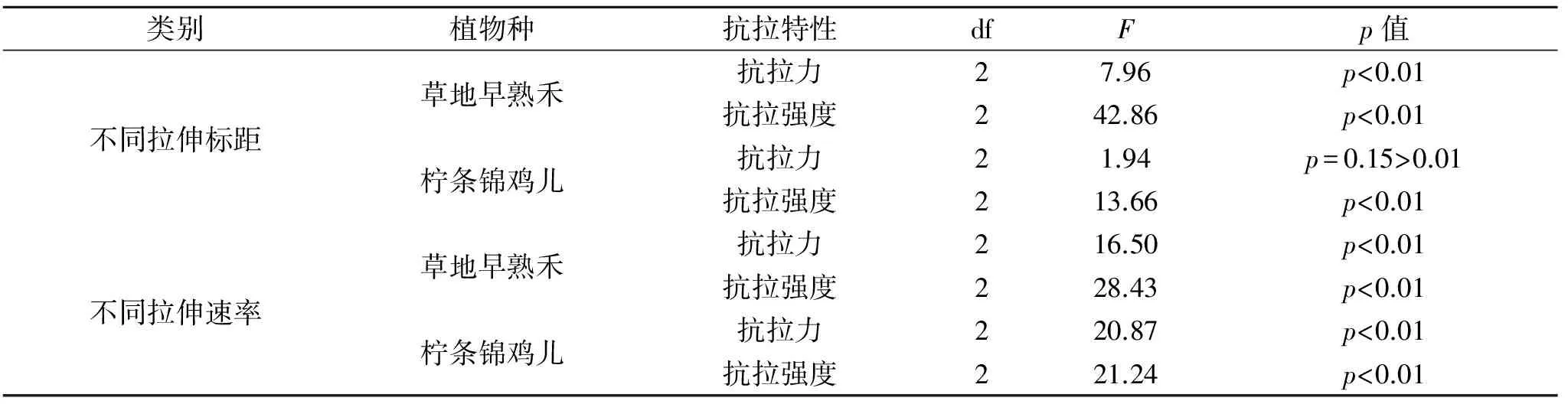
表4 不同因素下两种植物根系抗拉特性差异的单因素方差分析结果
由此可以看出:在相同根径的情况下,标距60 mm是草地早熟禾根系发挥最佳抗拉强度的标距;而柠条锦鸡儿在试验设定的不同标距下,其根系抗拉强度差异不明显。由此可以看出两种植物中草地早熟禾须根丰富,尤其是根径小于0.5 mm的活性根对土体具有较强的加筋作用,能够增强土体的抗剪、抗冲性能。但在不同标距下,两种植物根系的抗拉强度并没有随标距的变化呈规律性增加或减少。这与之前周朔[21]对乔木油松的抗拉强度研究中提到的相同根径的根系抗拉强度随标距的减小而减小的关系不是很相符。这种差异性很有可能是由于草本、灌木或乔木植物的根系微观组成有较大的差异,另外与不同地域土壤、气候、水分存在显著差异性有关,但具体原因有待深入分析。
3.3 不同速率下单根抗拉力及抗拉强度
通过对两种优势植物根系在拉伸标距为40 mm,拉伸速率分别为3,5,10 mm/min进行单根拉伸试验,分析不同拉伸速率对两种植被根系的抗拉力学特性的影响。由图5可知,随着根径的递增,两种植物根系的抗拉力均呈现增大的趋势。草地早熟禾根系在拉伸速率为10 mm/min时,其抗拉力增加幅度最大;对于根径<0.30 mm的根系抗拉力,随着拉伸速率的减小而减小。柠条锦鸡儿根系在拉伸速率为5 mm/min时,其抗拉力增加幅度最大;当根径>1.11 mm时,拉伸速率为5 mm/min下的根系抗拉力逐渐接近并超过拉伸速率为10 mm/min下的抗拉力。可以看出,在拉伸试验中,草地早熟禾和柠条锦鸡儿两种优势植物根系的平均抗拉力随拉伸速率的增大而减小。研究表明当研究区土坡发生崩塌时,其破坏速率越小,植物根系越能充分发挥其抗拉特性。

图5 试验区2种植物在不同速率下的根径与抗拉力关系曲线
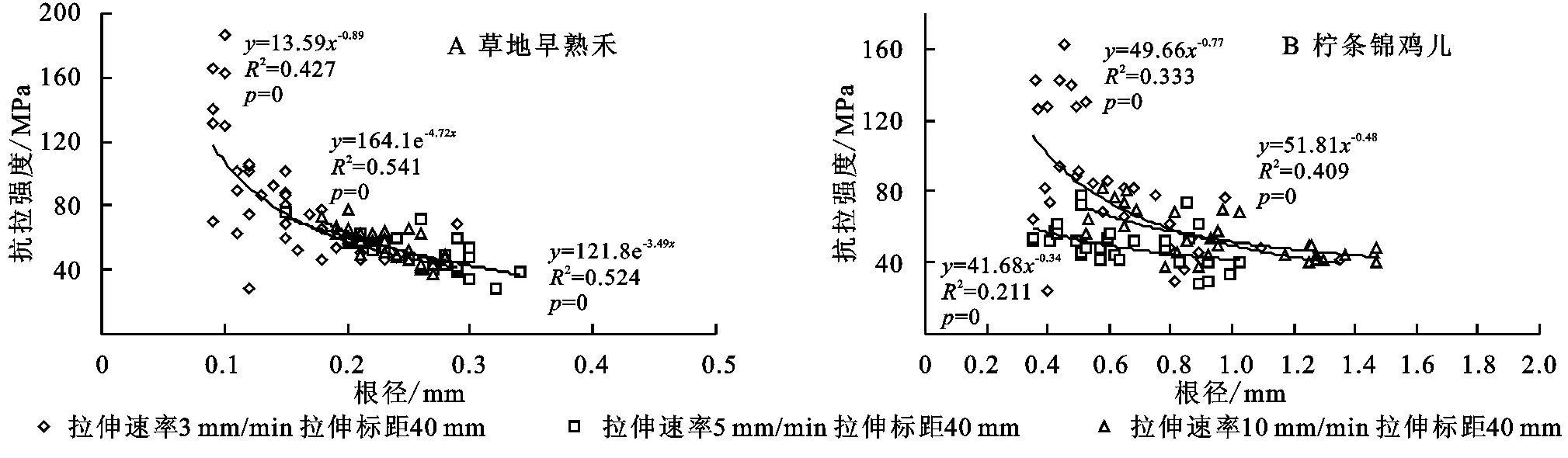
图6 试验区2种植物在不同速率下的根径与抗拉强度关系曲线
由图5可知,草地早熟禾和柠条锦鸡儿在拉伸标距为40 mm条件下,随着植物根径的增大,植物根系抗拉力随着拉伸速率的增大而显著增大,且同一根径下拉伸速率愈大,根系抗拉力愈小。由图6可知,当柠条锦鸡儿根径在0.3~0.5 mm、草地早熟禾根径在0.1~0.15 mm时,根系抗拉强度在3 mm/min时最大,且随着根径的增大,草地早熟禾和柠条锦鸡儿根系抗拉强度随着拉伸速率的增大而减小,且拉伸速率为3 mm/min时,根系抗拉强度下降幅度最大。草地早熟禾在根径>0.27 mm后,拉伸速率为5 mm/min下的根系抗拉强度趋近于拉伸速率为10 mm/min的根系抗拉强度;柠条锦鸡儿在根径>0.66 mm后,拉伸速率为3 mm/min的抗拉强度小于拉伸速率为10 mm/min的。所以在土体的快速崩塌过程中,较细的根系能发挥其抗拉力较高的特点,能有效提高河岸的稳定性。两种优势植物根系的抗拉力、抗拉强度在不同拉伸速率下差异显著性的方差分析结果见表4。
3.4 应力-应变曲线
通过对试验区两种植物单根拉伸应力—应变曲线分析可知,无论是草本植物还是灌木植物,由于其根系结构组成不同,根系拉伸的应力—应变特征也不同。试验区两种植物单根拉伸应力—应变曲线表现为弹塑性材料特性。分别由根径为0.22 mm的草地早熟禾和根径为0.69 mm的柠条锦鸡儿单根拉伸应力—应变曲线,由图7—8可知,在相同根径下,不同速率、不同标距下其应力—应变曲线也是不同的。两种植物根系在受拉的初始阶段,应力与应变呈比例增长,表现为线性关系;随着进一步拉伸,当应力超过弹性极限之后,应变较应力增长更快,无残余变形,表现为非线性弹性变形,此阶段的应力增量相对较小,抗拉应力—应变曲线的斜率也较为平缓,当达到破坏抗拉力的时候,根系断裂破坏。
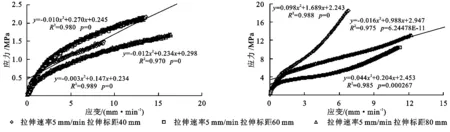
图7 试验区2种植物在不同标距下的应力-应变曲线的关系
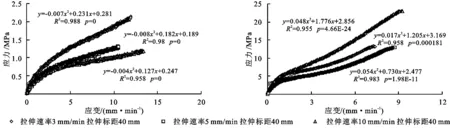
图8 试验区2种植物在不同速率下的应力-应变曲线的关系
从图8看出,草地早熟禾单根在受力时,表现有较大的弹性;当荷载超过弹性极限,拉力继续增加时,应力与应变关系逐渐偏离直线,呈上凸曲线,斜率渐减,表现出非线性弹性特征;柠条锦鸡儿单根受力时,也表现有较大的弹性;当荷载超过弹性极限,拉力继续增加时,应力与应变关系逐渐偏离直线,呈略微下凹曲线,这是由于柠条锦鸡儿在拉伸过程中首先是韧皮部受力,然后是其木质部受力的原因所导致的。
根系在拉伸过程中其延伸率是衡量根系拉断之前总的变形能力对坡体变形而言的一个重要指标,其指根系在单向拉伸下断裂前所达到的最大延伸率,包括弹性应变和塑性应变。当边坡土体间出现滑动裂缝时,延伸率较大的根系有利于把根系所受的拉力向土体深层传递,且延伸率较大的根系在受拉过程中根系形变量也较大,有利于使根系所受的拉力向根系侧摩阻转化而使根系本身所受的内力减小,这样有利于提高根系固土能力。通过对试验区草本植物和灌木植物根系的拉伸应力—应变分析知:草地早熟禾在相同根径、相同拉伸速率不同拉伸标距下的延伸率由大到小为:标距40 mm>标距60 mm>标距80 mm;在相同根径、相同拉伸标距不同拉伸速率下的延伸率由大到小为:3 mm/min>5 mm/min>10 mm/min;柠条锦鸡儿,在相同根径、相同拉伸速率不同拉伸标距下的延伸率由大到小为:标距40 mm>标距60 mm>标距80 mm;在相同根径、相同拉伸标距不同拉伸速率下的延伸率由大到小为:3 mm/min>5 mm/min>10 mm/min。可以看出两种植物在相同根径、拉伸速率为5 mm/min,拉伸标距为40 mm时的延伸率最大,随着标距的增加延伸率呈下降趋势;在相同根径、拉伸标距为40 mm,拉伸速率为3 mm/min时两种植物根系的延伸率最大,随着拉伸速率的增加延伸率也呈下降趋势。由表2—3可知,草本植物草地早熟禾的平均延伸率(24%)高于灌木植物柠条锦鸡儿(16.80%),由此可以看出草本植物根系所具有的较高的抗拉强度、较大的延伸性能和弹塑性的特征,对于提高试验区坡体浅层土体的强度和粘聚力,同时对抵抗坡体变形都具有重要作用;灌木植物以其较长的根系和根系所具有的弹塑性应力—应变特征对强化根—土复合体的抗剪切能力、锚固较深层的土体及提高土体粘聚力也都具有重要意义。
通过进一步观察分析根系的拉伸过程中的应力—应变曲线的特征,比较两种植物根系无论在不同标距还是不同速率下的应力—应变曲线发现,根系的应力—应变曲线整体都为单峰曲线,特别是草地早熟禾较细根系的曲线表现出明显的非光滑性的特征,这与钢筋受拉的应力—应变曲线的特征明显不同,说明草本植物草地早熟禾的根系比灌木植物柠条锦鸡儿的根系在拉伸过程中对外界拉力具有较强的缓冲能力。
3.5 根系拉伸断面微观特征及其与单根抗拉特性的关系
3.5.1根系拉伸断面微观特征对2种优势植物根系的显微结构进行观察后发现,草地早熟禾根系结构较柠条锦鸡儿根系有较大的差异。草地早熟禾根系的木质化程度较低,由表皮、皮层和中柱3部分组成,早熟禾根系的表皮较薄,皮层中的管孔受挤压大多呈现多边形,中柱中管孔直径相对较小,密度较高,且管孔呈团状,大小较为统一,分布在整个次生木质部中。柠条锦鸡儿根系木质化程度较高,主要由周皮、韧皮部、次生韧皮部以及中柱等组成。根系表皮较薄,次生韧皮部和次生木质部占横截面总面积的比例较大;中柱在根系横截面中所占的面积最大,由木质部、韧皮部等组成;管孔类型为复管孔,分布在整个次生木质部中;木纤维在根系横截面所占的比例较大[28]。
对比不同拉伸速率下发生断裂的植物根系横断面微观结构图,可知草地早熟禾根系在拉伸速率为3 mm/min时,周皮与管孔壁剥离,周皮断裂并发生塑性变形产生一定的延展现象,也有弹性变形。在断口处周皮形如褶皱,次生木质部在周皮断裂后断口不整齐见图9A,管孔变形大,撕裂程度高;当拉伸速率为5 mm/min时次生木质部断裂口较整齐,管孔形变不大。周皮撕裂程度较低,且与管孔连接较好。周皮产生一定的延展现象并形成褶皱,有明显二次断裂痕迹见图9B;当拉伸速率为10 mm/min时次生木质部断裂整齐,周皮与管孔连接完整,无剥离。周皮没有延展没有明显二次断裂痕迹,次生木质部断裂管孔没有明显被撕裂,管孔形状保持较好见图9C。柠条锦鸡儿根系在拉伸速率为3 mm/min时,根系断面呈现参差不齐状,管孔受到挤压变形较大,中柱中木纤维在拉伸作用下产生延展,发生二次断裂,撕裂程度高;当拉伸速率为5 mm/min时木质部断裂面整齐,皮层与中柱剥离,管孔变形不大,发生延展的木纤维的数量相比拉伸速率为3 mm/min时较少;当拉伸速率为10 mm/min时,木质部断裂面整齐,管孔形状保持较好,木纤维延展的距离相对较短(图10)。

图9 草地早熟禾根系在不同拉伸速率下的断面微观结构

图10 柠条锦鸡儿根系在不同拉伸速率下的断面微观结构
在观察不同速率下根系受拉横断面管孔变化情况时,分别测定了不同拉伸速率下受拉横断面电镜扫描照片中管孔短轴的平均直径,见表5。可以看出拉伸速率越大,根段受拉断裂面越平整,平均管孔短轴直径越小。表明在较慢拉伸速率下,根系中的纤维束能够逐渐发挥自身抗拉特性并直到拉断,因此断面参差不齐,且管孔变形严重。随着拉伸速率的增大,根系的弹性变形没有完成就进入了塑性变形,根系大多为脆性断裂,因此断面平整,管孔的直径变化也相对较小。
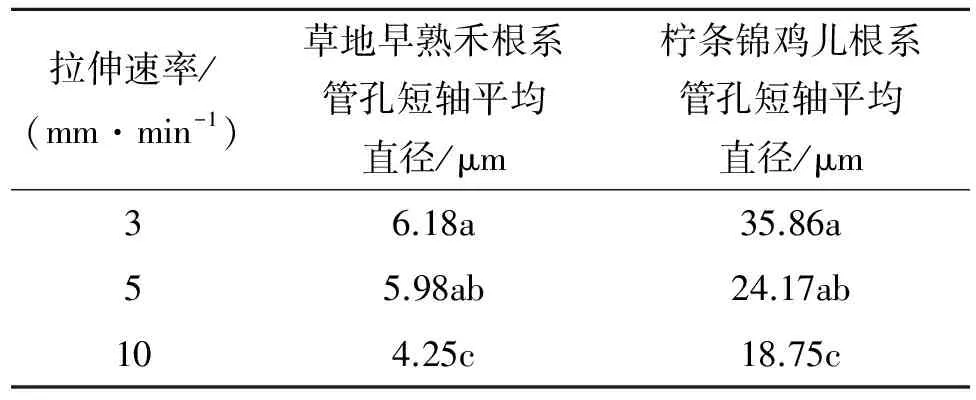
表5 根系拉伸断面管孔平均直径
图11为草地早熟禾根段分别在3 mm/min,5 mm/min和10 mm/min这3种拉伸速率下的纵断面电镜扫描照片。当拉伸速率为3 mm/min时,受拉后的植物根系周皮产生拉裂裂缝,撕裂程度严重,周皮表面有大量断裂的纤维丝。随着拉伸速率的增大,周皮表面的拉裂裂缝数量逐渐减少且裂缝宽度也愈不明显。表明根系在较慢的拉伸速率下能通过纤维丝的抗拉作用来减缓根系断裂,纤维束呈现渐进式破坏。观察柠条锦鸡儿根系的纵断面微观图片(图12),可知根系纵断面的拉裂裂缝数量是随着拉伸速率的增大而减少,拉裂程度也逐渐减弱。由于柠条锦鸡儿根系木质化程度较高,所以周皮表面没有断裂的纤维丝。

图11不同速率下草地早熟禾根系纵断面特征

图12 不同速率下柠条锦鸡儿根系纵断面特征
3.5.2两种植物的根系微观结构与抗拉特性的相关性在试验中发现,两种植物根系在速率为3 mm/min下大多为弹塑性断裂,尤其是柠条锦鸡儿一般都是外层韧皮部先被拉裂,裂纹呈45°方向交叉分布然后木质部依次被撕裂。随着速率的加快逐渐转为脆性断裂,断口平滑,由于根系的弹性变形没有完全进行完就进入了塑形变形,力学性质没有得到充分的发挥,导致在相同根径、相同标距的前提下抗拉力和抗拉强度随着速率的加快都有所降低。通过对早熟禾(图9)与柠条锦鸡儿(图10)根系微观结构观察发现,柠条锦鸡儿根系木质化程度高于早熟禾根系,表皮厚度也要明显大于早熟禾。由此可见,柠条的次生微管组织发达,使得根系的抗拉特性较强,草地早熟禾的次生微管组织发展相对较弱且细胞排列疏松,使得根系的抗拉特性较弱[11]。在对试验拉断根系横断面微观结构观察中发现,在不同的拉伸速率下柠条锦鸡儿根系横断面都有大量参差不齐的纤维状物质,即柠条锦鸡儿根系木纤维在根系横截面所占的比例要大于早熟禾。随着拉伸速率的增大,草地早熟禾根系周皮的延伸率逐渐降低;柠条锦鸡儿根系中木纤维的型变量也相应的减小(图9—10)。
通过分析试验数据以及观察2种植物根系横断面微观结构图得出,拉伸速率在较小的情况下,植物根系自身的周皮及木纤维会随着根系的受拉发挥一定抗拉作用,影响根系的抗拉特性。该结论也符合蒋坤云在灌木、草木植物根系抗拉特性中的研究结果[16-17]。
4 结 论
(1) 单根拉伸试验表明,两种植物根系抗拉力随着根径的增大呈幂函数增长;而根系抗拉强度随根径的增大而减小,草地早熟禾以指数函数形式递减、柠条锦鸡儿以幂函数形式递减。试验研究中,草地早熟禾根径<0.25 mm的根系与柠条锦鸡儿根径<0.5 mm的根系具有较大的抗拉强度。
(2) 在不同拉伸标距下的拉伸试验中,草地早熟禾发挥最大抗拉力的标距为40 mm;柠条锦鸡儿发挥最大抗拉力的标距为80 mm。根径相同条件下,草地早熟禾标距为60 mm时抗拉强度最大,且随着根径的增大,3种标距下单根的抗拉强度变化幅度逐渐减小;而柠条锦鸡儿在标距为80 mm时其抗拉强度最大。柠条锦鸡儿根系的抗拉力在不同标距下差异不显著。
(3) 在不同拉伸速率下的拉伸试验中,两种植物根系抗拉力和抗拉强度在拉伸速率为3 mm/min时最大。随着拉伸速率的加快根系从弹塑性断裂逐渐转为脆性断裂,断口平滑,由于根系的弹性变形没有完全进行完就进入了塑形变形,力学性质没有得到充分的发挥,导致在相同根径、相同标距的前提下抗拉力和抗拉强度随着速率的加快都有所降低。两种植物根系的抗拉特性在不同拉伸速率下均呈现显著差异性。
(4) 对两种植物根系拉伸断面电镜扫描观察断面SEM照片可知,不同拉伸速率、拉伸标距下两种植物根系周皮的延伸程度、撕裂程度以及管孔的变化均不相同。总体来说拉伸速率越小周皮延伸长度越大、撕裂程度越高、管孔变形越严重。同时随着植物根径变化、根系木质化程度加剧以及根系木纤维增加,植物根系的抗拉特性也会有相应的变化。
参考文献:
[1]Wu T H, McKinnell W P Ⅲ, Swanston D N. Strength of tree roots and landslides on Prince of Wales Island, Alaska[J]. Canadian Geotech Journal, 1979,16(1):19-33.
[2]Waldron L J, Dakessian S. Soil reinforcement by roots: Calculation of increased soil shear strength from root properties[J]. Soil Science, 1981,132(6):427-435.
[3]赵玉娇,胡夏嵩,刘昌义,等.寒旱环境植物根系增强边坡土体抗剪强度试验研究[J].水土保持研究,2016,23(4):212-220.
[4]栗岳洲,付江涛,余冬梅,等.寒旱环境盐生植物根系固土护坡力学效应及其最优含根量探讨[J].岩石力学与工程学报,2015,34(7):1370-1383.
[5]乔娜,余芹芹,卢海静,等.寒旱环境植物护坡力学效应与根系化学成分响应[J].水土保持研究,2012,19(3):108-113.
[6]李刚,赵祥,张宾宾,等.不同株高的柠条生物量分配格局及其估测模型构建[J].草地学报,2014,22(4):769-775.
[7]贺玺琳,杨慧茹,马玉寿,等.青海草地早熟禾植物量生长动态研究[J].青海畜牧兽医杂志,2016,46(1):13-16.
[8]熊燕梅,夏汉平,李志安,等.植物根系固坡抗蚀的效应与机理研究进展[J].应用生态学报.2007,18(4):895-904.
[9]胡夏嵩,李国荣,朱海丽,等.寒旱环境灌木植物根—土相互作用及其护坡力学效应[J].岩石力学与工程学报,2009,28(3):613-620.
[10]朱海丽,胡夏嵩,毛小青,等.青藏高原黄土区护坡灌木植物根系力学特性研究[J].岩石力学与工程学报,2008,27(2):3445-3452.
[11]李臻,余芹芹,杨占风,等.西宁盆地两种灌木植物原位拉拔试验及其护坡效应[J].水土保持研究,2011,18(3):206-209.
[12]Fung Y C. Biomechanics: Mechanical Properties of Living Tissues[M]. Germany: Springer-Verlag, 1993.
[13]李有芳,刘静,张欣,等.4种植物生长旺盛期根系易受损的外力类型研究[J].水土保持学报,2016,30(6):339-344.
时间一分一秒地过去,激荡在内心的愤恨盛怒已消失殆尽,恐慌焦虑悄然爬上心头。昨天老四曾嘱咐我们要善于发现对手的弱点,但皮特除了缺乏讨喜的品质外,没有任何弱点。他够高够强壮,块头大,但又没有大到使行动迟缓;他对别人的弱点有很好的判断力;他生性恶毒,肯定不会对我有半点慈悲心。我本想理直气壮地告诉他不要小瞧我,可这分明是个谎言。皮特猜得一点没错,我真是一点打斗能力都没有。
[14]Maeglin R R. National variation of tissue proportions and vessel and fiber length in mature northern red oak[J]. Silvae Genetica, 1976,25(3/4):122-126.
[15]朱海丽,胡夏嵩,毛小青,等.护坡植物根系力学特性与其解剖结构关系[J].农业工程学报,2009,25(5):40-46.
[16]蒋坤云,陈丽华,盖小刚,等.华北护坡阔叶树种根系抗拉性能与其微观结构的关系[J].农业工程学报,2013,29(3):115-123.
[17]蒋坤云,陈丽华,杨苑君,等.华北油松、落叶松根系抗拉强度与其微观结构的相关性研究[J].水土保持学报,2014,27(2):8-12.
[18]郭维俊,黄高宝,王芬娥,等.小麦根系力学性能及微观结构研究[J].农业机械学报,2010,41(1):92-95.
[19]唐朝生,施斌,顾凯.纤维加筋土中筋/土界面相互作用的微观研究[J].工程地质学报,2011,19(4):610-615.
[21]周朔.林木根系拉伸力学特性研究[D].北京:北京林业大学,2011.
[22]胡敏,李为萍,史海滨,等.布根方式及根系径级对根土复合体抗剪性能的影响[J].水土保持通报,2012,32(1):42-44.
[23]吕春娟,陈丽华,周硕,等.不同乔木根系的抗拉力学特性[J].农业工程学报,2011,27(1):329-335.
[24]Bischetti G B, Chiaradia E A, Simonato T, et al. Root strength and root area ratio of forest species in Lombardy(Northern Italy)[J]. Plant and Soil, 2005,278(1):11-22.
[25]陆桂红,欧国强,杨顺,等.泥石流源区典型草本植物根系抗拉试验[J].山地学报,2014,32(6):725-731.
[26]张兴玲,胡夏嵩,毛小青,等.青藏高原东北部黄土区护坡灌木柠条锦鸡儿根系拉拔摩擦试验研究[J].岩石力学与工程学报,2011,30(S2):3739-3745.
[27]中华人民共和国国家质量监督检验检疫总局,中国国家标准化管理委员会.金属材料 拉伸试验 第1部分:室温试验方法GB/T288.1—2010[S].北京:中国标准出版社,2010.
[28]陈世鐄.中国北方草地植物根系[M].长春:吉林大学出版社,2001.
