模拟极端石漠化土壤条件对降香黄檀根系空间分布的影响
2018-04-11倪周游杨振德杨昌儒玉舒中
倪周游,杨振德,杨昌儒,小 梅,玉舒中
喀斯特石漠化是指在亚热带脆弱的喀斯特环境背景下,受人类不合理社会经济活动的干扰破坏,造成土壤严重侵蚀,基岩大面积裸露,土地生产力严重下降,地表出现类似荒漠景观的土地退化过程[1]。喀斯特环境问题已成为当今最严重的环境地质问题[2],不断扩大的土地石漠化对当地生态环境造成严重破坏,正在吞噬当地民众的生存空间。当前,中国西南地区分布着集中连片的分布面积最广、发育最完整、生态环境最脆弱的喀斯特地貌。其中广西的岩溶面积以8.9万km2位列全国第二,石漠化土地面积以4.7 万 km2位列全国第三[3],分布涉及 10 个市的 76个县(市、区),严重影响农、林、牧业生产,制约社会经济的可持续发展[4]。目前,关于对于喀斯特石漠化地区的治理,主要还是依靠地表植被的恢复,其中比较好的经济植物有山葡萄、任豆、构树、金银花等[5]。
喀斯特石漠化地区生长的植物通常都具有耐旱、喜钙、石生等特点[6]。降香黄檀(Dalbergia odorifera)对土壤条件要求不高,干旱贫瘠的地方乃至陡坡、山脊、石边都可生长,具有很强的抗逆环境能力,适合在广西喀斯特石漠化地区生长[7]。杨振德等[8]研究表明,一年生降香黄檀幼苗的平均根系直径、总根体积和根尖数随着土壤含水量的减少呈降低趋势,总根长和总根表面积则先略有增加而后降低;刘震等[9]的研究表明,降香黄檀在50%石砾含量土壤条件下的生物量与全土壤体无显著差异,根冠比随着石砾含量的增加有所增大,表明降香黄檀对石漠化环境能够做出主动的生长适应性反应;杨昌儒等[10]的研究表明,适当的石砾栽培基质胁迫可以提升降香黄檀幼苗叶片的光合能力,降香黄檀叶片的气孔随着石砾含量的增加有增大和变长的趋势。为进一步阐明降香黄檀根系对石漠化环境的生长适应机制,本文探讨了降香黄檀幼苗在全石砾基质条件下根系生物量空间分布的情况。
1 材料与方法
1.1 幼苗种植
降香黄檀一年生幼苗(苗高 54~56 cm,地径0.7~0.9 cm)分别栽植于壤土和全石砾基质中。 全石砾基质为大小0.5 cm3的石灰岩碎石颗粒,壤土基质采自广西马山县 (107°41′~108°29′E,23°24′~24°2′N)石漠化地区土山赤红壤(pH 5.2,土壤容重1.26 g/cm3,土壤田间最大持水量 24.48%,全氮含量0.068%, 速效钾的含量为 0.006 g/kg, 速效磷含量0.014 g/kg)。每种基质栽植10株。试验在广西大学林学院苗圃进行,每天对幼苗滴灌一次,每次5 min,定期除去杂草。栽培容器为35 cm×30 cm的塑料桶,桶底部打直径2 cm的小孔以利于排水。幼苗正常生长12个月后进行测量。
1.2 根系取样
采用改良全根系挖掘法,分东、南、西、北4个方向,每个方向按照 0~5、5~10、10~15、15~20 cm 4 个垂直层次进行挖掘。为了保持根系完整性,小铲与手工相结合,将裸露的根系标号后剪下,将相同分组土层的装入袋内,带回实验室冲洗,用吸水纸吸干根系表面水分, 将根系按 d<0.5 mm、0.5≤d<1 mm、1 mm≤d<3 mm和d≥3 mm的标准分为4个等级。分别置入烘箱中在105℃杀青30 min后再于80℃烘干24 h至恒重,称量。
1.3 数据处理
用SPSS 17.0统计分析软件进行数据分析,采用独立样本T检验,比较降香黄檀各部分生物量差异、不同方向生物量差异以及不同径级降香黄檀根系生物量差异。比较降香黄檀根系生物量垂直分布差异用LSD最小显著性差异法检测数据间的显著性(P<0.05)。
2 结果与分析
2.1 降香黄檀各部分生物量
根系作为树木吸收外界物质能量的主要器官,根系的生理生态特征对林木地上部分生长起决定性作用,随着根系生物量的减少,地上部分的生长状况也受到影响。从表1可以看出,在壤土中降香黄檀的生物量要大于在石砾中降香黄檀的生物量;石砾中降香黄檀的根冠比大于壤土中降香黄檀的根冠比。

表1 降香黄檀各部分生物量
2.2 降香黄檀根系生物量的空间分布
2.2.1降香黄檀根系不同方向生物量 从表2可以看出,壤土中降香黄檀与石砾中降香黄檀的根系生物量在不同方向上具有不同的分布特征,壤土中降香黄檀的根系生物量表现为:南面>西面>东面>北面;石砾降香黄檀的根系生物量表现为:北面>西面>东面>南面。在东、南、西3个方向上,两个不同基质中的根系生物量差异显著(P<0.05)。

表2 降香黄檀根系各方向生物量分布
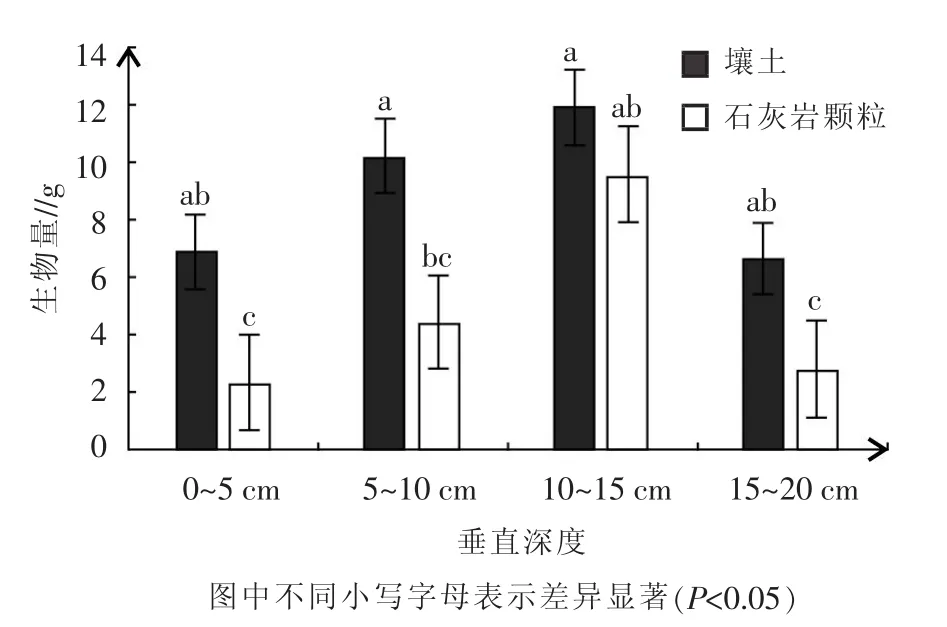
图1 不同土壤条件下降香黄檀根系生物量垂直分布
2.2.2降香黄檀根系生物量垂直分布 从图1可以看出,随土层深度的不断增加,两种不同基质中降香黄檀根系生物量均表现为先增大后减小的趋势,生物量的最大值都出现在10~15 cm的土层中。壤土中降香黄檀的根系生物量在不同深度的土层中无显著性差异,石砾中降香黄檀的根系生物量,10~15 cm处的根系生物量与0~5、15~20 cm处的根系生物量存在显著差异(P<0.05)。
2.2.3不同径级降香黄檀根系生物量 从表3可以看出,与其他径级生物量相比,直径d<0.5 mm根系生物量值分布最大,是决定降香黄檀根系生物量的主要因素,两个不同土壤基质中降香黄檀根系生物量的差别主要是由于直径d<3 mm的根系生物量的差别引起的。一般认为直径d<3 mm的根系是树木进行营养与水分吸收的主要器官[11],其生物量的差别直接影响了树木的生长发育。在两个不同的土壤基质中,直径d<3 mm的根系生物量存在显著差异,这是导致地上部分生长差异的重要原因。但通过进一步分析可以看出,石砾中的降香黄檀,直径d<0.5 mm的根系所占百分比要大于壤土中的降香黄檀。

表3 不同径级根系生物量(单位:g)
3 小结与讨论
根冠比是用来描述地下生物量与地上生物量之间关系的常用指标之一[12,13],生物量在不同器官的分配是植物对环境长期适应的结果,对植物的生长发育和繁殖有巨大作用。在石漠化土壤环境中,土壤中的有效资源在时间和空间上都是异质的,植物通过增大地下生物量的分配来获取有限的资源,寻觅有利生境,在刘震等[9]的研究中,根冠比随着石砾含量的增加有所增大,反映了降香黄檀对石漠化土壤环境的响应策略,与本研究的结果类似,在石砾中降香黄檀根冠比要大于壤土中降香黄檀的根冠比,这是降香黄檀对于逆环境的一种适应性表现。
光照度的空间变化规律随方向和坡度的变化而变化,而中国处于北半球的温带地区,无论在什么纬度,南面所受的光照要比北面多,且坡度越大差异越显著。本研究可以忽略坡度对光照度造成的影响。在不同方向上,两种不同土壤基质下栽培的降香黄檀根系的生物量分布具有完全不同的分布特征,尤其在南面差异最为显著。以往对根系的研究[14]表明,温度对根系生理活性的影响程度较大,适宜的温度能够提高根系的生理活性,过高的温度容易降低根系的生理活性。与壤土相比,石灰岩颗粒在相同光照下,温度升高要比壤土快,且夜间温度下降也要快于壤土。这也是影响降香黄檀在石漠化土壤条件下根系空间分布一个重要原因。
在对不同土层深度降香黄檀根系生物量的调查中,随土层深度的不断增加,两种不同基质中降香黄檀根系生物量均表现为先增大后减小的趋势,生物量的最大值都出现在10~15 cm的土层中。壤土中降香黄檀根系生物量在不同土层深度中无明显差异,石砾中降香黄檀根系在不同土层深度中存在显著差异,这与石砾中水土条件异质性有关,石砾表层更易受环境因素影响造成水分、养分缺失。
营养物质的丰缺会影响植物侧根的数量[15]。通过对不同径级根系生物量的分析结果表明,直径d<0.5 mm根系是决定降香黄檀根系生物量的主要因素,直径d<3 mm的根系存在显著差异,从吸收功能的角度来看,这样的差异反映了模拟极端石漠化土壤条件对于树木生长的影响。但在模拟极端石漠化土壤条件下生长的降香黄檀,直径d<0.5 mm的根系生物量所占百分比要高于壤土中生长的降香黄檀。根系直径越小,活跃度也高,对环境的变化越敏感[16,17],石砾在一定程度上促进了小径级根系的生长,提高根系的活跃度,使植株可以更好地在贫瘠的石漠化土壤环境中吸收营养物质。这与“植物生长/分化平衡假说:即在有限的营养条件下,植物会增加投资防御[18,19]”相一致。
总之,在模拟极端石漠化土壤环境下,根系的生长很大程度上会受外界环境的制约,但在逆环境中,降香黄檀根系还是会通过增加根系生物量配比以及小径级(d<0.05 mm)的根系生物量这种表型进化的方式来主动适应逆环境。
参考文献:
[1]王世杰.喀斯特石漠化——中国西南最严重的生态地质环境问题[J].矿物岩石地球化学通报,2003,22(2):120-126.
[2]袁道先.我国西南岩溶石山的环境地质问题[J].世界科技研究与发展,1997(5):93-97.
[3]曾 杰,陈青度,李小梅.世界紫檀属树种及其在我国的引种前景[J].广东林业科技,2000,16(4):38-44.
[4]农胜奇.广西岩溶地区石漠化危害及治理对策[J].广西林业科学,2007,36(6):170-172.
[5]曹建华,袁道先,童立强.中国西南岩溶生态系统特征与石漠化综合治理对策[J].草业科学,2008,25(9):40-50.
[6]曹建华,袁道先,章 程,等.受地质条件制约的中国西南岩溶生态系统[J].地球与环境,2004,32(1):1-8.
[7]周铁烽.中国热带主要经济树木栽培技术[M].北京:中国林业出版社,2001.
[8]杨振德,赵岩岩,玉舒中,等.干旱胁迫对降香黄檀幼苗生理特性及根系形态特征的影响[J].林业科技开发,2014,28(3):63-66.
[9]刘 震,玉舒中,陈 欢,等.不同石砾含量栽培基质对降香黄檀幼苗生长及生理指标的影响[J].湖北农业科学,2016,55(19):4941-4944,5038.
[10]杨昌儒,刘 震,倪周游,等.不同石砾含量栽培基质对降香黄檀生理指标的影响[J].湖北农业科学,2017,56(17):3287-3291.
[11]宋维峰,王希群.林木根系研究综述[J].西南林学院学报,2007,27(5):9-12.
[12] BROWN S.Measuring carbon in forests:C urrent status and future challenges[J].Environ Pollut,2002,116(3):363-372.
[13] MOKANY K,RAISON R,PROKUSHKIN A S.Criticalanalysis of root:S hoot ratios in terrestrial biomes[J].Global Change Biol,2006,12(1):84-96.
[14]李 鹏,赵 忠,李占斌,等.渭北黄土区刺槐根系空间分布特征研究[J].生态环境学报,2005,14(3):405-409.
[15] POSTMA J A,DATHE A,LYNCH J P.The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability[J].Plant Physiology,2014,166(2):590-602.
[16] NORBY R J,JACKSON R B.Root dynamics and global change:Seeking an ecosystem perspective[J].New Phytologist,2000,147(1):3-12.
[17] YUAN Z Y,CHEN H Y H.Fine root biomass,production,turnover rates,and nutrient contents in boreal forest ecosystems in relation to species,climate,fertility,and stand age:Literature review and meta-analyses[J].Critical Reviews in Plant Sciences,2010,29(4):204-221.
[18] WAREING P F.PHILLIPS I D J.Growth and differentiation in Plants[M].The Third Edition.Pergamon Press,Oxford:1981.481-500.
[19] MOREIRA X,ZAS R,SOLLA A,et al.Differentiation of persistent anatomical defensive structuresis costly and determined by nutrient availability and genetic growth-defence constraints[J].Tree Physiology,2015,35(2):112-123.
