基于主成分分析的杏李砧木耐盐性综合评价
2018-04-09卢明艳张东亚张富玮
卢明艳, 潘 越, 安 鹭, 张东亚, 张富玮, 王 涛, 张 浩
(新疆林业科学院园林绿化研究所,新疆乌鲁木齐 830000)
土壤盐碱化日趋成为一个全球化问题,目前我国盐碱土地面积达0.27亿hm2,约占全国耕地面积的10%[1-2]。新疆维吾尔自治区为封闭内陆地区,受气候条件影响,夏季高温少雨、昼夜温差较大,是我国盐渍化较为集中的省份之一[3]。杏李(Prunusdomestica×armeniaca)是杏和李通过多代杂交培育而成的新型优质水果,具有果个大、果实口味独特、富含多种营养、耐贮藏强等优点,逐渐成为市场不可或缺的水果[4]。新疆林业科学院自2004年陆续引种杏李试栽,并经阿克苏地区佳木、昌吉州吉木萨尔多地进行试验种植,各项技术指标均达到或接近美国原产地水平。
嫁接是常见的无性繁殖方法,而筛选抗逆性强的砧木能够提高杏李对极端盐碱环境的耐受力,进而为杏李早产、丰产提供保障[5]。目前,有学者在苹果、海棠优良砧木筛选上获得成功[6-7],国内未见杏李耐盐砧木的相关报道,筛选出适合杏李的耐盐砧木,对推进其产业发展具有重要的意义。本试验以2年生山杏、山桃、毛桃实生苗为试材,测定不同盐碱浓度胁迫下3种砧木的相关生理指标,并辅以生长量、生物量动态观测,基于主成分分析进行综合评价,以期为杏李砧木的选择及杏李砧木耐盐性评价提供科学依据和理论参考。
1 材料与方法
1.1 试验材料
试验在新疆林业科学院院内进行,选择2年生山杏、山桃、毛桃实生苗为试材,采用规格为28 cm×30 cm的盆进行栽种,4月上盆,每盆栽1株苗,基质为田园土 ∶草炭土=1 ∶3,质量 6 kg;缓苗3个月,选取长势基本一致的苗木进行盐胁迫处理。
1.2 试验设计
采用完全随机区组设计,对每个树种分别采取0%(对照)、0.1%、0.2%、0.4%、0.6%共5个NaCl盐浓度进行处理,每处理9株,重复3次。为避免盐冲击效应,采取多次施盐方法,即先采用0.1% NaCl进行低浓度处理,后每天递增0.1%进行较高浓度处理,连续处理5 d,7月15日5个处理同时达到预定NaCl浓度。此后进行常规管理,每个盆定量浇水1 000 mL,确保没有液体渗出花盆。胁迫处理35 d,观测记录各树种的盐害症状。
1.3 测定指标及方法
1.3.1株高生长量和相对生物量的测定每个树种随机选取3株,测定盐胁迫前后的株高,分别为H1、H2,则株高生长量=H2-H1。盐胁迫结束,每个树种随机选取3株,105 ℃杀青1 h,80 ℃烘至恒质量,称干质量,精确到0.01 g。
1.3.2生理指标的测定采集相同节位的叶片,液氮中速冻带回实验室,超低温冰箱中保存,备用。分别采用DDS-11型电导仪、丙酮法、考马斯亮蓝G-250法测定相对电导率、叶绿素含量、可溶性蛋白含量。
1.3.3保护酶及渗透调节物质的测定丙二醛、脯氨酸、可溶性糖含量及超氧化物歧化酶(SOD)、过氧化物酶(POD)活性的测定参照南京建成生化公司的试剂盒说明方法进行。采用9株幼苗混合取样,取0.1 g鲜叶,置于0.9 mL生理盐水冰浴中充分研磨,制备成10%的组织匀浆;4 ℃ 3 500 r/min 离心10 min;取上清液,备用。分别采用酶标仪、酸性茚三酮法、蒽酮比色法法测定丙二醛、脯氨酸、可溶性糖含量[8]。SOD活性测定以1 mg组织蛋白在1 mL反应液中SOD抑制率达50%时所对应的SOD量为1个活力单位(U),POD活性测定以1 mg组织蛋白1 min催化0.01 μmol H2O2的量为1个活力单位(U),均用U/mg表示酶活性。重复3次。
1.4 数据统计分析
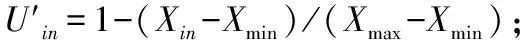
2 结果与分析
2.1 NaCl胁迫对山杏、毛桃、山桃株高生长量、生物量的影响
由图1、图2可见,盐胁迫下,山杏、毛桃、山桃3种砧木的生长量和生物量均不同程度地受到抑制;0~0.2%盐胁迫时,毛桃株高生长量先升后降,山桃株高生长量先降后升,变化幅度均不大;0.2%~0.6%盐胁迫时,山杏株高生长量大幅降低,树体生长受盐胁迫作用明显;含盐量为0.6%时,毛桃、山桃株高生长量与清水处理(对照)相比差异不明显,但均明显高于山杏。毛桃的生物量相对最高,山杏最低,二者平均相差达33.73 g。


2.2 NaCl胁迫对山杏、毛桃、山桃叶绿素含量的影响
盐胁迫会导致植物光同化作用减弱,叶绿素合成受阻[10]。由图3可见,0~0.2%盐胁迫时,毛桃、山杏的叶绿素含量均有所下降,山桃叶绿素含量不减反增,说明山桃受盐胁迫危害程度相对较小;0.2%~0.6%盐胁迫时,3个树种的叶绿素含量均有不同程度下降;含盐量为0.6%时,树种间叶绿素含量有明显差异,山桃叶绿素含量相对最高,毛桃次之,山杏相对最低。
2.3 NaCl胁迫对山杏、毛桃、山桃叶片渗透调节物质的影响
2.3.1可溶性糖含量由图4可见,随盐含量的增加,3个树种的可溶性糖含量变化趋势不一,毛桃、山杏的可溶性糖含量大致呈“先升后降”趋势,山桃的可溶性糖含量呈“升—降—升”趋势;盐含量为0.2%时,毛桃的可溶性糖含量相对最高,明显高于山桃、山杏,后出现大幅下降;0~0.2%盐胁迫时,山桃的可溶性糖含量变化不大,盐含量为0.4%时,山桃的可溶性糖含量相对最高,比CK高1.45 nmol/g,说明其在较高浓度盐环境中能通过储存可溶性糖来维持细胞的渗透调节功能。

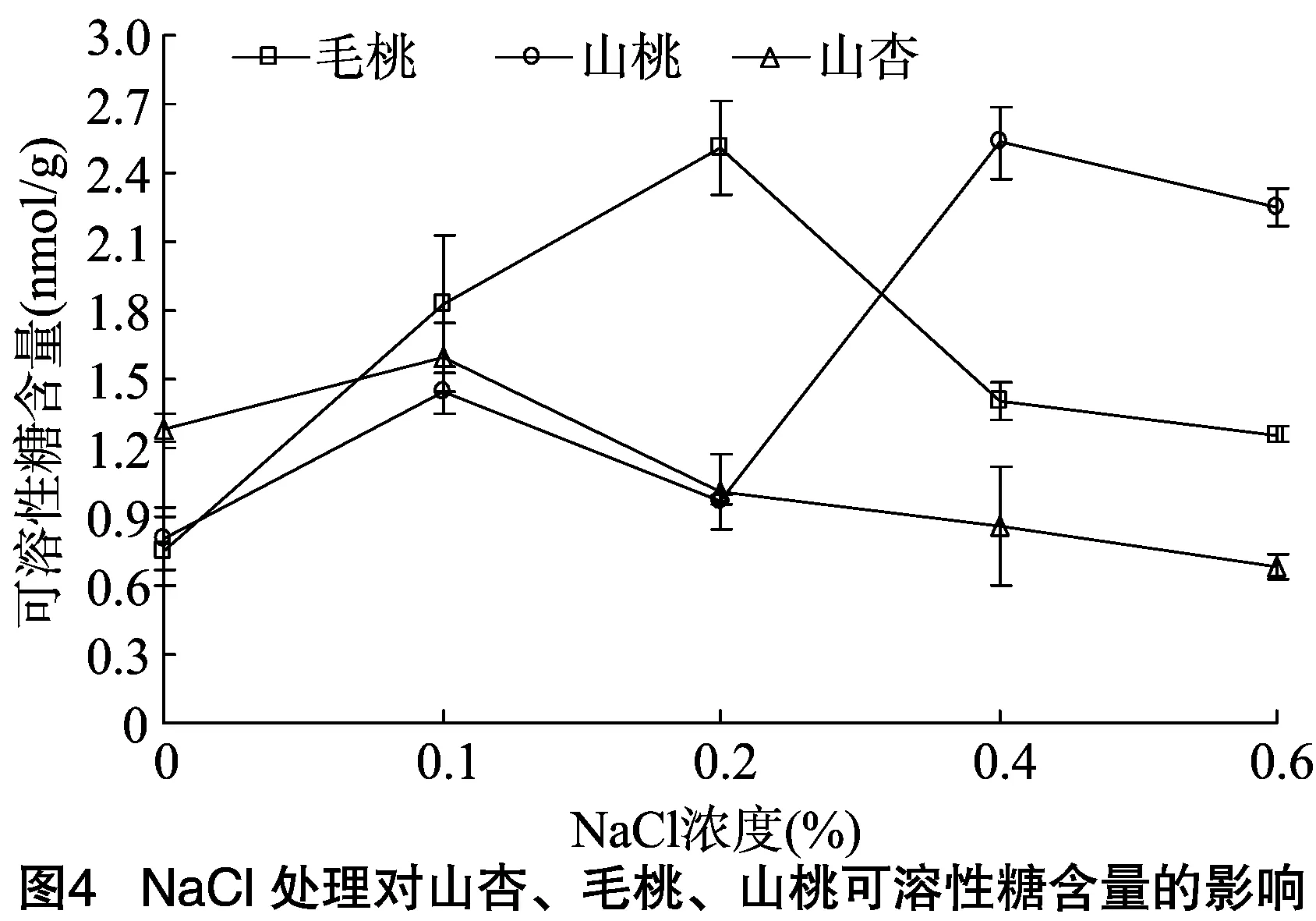
2.3.2可溶性蛋白含量可溶性蛋白质能通过降低细胞水势、调节叶片渗透势来保护叶片细胞膜的渗透调节功能。由图5可见,随盐胁迫程度的加剧,毛桃的可溶性蛋白含量呈逐渐降低趋势,含盐量为0.6%时可溶性蛋白含量较CK减少3.39 mg/g;0.2%盐胁迫时,山桃的可溶性蛋白含量较对照下降16.29%,0.2%~0.6%盐胁迫时,山桃的可溶性蛋白含量持续上升,并在盐含量为0.6%时相对最高,明显高于毛桃、山杏。
2.3.3脯氨酸含量由图6可见,随盐胁迫浓度的增加,山桃、山杏的脯氨酸含量总体呈“先升后降”变化趋势,毛桃呈递增趋势;0~0.2%盐胁迫时,山桃对脯氨酸积累相对最快,较CK增加57.54 μg/g,毛桃较CK增加43.72 μg/g,山杏脯氨酸含量虽有所增加,但与CK相比差异不明显;盐胁迫浓度为0.6%时,毛桃的脯氨酸含量分别是山桃、山杏的1.94、3.22 倍,说明毛桃对盐胁迫的应激保护生理能力相对较强。

2.4 NaCl胁迫对山杏、毛桃、山桃叶片保护酶活性的影响
由图7、图8可见,0~0.2%盐胁迫时,山桃、毛桃的POD活性逐渐增加,而山杏POD活性则较CK下降8.64 U/mg;0.2%~0.6%盐胁迫时,山桃叶片的POD活性呈“先升后降”趋势,且含量始终高于毛桃和山杏,毛桃的POD活性虽有所降低,但下降幅度不大;山杏的POD活性在盐含量为0.2%时达到最小值,后随盐浓度的增加逐渐上升。山桃的SOD活性变化呈“先降后升”趋势,盐含量为0.1%时达到最小值;随盐胁迫浓度的增加,毛桃的SOD活性变化不大。


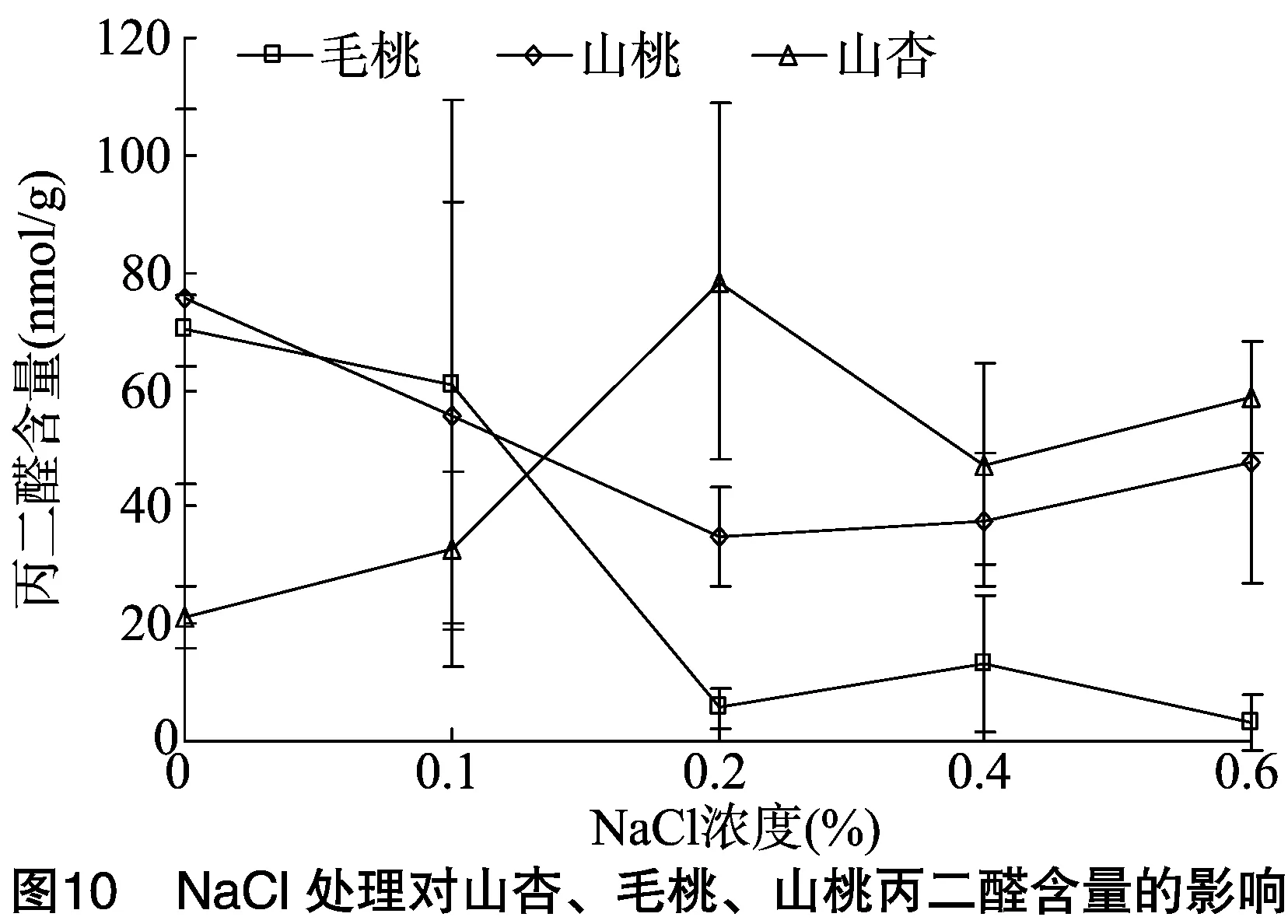
2.5 NaCl胁迫对山杏、毛桃、山桃叶片相对电导率和丙二醛含量的影响
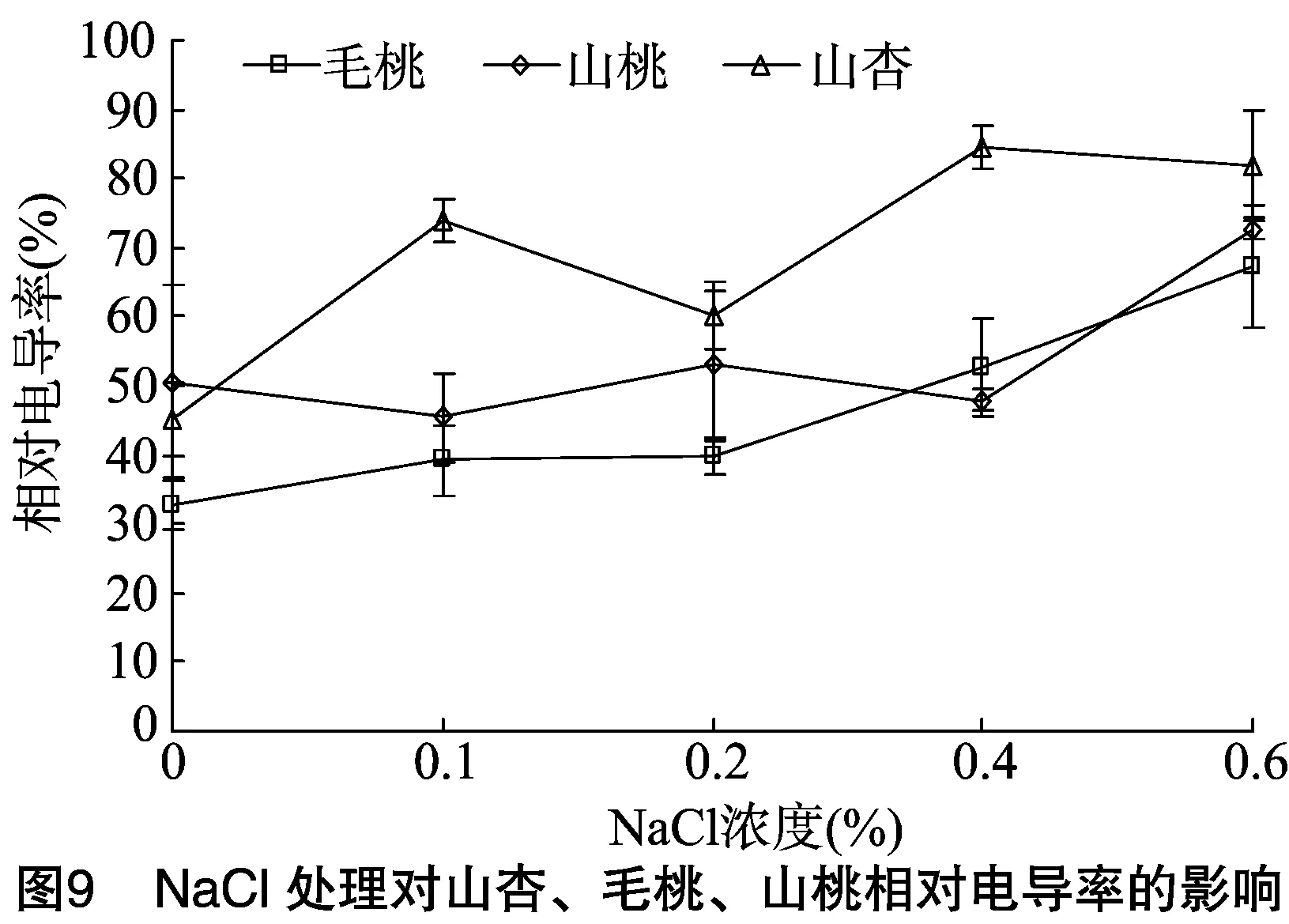
2.5.1相对电导率由图9可见,3种砧木叶片的相对电导率呈不同程度上升趋势,但不同砧木的相对电导率上升幅度差异相对较大;0~0.2%盐胁迫时,山杏变化幅度相对最大,对盐环境的应激反应相对较强;0.2%~0.6%盐胁迫时,毛桃相对电导率上升幅度相对最大,0.6%盐胁迫时的相对电导率较0.2%盐胁迫时的提高27.28%, 山桃上升幅度相对较小,较0.2%盐胁迫时的提高19.68%,对盐胁迫不敏感。
2.5.2丙二醛含量丙二醛是评价细胞受盐碱破坏程度高低的重要指标之一[11]。由图10可见,3种砧木受盐胁迫时丙二醛含量的变化趋势不一,随盐胁迫浓度增加,山杏丙二醛含量先升后降,在盐碱浓度为0.2%时丙二醛含量达到最高,明显高于毛桃、山桃;0.2%盐胁迫时,毛桃的丙二醛含量较CK下降92.04%,高于山桃的53.98%。说明毛桃具有一定的耐盐能力,在轻度和重度盐环境下其细胞膜并未受到明显的损坏。
2.6 3种杏李砧木的综合评价
2.6.1基于隶属函数法的综合评价由于试验调查的耐盐性指标数量纲不一致,故先采用隶属函数法对试验数据进行标准化,将各指标隶属函数值求和即可得出不同砧木的耐盐综合得分。由表1可见,3种砧木耐盐能力由高到低排序为毛桃>山桃>山杏。

表1 不同砧木耐盐性指标标准化结果
2.6.2基于主成分分析法的综合评价对标准化后的数据进行基于主成分分析的综合评价。由表2可见,10项耐盐指标提取出2个主成分,旋转后因子特征与方差贡献结果可简化变量的因子数,其中第1主成分特征根为5.790,代表10项耐盐指标信息总量的57.90%,第2主成成分特征根为 4.210,代表10项耐盐指标信息总量的42.10%,累计贡献率达100%,通过2个主成分能够反映原始变量的所有信息。因此,采用2个公因子代替原有的10项指标对杏李砧木耐盐性进行综合评价,因子经旋转后与变量间的对应关系更加清晰[12]。由表3可见,第1主成分系数为正且载荷值相对较高的指标有叶绿素、可溶性糖、可溶性蛋白含量和POD活性,载荷值分别为0.768、0.890、0.993、0.991,载荷值较高且系数为负的指标有生长量,载荷值为-0.989,说明第1主成分主要反映叶绿素含量、可溶性糖含量、可溶性蛋白含量、植株生长量、POD活性这5项耐盐指标对砧木的影响;第2主成分载荷值相对较高且系数为正的指标有生物量、相对电导率、丙二醛含量,载荷值分别是0.951、0.780、0.987,载荷值较高且系数为负的指标有SOD活性,载荷值为-0.743,说明第2主成分主要反映生物量、相对电导率、丙二醛含量、SOD活性这4项耐盐指标对砧木的影响程度。用表3各项调查指标的载荷值除以相应主成分对应的特征根,然后开平方根,即可得到各个指标的系数即为特征向量,将特征向量与标准化后的数据相乘,可以得到2个主成分的函数表达式分别为
f1=-0.413ZX1+0.232ZX2+0.364ZX3+0.392ZX4+0.414ZX5+0.349ZX6+0.414ZX7-0.340ZX8+0.329ZX9-0.166ZX10;
f2=-0.186ZX1+0.475ZX2+0.390ZX3+0.329ZX4+0.168ZX5+0.410ZX6+0.178ZX7-0.420ZX8+0.430ZX9+0.484ZX10。
由表4可见,以公因子f1排序时,山桃排名第1,其作为砧木主要优势体现在受盐胁迫时枝条营养储存充足,对叶片光合作用、枝条生长影响相对较低,山杏排名相对最低,说明其受盐胁迫危害程度相对最大;以公因子f2排序时,毛桃排名第1,作为砧木其优势在于细胞膜脂氧化受盐胁迫影响相对较低,同时质膜透性受盐离子破坏程度较低,干物质积累量相对较大,山杏细胞受盐胁迫影响程度相对最大,细胞平衡破坏严重,得分相对最低;综合得分值由低到高依次为毛桃>山桃>山杏,这与隶属函数法分析结果一致。

表2 主成分的特征根、方差贡献率和累计贡献率

表3 旋转后的主成分载荷矩阵

表4 不同砧木耐盐碱指标得分及综合得分
3 结论与讨论
采用隶属函数法对生化指标单位进行统一,能够避免因数量纲不一致导致的试验误差,增加数据结果的精确性和可靠性。综合来看,3种砧木耐盐性由高到低为毛桃>山桃>山杏,其中山桃长势强,可溶性糖、可溶性蛋白含量高,叶绿素含量也较高,毛桃干物质含量积累多,细胞膜脂保护作用及抗氧化能力相对较强。当细胞组织受到伤害时,叶片MDA含量增加,膜脂过氧化加剧[13-14],引起叶片膜透性的增加[15]。夏阳等研究NaCl胁迫下苹果幼树叶片膜透性的变化发现,0.4% 盐胁迫处理的相对电导率在中前期高于对照,植物细胞膜透性受到伤害[16]。杜中军等报道,小金海棠和平邑甜茶叶片细胞膜透性随盐胁迫加重而增大,耐盐能力强的小金海棠膜透性在不同盐胁迫时间和胁迫强度下都低于平邑甜茶[17]。本试验研究发现,随NaCl浓度的增加,山桃、毛桃、山杏的相对电导率呈增加趋势,0.6% NaCl处理的山桃、毛桃、山杏叶片相对电导率分别为对照的2.26、1.79、1.58倍,说明山桃、毛桃、山杏对0.6% NaCl处理都有一定程度的耐受性。
活性氧代谢失调引起自由基积累及自由基对大分子的破坏作用是需氧生物遭受逆境伤害的重要特征。活性氧在植物体内的清除则需SOD、POD等膜保护酶系统[18-19]。克热木·伊力等发现,NaCl胁迫下长果阿月浑子叶片中的SOD、POD活性随盐浓度的增大而增强[20]。李会云等研究葡萄砧木品种(系)的耐盐性结果表明,随土壤含盐量增加,SOD、POD活性呈先升后降趋势[21]。本试验结果表明,在盐胁迫初期(0~0.2%),山桃和毛桃POD活性均逐渐增加,而山杏POD活性呈下降趋势。而盐胁迫后期(0.2%~0.6%),山桃叶片POD活性呈先升后降趋势,但含量始终高于毛桃和山杏;毛桃次之,POD活性虽降低,但幅度不大;山杏较低,在盐含量0.2%达最低后POD活性开始上升。山桃SOD活性变化均呈"先降后升"趋势,盐含量0.1%时达最低值,随着对盐碱土壤的适应,SOD活性明显上升,毛桃随盐胁迫SOD变化不大。由此可见,SOD、POD活性在不同果树盐胁迫反应中的变化规律不尽相同,可能是不同果树应答盐胁迫的机制不同,也可能是不同酶在不同树种中的作用途径不完全一样所致。
参考文献:
[1]李淑娟,詹亚光,杨传平,等. 混合盐胁迫对引种绒毛白蜡生长及相关生理指标的影响[J]. 东北林业大学学报,2010,38(1):15-17.
[2]吴运荣,林宏伟,莫肖蓉. 植物抗盐分子机制及作物遗传改良耐盐性的研究进展[J]. 植物生理学报,2014,50(11):1621-1629.
其中,L为双基基线长度,RS为双基距离和,RT,RR分别为发射机、接收机与目标之间的距离,HT,HR分别为发射机、接收机的飞行高度,VT,VR分别为发射机、接收机的平台运动速度,φT,φR分别为地面目标散射点相对于发射、接收平台天线轴向的俯仰角,ψT,ψR分别为发射、接收平台波束指向的空间角,δT,δR分别为发射、接收平台载机飞行方向相对于基线方向的方位角,θFT,θFR分别为发射、接收雷达天线波束指向相对于基线方向的方位角,θAT,θAR分别为发射、接收雷达天线相对于载机飞行方向的夹角。
[3]寇云玲. 杂交榛耐盐碱性品种(系)筛选研究[D]. 乌鲁木齐:新疆农业大学,2013:1-56.
[4]牛俊莉,刘茂秀,彭秋梅. 不同采摘期杏李味厚品种理化指标主成分及聚类分析[J]. 新疆农业科学,2015,52(1):33-36.
[5]桂毓,杨静慧,刘艳军,等. 盐碱土对不同樱桃品种生长的影响[J]. 北方园艺,2016(13):33-35.
[6]曹敏格,杨海玲,张文,等. 苹果砧木矮化性评价指标的研究[J]. 中国农业大学学报,2008,13(5):11-18.
[7]成明昊,杨晓红,曾维光. 苹果砧木资源——小金海棠的调查研究初报[J]. 西南农学院学报,1984(3):38-43.
[8]韩俊威,董玉芝,宋锋惠,等. 平欧杂交榛抗寒生理特性研究[J]. 西北农林科技大学学报(自然科学版),2014,42(6):138-144.
[9]潘越,史彦江,陈淑英,等. 喷施叶面肥对平欧杂种榛‘新榛1号’嫩枝扦插的影响[J]. 江西农业大学学报,2016,38(5):920-926.
[11]李学强,李秀珍. 盐碱胁迫对欧李叶片部分生理生化指标的影响[J]. 西北植物学报,2009,29(11):2288-2293.
[12]公丽艳,孟宪军,刘乃侨,等. 基于主成分与聚类分析的苹果加工品质评价[J]. 农业工程学报,2014,30(13):276-285.
[13]Ferrari I R,Phamt A,Zuily F Y,et al. Effect of drought on photodynamic peroxidation of leaf total lipophilic extracts[J]. Phytochemistry,1994,37(5):1237-1243.
[14]Pallak G,Waise L Y. Exophysiolgy of salt excretion inAeltropslitrolis(Gramineae)[J]. Physiologia Plantarum,1979,47(3):177-184.
[15]孙黎,刘士辉,师向东,等. 10种藜科盐生植物的抗盐生理生化特征[J]. 干旱区研究,2006,23(2):309-818.
[16]夏阳,梁慧敏,束怀瑞,等. NaCl胁迫下苹果幼树叶片膜透性、脯氨酸及矿质营养水平的变化[J]. 果树学报,2005,22(1):1-5.
[17]杜中军,翟衡,李健,等. 盐胁迫对苹果砧木的膜伤害[J]. 山东农业大学学报(自然科学版),2001,32(4):532-534.
[18]杜中军,翟衡,潘志勇,等. 盐胁迫下苹果砧木光合能力及光合色素的变化[J]. 果树学报,2001,18(4):200-203.
[19]黄雪妮,屈凡,马名立,等. 镉胁迫对2个宁夏主栽水稻品种幼苗期抗氧化同工酶亚基及其活性的影响[J]. 江苏农业科学,2016,44(7):107-112.
[20]克热木·伊力,袁琳,齐曼·尤努斯,等. 盐胁迫对阿月浑子SOD、CAT、POD活性的影响[J]. 新疆农业科学,2004,41(3):129-134.
[21]李会云,郭修武. 盐胁迫对葡萄砧木叶片保护酶活性和丙二醛含量的影响[J]. 果树学报,2008,25(2):240-243.
