海岛棉和陆地棉花铃期光合特性及氮素累积特性的差异
2018-04-09李春艳石洪亮文如意严青青张巨松
李春艳,石洪亮,文如意,严青青,张巨松
(新疆农业大学/教育部棉花工程研究中心,乌鲁木齐830052)
棉纤维是用途广泛的天然纤维[1]。棉花是新疆的支柱产业,同时新疆也是我国最大的产棉区[2]。海岛棉纤维具有长、细、强等突出特点,在棉花栽培种中品质最优,是纺高支精梳纱和特种织物的重要原料[3-4]。前人在陆地棉与海岛棉冠层结构以及叶片内在组织结构等方面做了许多研究,结果表明冠层结构差异是导致海岛棉与陆地棉干物质质量不同的原因之一[5-7],同时栅栏组织较薄限制了海岛棉光合潜力进一步发挥[8]。对海岛棉产量形成的相关研究表明,由于铃重、衣分均较小,丰产和稳产性能差,海岛棉产量水平总体不高。
虽然大量研究已表明海岛棉在各方面均与陆地棉存在差异,但在实际生产中除打顶时间与陆地棉不同,水肥的管理均相同,这对提高海岛棉的产量极为不利[9]。目前针对陆地棉与海岛棉在相同管理措施下,花铃期净光合速率及叶片温度,全生育期干物质、氮素累积分配的差异这方面的研究还较少。本研究选用具有相同生育期的陆地棉与海岛棉品种,分析花铃期光合速率、叶片温度、光照度以及干物质、氮素积累与分配的规律,探索海岛棉与陆地棉干物质、氮素的差异,通过大田栽培措施来调节干物质、氮素在生殖器官与营养器官中的分配,从而提高产量。
1 材料与方法
1.1 试验区概况
试验于2015年4―10月在新疆阿瓦提县新疆农业科学院经济作物研究所试验基地进行,地处北纬 39°31′~40°50′、东经 79°45′~81°5′,无霜期 183~227 d,年均日照时间 2 750~3 029 h,全年≥10℃积温3 802.9℃。该地区属暖温带大陆性干旱气候,年平均降水量46.7 mm,年平均蒸发量1 890.7 mm。试验地土壤为砂壤土,前茬作物为棉花,播前测定小区肥力,结果见表1。
1.2 试验设计
以生育期作为选择品种的主要依据,选择具有相近生育期的4个品种,分别为新陆中75号、新陆中54号、新海35号和新海48号,其中新陆中54号、新海35号生育期为140 d,新海48号的生育期为136~140 d,新陆中75号为143 d左右。试验采用单因素随机区组设计。采用机采棉种植模式,行距、株距配置为[(66+10)×11]cm,一膜4行,重复4次,小区长6.5 m,宽4.56 m,小区面积为29.64 m2,试验区总面积约为474.24 m2,理论密度为24万株·hm-2,总灌水量为4 500 m3·hm-2,总施肥量为 300 kg·hm-2。 4 月 14 日人工点播,7月10日统一打顶,其他管理措施同大田。

表1 试验地土壤基础肥力Table 1 Foundation fertility of the trial soil
1.2.1测定项目及方法。干物质及全氮含量:于棉花现蕾期、盛蕾期、初花期、盛花期、盛铃期和吐絮期取样,选取具有代表性的6株棉花,按根、叶、茎、蕾、铃壳和棉纤维等不同器官分开,放入电热恒温鼓风干燥箱105℃杀青30 min,然后80℃恒温烘干至质量恒定,测定其干物质质量。烘干的棉株样品经粉碎,过孔径0.5 mm的筛,备用。以H2SO4-H2O2进行消煮定容后,采用奈氏比色法测定棉株不同部位的全氮含量。
净光合速率(Pn)、 气孔导度(Gs): 利用Hansatech公司(英国)生产的TPS-2光合仪,于棉花盛蕾期、盛花期、盛铃期,选择晴朗天的11:00-13:00,测定叶片净光合速率及气孔导度,打顶前测定倒四叶,打顶后测定倒三叶,每个处理测定5张叶片,每张叶片重复测3次。在盛花期,选择晴朗天,08:00-20:00 每隔 2 h 测定 1次净光合速率和气孔导度,测定倒三叶。
叶片温度及光照度:从现蕾期开始至盛铃期,应用防水型UA-002-08数字温度光照记录仪,于棉株宽行测定棉株冠层中部的叶片温度及光照度(参数设置为每隔1 h测1次)。
产量:在吐絮期取上、中、下各30个棉铃测定铃重和衣分,取平均值,以各小区实际收获籽棉产量计产。
1.2.2数据处理。所获得数据用MS Excel 2010数据处理软件进行初步分析和表格制作,用DPS 7.05统计软件进行方差分析,采用最小显著差数法(Least significant difference,LSD)进行差异显著性检验(α=0.05)。本文对大面积推广的具有相近生育期的新陆中54号及新海48号相关数据进行详细分析。
2 结果与分析
2.1 花铃期陆地棉与海岛棉净光合速率、叶片温度的差异

图1 陆地棉与海岛棉花铃期净光合速率的差异Fig.1 Differences of net photosynthetic rate of upland cotton and island cotton in boll stage
海岛棉2个品种净光合速率在盛蕾期、盛花期、盛铃期均低于陆地棉2个品种,4个品种Pn最大值均出现在盛花期,陆地棉与海岛棉品种之间Pn差异不显著(图1)。新陆中54号与新海48号比较,在盛蕾期、盛花期2个品种之间存在显著性差异,新陆中54号较新海48号高20.62%、27.46%,在盛铃期新陆中54号较新海48号高11.72%,且差异也显著。在花铃期测定4个品种冠层中部温度和光照度,结果表明海岛棉2个品种的叶片温度、光照度较陆地棉2个品种高(表2),其中新陆中54号的叶片温度较新海48号在盛蕾期、盛花期、盛铃期分别低0.824、2.421、2.993℃。如图2所示,在盛花期定点测定棉株倒三叶光合速率日变化,从而建立Gs和Pn的相关性,结果显示,2条直线重合度较高,说明海岛棉和陆地棉之间Gs对Pn的影响差异极小。
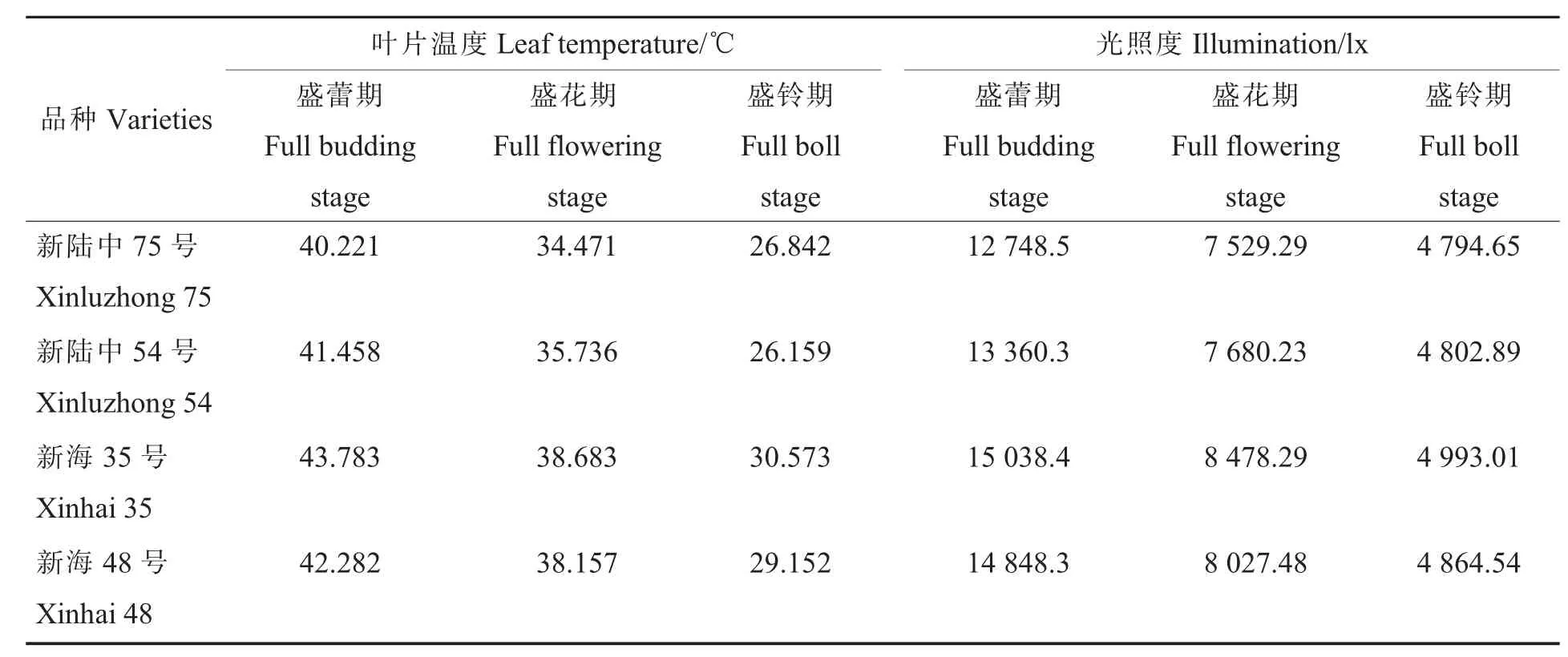
表2 陆地棉与海岛棉叶片温度及光照度的差异Table 2 Differences between island cotton and upland cotton leaf temperature and intensity
2.2 陆地棉与海岛棉干物质积累与分配
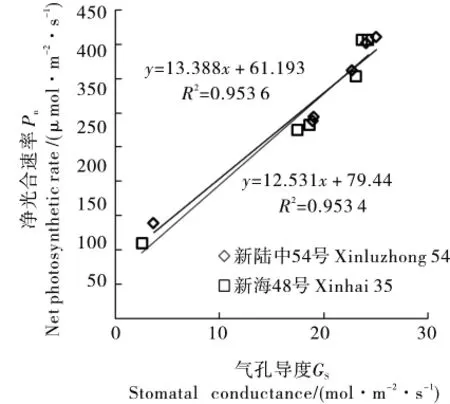
图2 盛花期海岛棉、陆地棉气孔导度(Gs)和光合速率(Pn)的关系Fig.2 The relationship between stomatal conductance(Gs)and photosynthetic rate(Pn)of island cotton and upland cotton at the full flowering stage

表3 陆地棉和海岛棉干物质积累与分配Table 3 Dry matter accumulation and distribution of boll in upland cotton and island cotton
由表3可知,在相同的栽培管理条件下,整个生育期内,陆地棉2个品种的单株干物质总量均高于海岛棉2个品种,除在盛铃期新海48号较新陆中75号多8.36 g。陆地棉2个品种根、叶片的干物质积累量占总量的比例在花铃期内均小于海岛棉2个品种;茎干物质积累量占总量的比例在花铃期内均大于海岛棉两个品种;蕾花铃干物质积累量占总量的比例在盛蕾期、盛花期小于海岛棉2个品种,在盛铃期大于海岛棉2个品种;其中以生育期相同的新陆中54号和新海48号进行干物质积累量及分配比例的比较,均符合以上规律;在整个花铃期内新陆中54号与新海48号的干物质总量、根、茎、叶、蕾花铃都存在显著性差异(P<0.05)。从单株干物质总量的日增量来看,从盛花期至盛铃期新陆中54号为2.47 g·d-1,新海 48号的为1.32 g·d-1;分析比较其他时期单株干物质积累量,发现此阶段是陆地棉干物质积累量较海岛棉多的关键时期。从单株生殖器官日增长量来看,从盛蕾期到盛铃期新陆中54号为0.66 g·d-1,新海48号的为0.19 g·d-1,虽然新陆中54号的生殖器官占干物质总量的比例小,但生殖器官日增长量较新海48号快。以上结果均可说明陆地棉的光合产物较海岛棉优先向生殖器官分配。
2.3 陆地棉与海岛棉氮素积累与分配
2.3.1陆地棉与海岛棉氮素积累总量的Logistic模型及其特征值。陆地棉和海岛棉氮素总量均随着生育进程的推进呈“S”型变化曲线,亦符合Logistic生长函数模型(表4)。陆地棉2个品种氮素快速积累开始期较海岛棉2个品种晚1~2 d,氮素快速积累结束期早3~8 d;氮素快速积累持续时间上,陆地棉2个品种较海岛棉2个品种短4~10 d;从氮素积累速率最大时刻来看,各栽培种品种之间开始时间差异较大。陆地棉2个品种的氮素快速积累持续期短于海岛棉2个品种,但陆地棉积累速率快,因此氮素积累总量多。其中新陆中54号较新海48号氮素快速积累开始期晚1 d,氮素快速积累结束期早8 d,氮素快速积累持续时间少10 d,但氮素积累速率最大时刻的积累速率高;因此,新陆中54号较新海48号氮素积累量多。
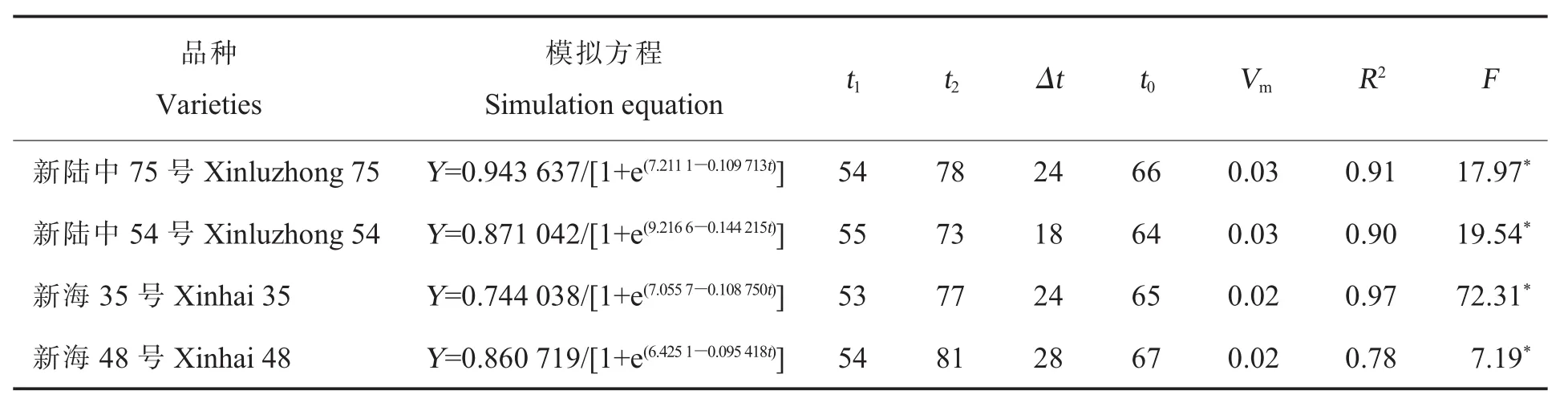
表4 氮素积累总量的Logistic模型及其特征值Table 4 Logistic model of total nitrogen accumulation and its characteristic value
2.3.2陆地棉与海岛棉氮素生殖器官氮素积累的Logistic模型及其特征值。陆地棉和海岛棉生殖器官氮素积累总量均随着生育进程的推进呈“S”型变化曲线,亦符合Logistic生长函数模型(表5)。从生殖器官氮素快速积累开始期来看,陆地棉2个品种较海岛棉2个品种晚0~10 d,生殖器官氮素快速积累结束期晚5~7 d,在氮素快速积累持续时间方面,不同品种之间差异较大,其中陆地棉2个品种较新海48号持续时间长3~7 d,但新海35号较陆地棉2个品种持续时间长1~5 d。生殖器官氮素积累速率最大时刻陆地棉2个品种较海岛棉2个品种早3~8 d,生殖器官氮素积累速率最大时刻的速率陆地棉2个品种与海岛棉2个品种相同。其中新陆中54号较新海48号,生殖器官氮素快速积累开始期相当,快速积累持续期长7 d,生殖器官氮素积累速率最大时刻的积累速率相同,即新陆中54号较新海48号生殖器官氮素快速积累开始期相同,但持续时间长,使得新陆中54号较新海48号生殖器官氮素积累多。这与其干物质、生殖器官氮素积累量的变化规律相一致。

表5 生殖器官氮素积累的Logistic模型及其特征值Table 5 Logistic model of nitrogen accumulation in reproductive organs and its characteristic
2.4 陆地棉与海岛棉产量的差异
由表6可知,陆地棉2个品种较海岛棉2个品种的单株成铃少0.38~1.49个,但不存在显著性差异;2个栽培品种之间铃重存在显著性差异,陆地棉较海岛棉2个品种高1.72~1.87 g;在籽棉产量方面,陆地棉2个品种较海岛棉2个品种高 862.69~1 429.09 kg·hm-2。在本试验条件下,陆地棉2个品种较海岛棉2个品种虽然单株结铃数少,但铃大且衣分高,使得籽棉产量高。以生育期相同的新陆中54号和新海48号进行比较,籽棉产量高 872.94 kg·hm-2,铃重高 1.8 g,衣分高10.82百分点。
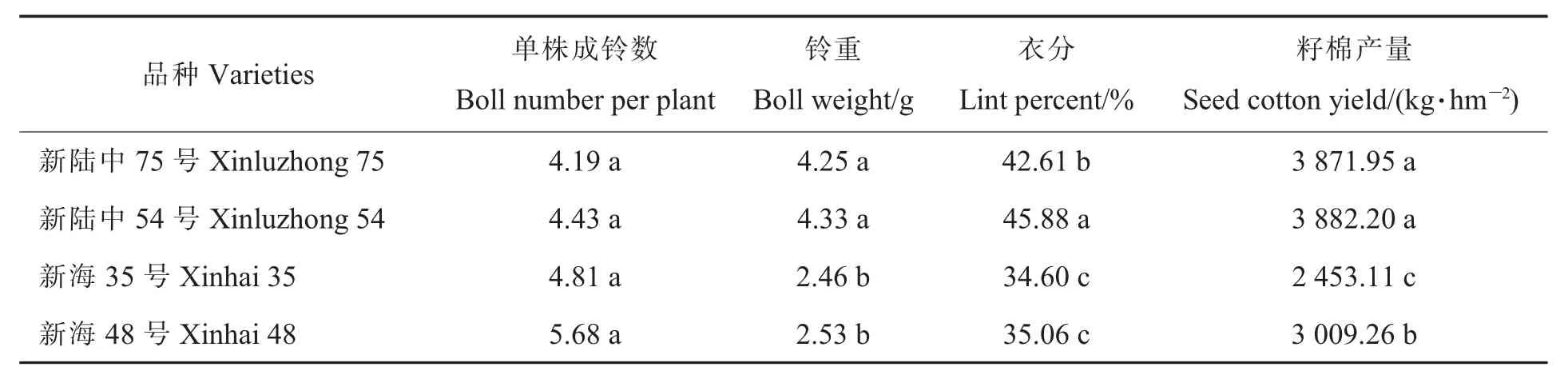
表6 陆地棉和海岛棉产量的差异Table 6 Differences in yield of upland cotton and island cotton
3 结论与讨论
较高的干物质质量是棉花获得高产的基础[10],但棉花的生殖生长中心与顶端优势不重合,光合作用及干物质氮素积累与分配比较复杂[11]。前人研究表明,棉花的净光合速率及干物质、氮素的积累与分配对产量存在一定的影响[12-14]。有研究表明,在1 d中,陆地棉的净光合速率均高于海岛棉[15],这与海岛棉叶片不具有横向日性运动有关[8,16],使其利用外界强光的能力较弱,从而影响其光合物质生产潜力[17-18]。叶片温度、光照度会影响气孔的开闭程度,气孔的开闭会影响气孔导度,从而影响净光合速率[19-20]。本试验结果表明,在花铃期,新陆中54号较新海48号Pn高,叶片温度、光照度低,气孔导度大。
棉株通过调节气孔导度来调节叶片温度[19-20],在盛花期测定光合参数日变化,从而建立新陆中54号与新海48号Pn与Gs相关性曲线,结果显示这2个品种响应曲线无明显差异,表明在相同的胞间CO2浓度下,它们的光合潜力相同[21]。因此,可通过调节种植密度及灌水量、施肥量来调节海岛棉冠层叶片温度及光照度,影响气孔导度和净光合速率,进而使产量有所提升[22]。
在研究陆地棉与海岛棉净光合速率的基础上探索了干物质与氮素的积累与分配规律。在本试验条件下,维持光合产物在生殖器官与营养器官之间的合适分配,是海岛棉提高产量的关键[22]。以生育期相同的新陆中54号与新海48号进行比较可知,在干物质积累与分配方面,新陆中54号较新海48号干物质积累总量多,生殖器官所占干物质总量的比例在盛铃前期小,在盛铃后期大;同时新陆中54号的日积累量及生殖器官日增长量多。有研究表明作物含氮量与干物质积累呈线性正相关性[23]。本试验结果表明,氮素积累量的变化规律与干物质总量的积累与分配相同。
在较高光合物质生产的基础上,进一步提高生物产量的转化率,是提高棉花产量的途径之一[24-25]。本试验条件下,新陆中54号的单株结铃数与新海48号无显著差异,但铃重、衣分显著高于新海48号。可见,相较于新陆中54号,新海48号棉铃的容量小,植株光合物质生产转化效率低。
参考文献:
[1]熊宗伟,顾生浩,毛丽丽,等.中国棉花纤维品质和气候因子的空间分布特征[J].应用生态学报,2012,23(12):3385-3392.Xiong Zongwei,Gu Shenghao,Mao Lili,et al.Spatial distribution characteristics of fiber quality and climate factors in Chinese cotton[J].ChineseJournalofAppliedEcology,2012,23(12):3385-3392.
[2]孔庆平,孔杰,徐海江,等.新疆棉花集约高效生产技术研发策略[J].新疆农业科学,2015,52(7):1352-1358.Kong Qingping,Kong Jie,Xu Haijiang,et al.Strategy of intensive and efficient cotton production technology in Xinjiang[J].Xinjiang Agricultural Sciences,2015,52(7):1352-1358.
[3]孔庆平.我国海岛棉生产概况及比较优势分析[J].中国棉花,2002,29(12):19-23.Kong Qingping.Analysis of production situation and comparative advantage of island cotton in China[J].China Cotton,2002,29(12):19-23.
[4]闫曼曼,郑剑超,张巨松,等.蕾期调亏灌溉对海岛棉光合特性及产量的影响[J].干旱区资源与环境,2016,30(4):115-121.Yan Manman,Zheng Jianchao,Zhang Jusong,et al.Bud period of deficit irrigation on cotton photosynthetic characteristics and yield of island cotton[J].Journal of Arid Land Resources and Environment,2016,30(4):115-121.
[5]姚贺盛,张亚黎,易小平,等.海岛棉和陆地棉叶片光合特性、冠层结构及物质生产的差异[J].中国农业科学,2015,48(2):251-261.Yao Hesheng,Zhang Yali,Yi Xiaoping,et al.The island cotton and upland cotton leaf photosynthesis,canopy structure and material production difference[J].Scientia Agricultura Sinica,2015,48(2):251-261.
[6]胡根海,王志伟,王清连,等.海岛棉与陆地棉叶绿素含量变化的差异研究[J].生物学杂志,2010,27(4):31-34.Hu Genhai,Wang Zhiwei,Wang Qinglian,et al.Study on the variation of chlorophyll content in sea island cotton and upland cotton[J].Journal of Biology,2010,27(4):31-34.
[7]伍维模,董合林,危常洲,等.南疆陆地棉与海岛棉光合-光响应及叶绿素荧光特性分析[J].西北农业学报,2006,15(4):141-146.Wu Weimo,Dong Helin,Wei Changzhou,et al.Photosynthetic light response and chlorophyll fluorescence characteristics of upland cotton and island cotton in South Xinjiang[J].Acta Agriculturae Boreali-Occidentalis Sinica,2006,15(4):141-146.
[8]张亚黎,姚贺盛,罗毅,等.海岛棉和陆地棉叶片光合能力的差异及限制因素[J].生态学报,2011,31(7):1803-1810.Zhang Yali,Yao Hesheng,Luo Yi,et al.The difference and limiting factors of photosynthetic capacity of island cotton and upland cotton[J].Acta Ecologica Sinica,2011,31(7):1803-1810.
[9]菲力申.陆地棉与海岛棉杂种优势利用研究[D].南宁:广西大学,2008.Ndarata F.Study on heterosis utilisation between upland cotton and sea island cotton[D].Nanning:Guangxi University,2008.
[10]刘翠,张巨松,郑慧,等.氮肥基追比对杂交棉光合生理特性及产量的影响[J].干旱地区农业研究,2015,33(2):159-164,169.Liu Cui,Zhang Jusong,Zheng Hui,et al.Effects of topdressing ratio of nitrogen fertilizer on photosynthetic characteristics and yield of hybrid cotton[J].Agricultural Research in the Arid Areas,2015,33(2):159-164,169.
[11]郭仁松,魏红国,富艳荣,等.南疆超高产棉花干物质积累分配与养分吸收运移特征的研究[J].新疆农业科学,2011,48(3):410-418.Guo Rensong,Wei Hongguo,Fu Yanrong,et al.Study in super high yield cotton dry matter accumulation and nutrient absorption and migration characteristics[J].Xinjiang Agricultural Sciences,2011,48(3):410-418.
[12]罗宏海,李俊华,勾玲,等.膜下滴灌对不同土壤水分棉花花铃期光合生产、分配及籽棉产量的调节[J].中国农业科学,2008,41(7):1955-1962.Luo Honghai,Li Junhua,Gou Ling.et al.Regulation of mulched drip irrigation on soil moisture of different cotton boll period of photosynthetic production,distribution and cotton yield[J].Scientia Agricultura Sinica,2008,41(7):1955-1962.
[13]伍维模,郑德明,董合林,等.南疆棉花干物质和氮磷钾养分积累的模拟分析[J].西北农业学报,2002,11(1):92-96.Wu Weimo,Zheng Deming,Dong Helin,et al.Simulation analysis of the accumulation of nitrogen phosphorus and potassium nutrients[J].ActaAgriculturaeBoreali-OccidentalisSinica,2002,11(1):92-96.
[14]Hebbar,K B,Perumal N K,Khadi B M.Photosynthesis and plant growth response of transgenic Bt cotton(Gossypium hirsutumL.)hybrids under field condition[J].Photosynthetica,2007,45(2):254-258.
[15]张亚黎,罗毅,姚贺盛,等.田间条件下海岛棉和陆地棉花铃期叶片光保护的机制[J].植物生态学报,2010,34(10):1204-1212.Zhang Yali,Luo Yi,Yao Hesheng,et al.Mechanism of leaf light protection in island cotton and upland cotton under field conditions[J].Chinese Journal of Plant Ecology,2010,34(10):1204-1212.
[16]Lu Z M,Chen J W,Percy R G.Photosynthetic rate,stomatal conductance and leaf area in two cotton species(Gossypium barbadenseandGossypium hirsutum)and their relation with heat resistance and yield[J].Function Plant Biology,1997,24(5):693-700.
[17]Ehleringer J R,Hammond S D.Solar tracking and photosynthesis in cotton leaves[J].Agricultural and Forest Meteorology,1987,3(1):25-35.
[18]Wise R R,Sassenrath-Cole G F,Percy R G.A comparison of leaf anatomy in field-grownGossypium hirsutumandG.barbadense[J].Annals of Botany,2000,86(4):731-738.
[19]FarquharGD,SharkeyTD.Stomatalconductanceandphotosynthesis[J].AnnualReviewof PlantBiology,1982,33:317-345.
[20]Berry J.Photosynthetic response and adaptation to temperature in higher plants[J].Annual Review of Plant Biology,1980,31:491-543.
[21]Bange M P,Miroy S P.Timing of crop maturity in cotton:Impact of dry matter production and partitioning[J].Field Crops Research,2000,68(2):143-155.
[22]薛晓萍,郭文琦,王以琳,等.不同施氮水平下棉花生物量动态增长特征研究[J].棉花学报,2006,18(6):323-326.Xue Xiaoping,Guo Wenqi,Wang Yilin,et al.Study on the dynamic growth characteristics of cotton biomass under different nitrogen application rates[J].Cotton Science,2006,18(6):323-326.
[23]董合忠,毛树春,张旺锋,等.棉花优化成铃栽培理论及其新发展[J].中国农业科学,2014,47(3):441-451.Dong Hezhong,Mao Shuchun,Zhang Wangfeng,et al.On boll-setting optimization theory for cotton cultivation and its new development[J].Scientia Agricultura Sinica,2014,47(3):441-451.
[24]张亚黎,冯国艺,胡渊渊,等.棉花非叶绿色器官光合能力的差异及与物质生产的关系[J].作物学报,2010,36(4):701-708.Zhang Yali,Feng Guoyi,Hu Yuanyuan,et al.Differences in photosynthetic capacity of cotton non leafy green organs and their relation to matter production[J].Acta Agronomica Sinica,2010,36(4):701-708.
[25]杨英,占东霞,张国娟,等.水分亏缺对棉花叶片与非叶绿色器官光合特性及产量的影响[J].新疆农业科学,2015,52(11):1989-1996.Yang Ying,Zhan Dongxia,Zhang Guojuan,et al.Effects of water deficit on photosynthetic characteristics and yield of cotton leaves and non leaf green organs[J].Xinjiang Agricultural Sciences,2015,52(11):1989-1996.
